
Abstract
Patients with intermittent claudication suffer from both muscle pain and an exacerbated exercise pressor reflex. Excitability of the group III and group IV afferent fibers mediating these functions is controlled in part by voltage-dependent sodium (NaV) channels. We previously found tetrodotoxin-resistant NaV1.8 channels to be the primary type in muscle afferent somata. However, action potentials in group III and IV afferent axons are blocked by TTX, supporting a minimal role of NaV1.8 channels. To address these apparent differences in NaV channel expression between axon and soma, we used immunohistochemistry to identify the NaV channels expressed in group IV axons within the gastrocnemius muscle and the dorsal root ganglia sections. Positive labeling by an antibody against the neurofilament protein peripherin was used to identify group IV neurons and axons. We show that >67% of group IV fibers express NaV1.8, NaV1.6, or NaV1.7. Interestingly, expression of NaV1.8 channels in group IV somata was significantly higher than in the fibers, whereas there were no significant differences for either NaV1.6 or NaV1.7. When combined with previous work, our results suggest that NaV1.8 channels are expressed in most group IV axons, but that, under normal conditions, NaV1.6 and/or NaV1.7 play a more important role in action potential generation to signal muscle pain and the exercise pressor reflex.
Patients with peripheral artery disease often experience intermediate claudication (i.e. muscle pain), which results from atherosclerotic plaques in the major arteries of lower extremities. 1 It has been estimated that 12–18 million adults have peripheral artery disease and one-third of them experience claudication, which negatively impacts their quality of life.2,3 Along with pain, patients suffering from peripheral vascular disease can also experience an exaggerated exercise pressor reflex (EPR) that significantly increases their risk for myocardial infarction. 4 In healthy individuals, the EPR is one of the mechanisms mediating cardiovascular adjustments to exercise.3,5
Muscle nociceptive and EPR signals are transmitted along thinly myelinated group III and unmyelinated group IV axons. 5 In patients with peripheral artery disease, poor muscle perfusion leads to ischemia, which is an activator of group III and IV afferents leading to muscle pain and an enhanced EPR.5,6
The electrogenesis of these signals is mediated via voltage-gated sodium channels (NaV). Dorsal root ganglia (DRG) neurons have been reported to express several NaV channel isoforms, including NaV1.1, NaV1.6–NaV1.9 channels.7,8 One method to distinguish NaV channels is by their sensitivity to tetrodotoxin (TTX), with some channels such as NaV1.6 and NaV1.7 being TTX-sensitive (TTX-S), and others such as NaV1.8 being resistant to the toxin (TTX-R). Using immunocytochemistry, we previously found evidence for NaV1.6–NaV1.9 channel expression in isolated muscle afferent neurons, but not NaV1.1. 9 However, using TTX and other NaV channel blockers, our electrophysiological results supported NaV1.8 as the dominant NaV channel in muscle afferent somata.9,10
While TTX minimally affected somatic NaV currents (average 14% block), 9 axonal action potentials from both group III and IV afferents were almost completely blocked by TTX applied to the dorsal roots, 11 which supports little, if any, role in action potential generation by TTX-R channels, such as NaV1.8. So, we were interested in knowing if NaV1.8 channels are expressed in group IV fibers. In addition, since TTX cannot distinguish different TTX-S NaV channels, we wanted to determine which TTX-S channels are expressed by these afferent neurons. To better understand the NaV channel distribution patterns, we compared the expression of these channels by immunohistochemical staining of the muscle and DRG sections. Group IV neurons and axons were identified by labeling with a peripherin antibody.12,13 We show that NaV1.6, NaV1.7, and NaV1.8 channels are expressed in >67% of muscle afferent axons and somata. While there were no significant differences in the percentage of axons vs. somata labeled by the NaV1.6 and NaV1.7 antibodies, the antibody targeting NaV1.8 channels labeled significantly fewer group IV axons than somata, which fit well with the electrophysiological data. It seems possible that NaV1.8 could play a role in action potential generation within group IV axons. However, the conditions under which that could occur have yet to be identified.
Experimental procedures
All animal protocols were approved by the Kirksville College of Osteopathic Medicine IACUC and were consistent with the National Research Council guide for the Care and use of Laboratory Animals. Adult male Sprague-Dawley rats (Hilltop Lab Animals, Scottsdale, PA) weighing between 150 and 400 g were used in these experiments. Rats were housed in an USDA-Approved, AAALAC-Certified Animal Care facility at a constant temperature of 24 ± 1℃, controlled 12-h light dark cycles and access to commercial rat chow and water ad libitum.
Muscle preparation
Rats were sacrificed using CO2 followed by decapitation using a laboratory guillotine (Kent Scientific Corp, Torrington, CT). 9 Both gastrocnemius muscles were dissected out along with the tendons, washed in ice-cold PBS solution, and flash frozen in dry ice-cooled isopentane. The muscles were kept frozen at −80℃ until use. Frozen muscles were cut longitudinally in 25 µm sections using a Leica CM1900 cryostat (Leica Microsystems Inc., Buffalo Grove, IL). The sections were mounted on polylysine-coated slides, allowed to dry and stored at −20℃ until use.
DRG preparation
Rats were anesthetized with an intraperitoneal injection of a mixture of ketamine, xylazine, and acepromazine, and both right and left gastrocnemius muscles were injected with 100 µl of 1,1-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (DiI; Life Technologies, Grand Island, NY) solution (1.5% in DMSO) as previously described. 9 Four to five days were allowed for transport of DiI from the muscle to the somata of muscle afferents in the DRG. Rats were sacrificed and the Lumbar 4 and 5 DRG were dissected out, frozen, and sectioned as described above.
Superior cervical ganglia preparation
Superior cervical ganglia (SCG) were dissected from sacrificed rats (as described for muscle preparation), frozen, and sectioned as described above to be used as control tissue to test the antibodies.
Staining
List of antibodies used for labeling.

Quantification
Nerve fibers that were at least 100 µm long and positive for peripherin were counted and checked for colabeling by NaV antibodies. The data are expressed as the percentage of peripherin labeled fibers that were positively stained by the NaV antibody. Peripherin negative fibers were not counted. The data were compiled from gastrocnemius muscles from 3–4 rats for each NaV antibody.
For DRG and SCG neuronal counts, images were captured at 10×, fluorescence intensity and cell size were measured from individual neurons using ImageJ (rsbweb.nih.gov/ij/index.html). The fluorescent intensity of unlabeled neurons was measured and plotted in a histogram to determine the threshold for positively labeled neurons as previously described. 9 The DRG somata that were positively labeled with DiI and the peripherin antibody were considered as group IV muscle afferent neurons.
Results
To better understand the NaV channels expressed in muscle afferent axons, we used immunohistochemistry to identify the expressed channels. We visualized group IV afferents by staining muscle sections with an antibody against peripherin, which is a neurofilament protein that is selectively expressed in unmyelinated (e.g. group IV) sensory axons. 12 For comparison with our electrophysiological data, we also examined NaV channel expression in DRG sections. Muscle afferent neurons within these sections were identified by retrograde labeling using DiI that was injected into the gastrocnemius muscle, and group IV neurons were further identified by positive peripherin antibody labeling.
We previously defined group IV neurons as those with cell body diameters <30 µm.9,10 This definition was tested by calculating the diameters of DiI and peripherin labeled (i.e. group IV) somata in the DRG sections. The diameters ranged from 16 to 35 µm with an average of 23 ± 4 µm with 96% of group IV neurons having a diameter <30 µm. (Figure 1). These results support our inclusion of neurons with diameters <30 µm in group IV.
Neuronal diameter histogram of group IV neurons. Neurons from DRG sections that were labeled by both DiI and the peripherin antibody were counted as group IV.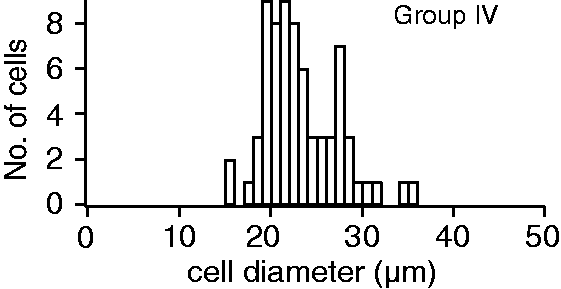
NaV1.8 channels in group IV afferent fibers
Previous electrophysiology experiments supported a dominant role for NaV1.8 channel activity in group III and IV somata,
9
but a minimal role in the axons.
11
This suggested that NaV1.8 channels may not be expressed in group IV axons. However, we found that group IV axons within gastrocnemius muscles were positively labeled by the NaV1.8 channel antibody (Figure 2(a)). Labeling was found in individual group IV nerve fibers (Figure 2(a)) and in group IV fibers within nerve tracts (Figure 2(b)). No labeling was observed in control slides (Figure 2(c)), which was true for all experiments regardless of the antibody used (not shown). Individual peripherin-positive fibers were counted to determine the percentage of those fibers labeled by the NaV1.8 antibody. On average, we found that 67 ± 1% (mean ± SD from three muscles) of the peripherin-positive fibers were labeled by the NaV1.8 channel antibody (Table 2). The percentage of NaV1.8-positive group IV fibers ranged from 66% to 68% for the three muscles tested with a total of 156 fibers counted (34–63 fibers/muscle). Each muscle came from a different animal. Interestingly, the counts from DRG sections showed that 96 ± 7% (20/21) of group IV somata were labeled by the NaV1.8 channel antibody, which was significantly higher than for group IV fibers (Table 2). Three DRG sections from each of three different animals were counted (total = 9 sections). These results support the electrophysiological results that showing NaV1.8 channels play a larger role in somatic vs. axonal Na+ influx.9,11
NaV1.8 antibody labeling in muscle and DRG sections. Muscle and DRG sections were colabeled with a mouse NaV1.8 channel antibody and a chicken peripherin antibody. NaV1.8 channels are present in peripherin-positive group IV nerve fibers (a) and also in the nerve bundles (b). NaV1.8 channel antibody also labeled some of the peripherin-negative fibers (arrow). (c) Muscle sections exposed only to the secondary antibodies (control) showed no labeling, which was consistently found in all experiments. (d) DRG sections showing that group IV neurons (colabeled with DiI and the peripherin antibody) are also labeled by the NaV1.8 antibody (arrows). (e) DRG sections exposed only to the secondary antibodies (negative control) showed only DiI labeled neurons. Scale = 50 µm. NaV antibody labeling of group IV afferents. Significant difference between somal and axonal labeling.

TTX-S NaV channels in group IV axons and somata
Previous electrophysiology studies demonstrated that muscle afferent somata
9
and axons
11
express TTX-S NaV channels. In addition, we showed that antibodies directed against NaV1.6 and NaV1.7 channels could label isolated muscle afferent neurons.
9
However, the expression of these TTX-S NaV channels in muscle afferent axons has not been investigated. We colabeled muscle afferents with peripherin and antibodies targeting NaV1.1, NaV1.6, or NaV1.7 channels.
7
Similar to our previous study of muscle afferent somata,
9
we failed to find any NaV1.1-positive fibers (Figure 3). Since we did not find NaV1.1 channel labeling in either cultured cells or muscle afferent axons, we did not further investigate this channel in DRG sections.
An antibody against NaV1.1 channels fails to label muscle afferent axons in gastrocnemius muscle. Peripherin-positive group IV axons are shown, but we never found muscle afferent fibers co-labeled by the NaV1.1 channel antibody. Scale = 50 µm.
NaV1.6 channels are widely expressed in sensory neurons, including unmyelinated sensory fibers.14,15 The NaV1.6 channel antibody positively labeled 84 ± 9% of peripherin-positive fibers (Table 2; Figure 4(a) and (b)). For the gastrocnemius muscles from the three animals tested, the percentage of NaV1.6-positive group IV axons ranged from 74% to 93% from a total of 208 group IV fibers counted. As with NaV1.8, NaV1.6 channel labeling was found in both individual group IV fibers and those within nerves (Figure 4(a) and (b)). We consistently saw a light, diffuse muscle fiber staining by the NaV1.6 channel antibody, which may result from non-specific myosin labeling.
16
In the DRG sections, 98 ± 4% (21/22) of the muscle afferent somata were labeled by the NaV1.6 channel antibody (Figure 4(c)). There was no statistical difference between the percentages of NaV1.6-positive group IV axons vs. somata (Table 2).
A NaV1.6 channel antibody labels group IV fibers in muscle sections and neurons in DRG sections. Gastrocnemius and DRG sections were colabeled with a rabbit NaV1.6 channel antibody and a chicken peripherin antibody. NaV1.6 channels were present in peripherin-positive group IV nerve fibers (a, b). Some of the peripherin-positive fibers are not stained positive for NaV1.6 (a, upper right corner). (c) Most DRG group IV somata are labeled by the NaV1.6 channel antibody (arrows). Scale = 50 µm.
NaV1.7 channel activity is important for action potential generation and propagation in nociceptors.17,18 Within the peripherin-positive group IV fibers, we found that 77 ± 6% were labeled by the NaV1.7 antibody (Table 2; Figure 5), which included both individual fibers (Figure 4(a)) and fibers within axon bundles (Figure 4(b)). For gastrocnemius muscles from the three animals tested, between 71% and 83% of group IV fibers were labeled by the NaV1.7 antibody with a total of 185 group IV axons counted. As with NaV1.6, we saw a diffuse labeling of muscle fibers with the NaV1.7 antibody, which may result from non-specific myosin labeling.
16
Group IV fibers and neurons are labeled by a NaV1.7 channel antibody. Gastrocnemius and DRG sections were colabeled with a mouse NaV1.7 channel antibody and chicken peripherin antibody. NaV1.7 channels are expressed in by most individual group IV axons (a) as well as those in nerve bundles (b). However, some group IV axons failed to show evidence of NaV1.7 channel labeling (panel a). (c) The majority of peripherin-positive, DiI-positive (group IV) neurons were also positive for NaV1.7 channel antibody labeling (arrows). Scale = 50 µm.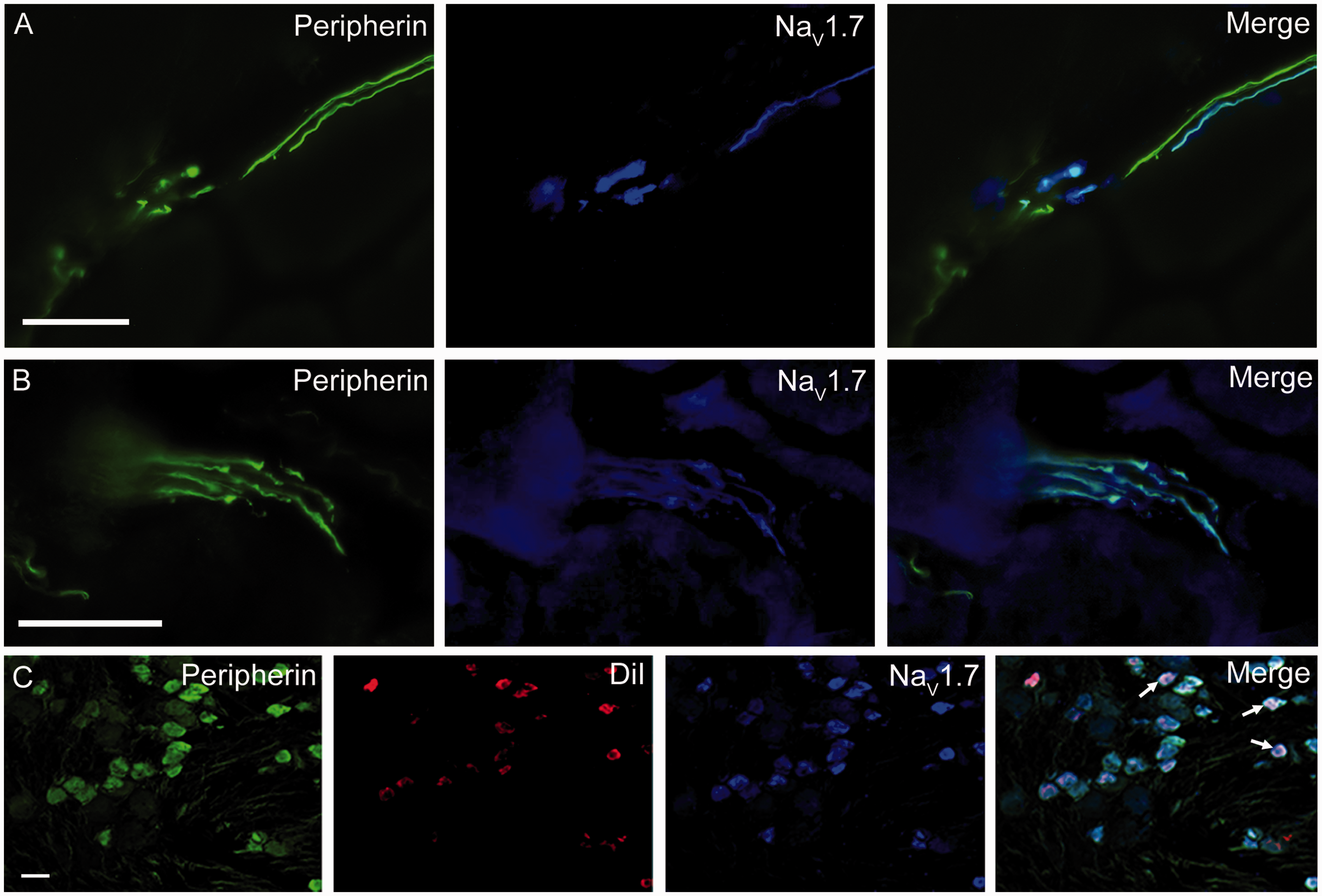
In the DRG sections, 82 ± 20% (23/27) of the group IV somata were NaV1.7-positive, which was not significantly different from the percentage of NaV1.7-positive group IV fibers. As a result of the high percentage of axons expressing these TTX-S channels, it is likely that both NaV1.6 and NaV1.7 channels contribute to the TTX-S Na+ influx in both group IV axons and somata.
SCG labeling
As a test for NaV antibody specificity, we examined antibody labeling of SCG neurons, which express NaV1.6 and NaV1.7, but not NaV1.8 channels.19,20 Symphathetic neurons are B type, so the peripherin antibody did not label these cells (Figure 6 (a), (c), and (e)). For NaV antibody labeling, the NaV1.6 antibody labeled 91% (Figure 6(b) and (g)) and NaV1.7 antibody labeled 95% of neurons in the SCG (>100 cells for each antibody tested) (Figure 6(d) and (h)). Histograms of fluorescent intensity show peaks that were statistically distinct from baseline labeling (Figure 6 (g) and (i)). However, the histogram for NaV1.8 antibody labeling shows complete overlap with background, which demonstrates no labeling by this antibody (Figure 6(f) and (h)), as expected.
20
SCG labeled with NaV antibodies. As expected, the peripherin antibody failed to label SCG neurons or fibers (a, c, e). NaV1.6 channel antibody labeled most neurons. Note that some neurons were labeled more intensely than others (b). Almost all cells showed intense labeling by NaV1.7 channel antibody (d). NaV1.8 channel antibody failed to label any SCG neurons (f). Histograms of the fluorescent intensity show background (Bkgd, black bars) and the NaV antibody labeling (red bars). The histograms were fit using a Gaussian equation (g–i). At least 100 SCG neurons were counted for each NaV antibody tested.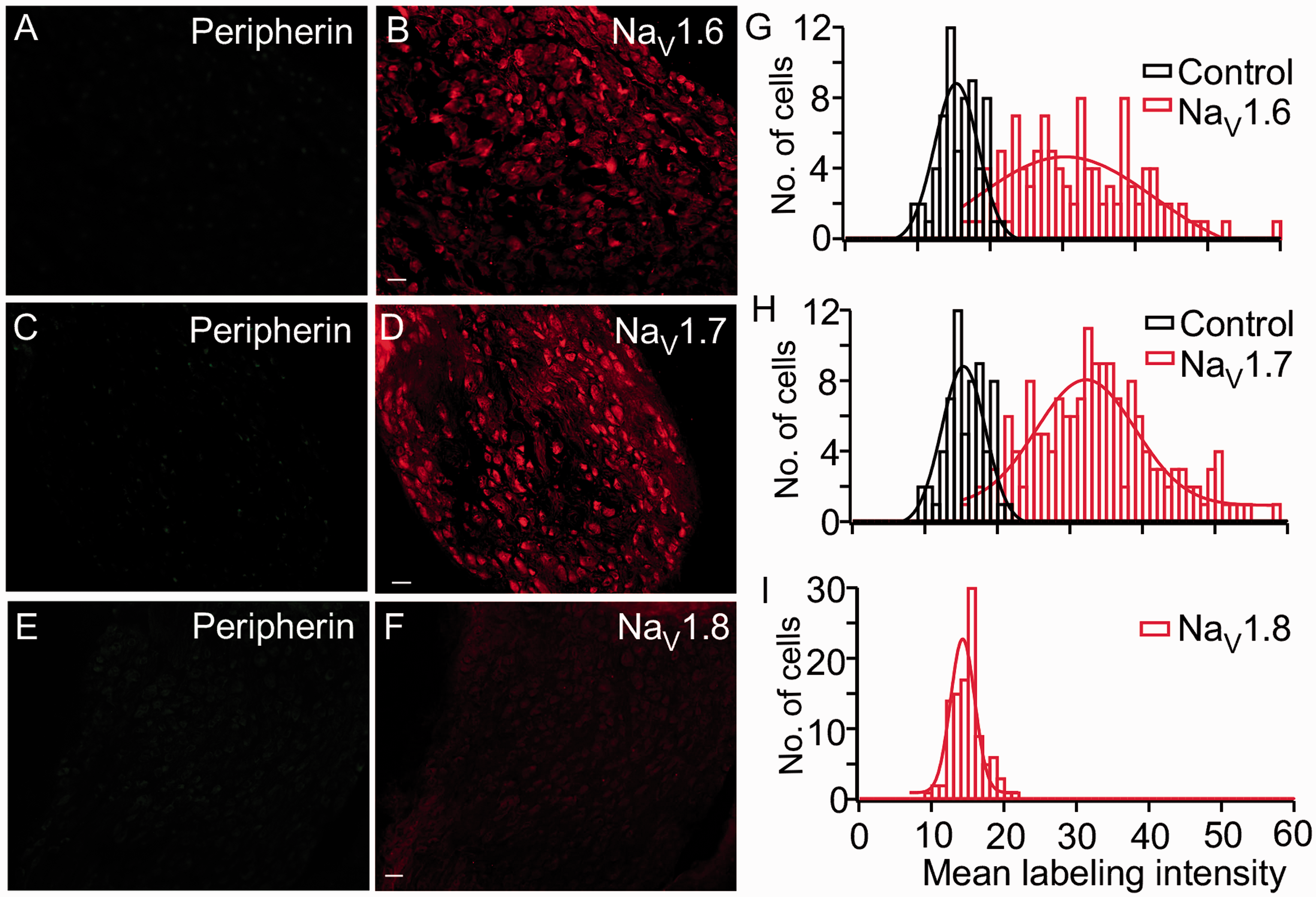
Discussion
The NaV isoforms that generate action potentials in group IV afferents are not known, but previous reports suggested that the dominant isoforms could differ between muscle afferent somata vs. axons.9,11 Using the peripherin antibody, we were able to label a subset of axons within rat gastrocnemius muscles, which based on previous publications we identified as unmyelinated group IV afferents.12–14,21 The majority of group IV afferent axons were positively labeled by antibodies to either NaV1.8 (67%), NaV1.6 (83%), or NaV1.7 (77%) channels. A diffuse muscle staining was observed with both the NaV1.6 and NaV1.7 channel antibodies. While both NaV1.6 and NaV1.7 channels are reported to be expressed in non-neuronal tissues,22–25 these channels have not been found in skeletal muscle fibers.26,27 NaV1.7 channel antibodies have been previously observed to label skeletal muscle proteins, but this was interpreted as non-specific labeling of myosin, since the labeling was not changed when the antibody was pre-blocked with peptide. 16 Thus, the diffuse muscle labeling likely results from non-specific absorption of these antibodies by myosin. 16
Within the DRG, a significantly larger percentage of group IV somata were labeled by the NaV1.8 channel antibody relative to afferent fibers, but there was no statistical difference between axonal and somata labeling for antibodies to either NaV1.6 or NaV1.7 channels. Given the high percentage of NaV-positive fibers and somata, it appears that a majority express at least two of these NaV channel isoforms, and many may express all three. If these channels are functional in the axon, it is likely that all three participate in AP generation within group IV axons to signal muscle pain and the EPR.
NaV1.1
We have been unable to produce evidence for NaV1.1 channel expression in muscle afferents, including group IV muscle afferent fibers and muscle afferent somata. 9 These findings are not surprising since NaV1.1 channel antibodies failed to label cutaneous afferents and only weak labeling was observed in isolated somata. 21 mRNA for NaV1.1 channels can be detected in sensory afferents,28,29 which suggests that these channels could be produced. However, NaV1.1 channel levels may be too low to be detected in our assay. Alternatively, NaV1.1 channels have been reported to be only expressed in large diameter myelinated sensory neurons, 30 which were not studied here. A study using conotoxins that blocked a range of NaV channels concluded that NaV1.1 channels did not significantly contribute to action potential generation in either C-fiber (group IV) or Aδ-fibers (group III) in sciatic nerve. 31
NaV1.6
Several reports have demonstrated that TTX-S NaV1.6 channels are present at the nodes of Ranvier in myelinated nerve fibers and play a role in saltatory conduction, 32 but NaV1.6 channel expression has also been identified in unmyelinated sensory neurons and may play an important role in pain signaling. Indeed, previous reports showed that 100% of peripherin-positive fibers within sciatic nerve were positive for NaV1.6 channels.14,30 Similarly, we found NaV1.6 expressed in 84% of the group IV (unmyelinated) nerve fibers and 96% of somata. However, the role in group IV action potential generation may be limited. The transient knockdown of NaV1.6 channels was found to have only a small effect on C-fiber excitability, while strongly reducing Aδ fiber excitability. 15 In addition, NaV1.6-targeted conotoxins failed to affect compound action potentials in C-fibers (group IV) but completely blocked Aδ-fiber (Group III) action potentials in sciatic nerve. 31 Thus, the loss of functional NaV1.6 channels does not appear to change excitability of unmyelinated sensory afferents (C-fibers and group IV fibers). Interestingly, NaV1.6 knockdown reduced pain behavior in the animals, which led to the conclusion that NaV1.6 could be an important analgesic drug target. 15 The role of NaV1.6 channels in controlling group IV axonal excitability requires further investigation.
NaV1.7
The role of NaV1.7 channels in human nociception has been well established based on the effects of NaV1.7 channel mutations on human pain perception.18,30,33–35 Therefore, it is not surprising that we found the majority of group IV neurons and fibers labeled by the NaV1.7 channel antibody. However, a previous study showed a lower percentage of NaV1.7-positive C-fibers in sciatic nerve (27%) vs. DRG neurons (63%), 13 and these percentages are lower than we found for group IV fibers (77%) and neurons (82%). The source of these differences is not clear, but some possibilities include differences (1) in NaV channel expression between cutaneous and muscle afferents, (2) in NaV channel expression between terminal fibers and those within the sciatic nerve, and (3) technical differences perhaps involving NaV antibody penetration of individual fibers vs. those within the sciatic nerve. The available evidence appears to support the third possibility since toxins that target NaV1.7 channels strongly inhibit the C-fiber component of the compound action potential.31,36 The strong expression by group IV afferents suggests NaV1.7 channels will play an important role in muscle pain and the EPR. Indeed, the NaV1.7 channel blocker Ssm6a significantly suppressed the muscle contraction-induced pressor effect mediated by the EPR. 36
NaV1.8
The TTX-R NaV1.8 channels are dominantly expressed in a majority of muscle afferent somata, 9 and these channels have been shown to play an important role in action potential generation in nociceptor somata.7,37,38 Thus, it is possible that NaV1.8 channels play a role in muscle nociception.21,39 We found 67% of the group IV fibers were positively labeled with the NaV1.8 antibody, which is a significantly smaller percentage than for group IV somata (96%). This is in line with electrophysiological studies showing that NaV1.8 channels provide the dominant pathway for Na+ influx in muscle afferent neurons 9 but may not have sufficient axonal activity to support action potential propagation in group IV afferents. 11 Indeed, muscle specific group III and IV signals are almost completely blocked by TTX, 11 which indicates that NaV1.8 channels play a minimal role.
Although not a focus of this study, we found that non-peripherin labeled fibers could also be labeled by the NaV1.8 antibody. This supports the presence of NaV1.8 channels in non-group IV afferents and reinforces our earlier studies that supported expression of NaV1.8 channels in small, medium, and large diameter afferent neurons.9,10
Using immunohistochemistry, NaV1.8 channels have been previously shown to be expressed by the majority of sensory free nerve endings in the skin. 21 Indeed, 96% of these cutaneous endings were positive for NaV1.8 channels, which is a higher percentage than we found in muscle. However, we used peripherin to selectively examine unmyelinated sensory fibers, while a PGP9.5 antibody were used to identify afferent axons in the skin, 21 and PGP9.5 is a marker for both unmyelinated and myelinated sensory afferents. 40 Thus, one possible explanation for the higher percentage of labeled cutaneous sensory fibers is that NaV1.8 channels are more highly expressed by myelinated axons. An additional possibility is that NaV1.8 channels are indeed expressed by a higher percentage of cutaneous afferent fibers than muscle afferent fibers. One potential reason for such an difference is that NaV1.8 channel activity helps to sustain action potential firing at lower temperatures, 41 and the skin is likely to experience much larger temperature changes than muscle.
NaV channels mediating muscle pain
Pathological conditions have been shown to alter the expression and axonal distribution of NaV channels, 42 which could be important for mediating muscle afferent hyperexcitability. Indeed, NaV1.7 and NaV1.8 channels are involved in inflammatory pain, 43 and it is known that mutations of these channels are associated with neuropathic pain in human patients.34,44 It is interesting that hyperexcitability resulting from a mutant NaV1.7 channel has been found to require NaV1.8 channel expression. 20 The NaV1.7 channel mutant, L858H, produces pain in response to mild warmth in humans and induces hyperactivity when expressed in DRG, but not in SCG. 20 However, coexpressing NaV1.8 in SCG along with L858H produces SCG hyperactivity. It is thought that the more depolarized voltage dependence of activation and inactivation of NaV1.8 channels 38 is a major determinant factor in producing nociceptor hyperactivity and thermal allodynia in patient carrying L858H mutation. 20
The NaV1.8 channel expression has been shown to be increased in PGP9.5 labeled muscle nerve fibers in patients with chronic myofascial temporomandibular disorders. 45 However, a claudication animal model (peripheral vascular disease-reduced muscle blood flow) produced no changes in TTX sensitivity of group III/IV afferent activity, 11 which suggests that TTX-R channel activity (i.e. NaV1.8) is not upregulated under muscle ischemia conditions. Potential changes in TTX-S channel activity (e.g. NaV1.7) have not yet been investigated using this animal model. One caveat is that the ischemia duration produced by this animal model is three days, which may be too short to observe changes in axonal NaV channel expression. Previous work showing changes in NaV channel expression were done seven days following nerve injury.42,46 Thus, it is possible changes in group III/IV NaV channel expression could be observed in a longer duration muscle ischemia model.
Group IV fibers carry the action potentials that signal the EPR and muscle pain, 5 and the evidence supports the involvement of multiple NaV channels in action potential generation in pain fibers, 21 as well as other sensory afferent fibers.47–49 A large fraction of group IV fibers express the NaV channels, with 67% expressing NaV1.8 channels, 84% expressing NaV1.6 channels, and 77% expressing NaV1.7 channels. Thus, it is highly likely that the majority of group IV fibers expressed at least two NaV channel isoforms, and the expression of all three channel isoforms within single fibers could be common. Although we have not determined which of these channels participate in action potential generation, recent studies from the Kaufman group supports the involvement of multiple TTX-S channels, with a minimal role for TTX-R NaV1.8 channels. Muscle afferent signals that generate the contraction-induced pressor response are almost completely blocked by TTX, while the NaV1.8-specific blocker A803467 had little or no significant effect. 11 The NaV1.7 channel blocker, Ssm6a, blocked these muscle specific signals by ∼50%, 36 which, combined with the TTX results, suggests a substantial role for another TTX-S channel type, such as NaV1.6. One issue is that the contraction-induced pressor signals are carried by both group III and IV neurons, so it is possible that the Ssm6a resistant signals are mediated by group III axons. However, Ssm6a partially blocked the compound action potentials recorded from both group III-Aδ and group IV-C fibers, 36 which supports NaV1.7 channel expression in both small myelinated (group III-Aδ) and unmyelinated axons (group IV-C). Thus, it appears that both NaV1.6 and NaV1.7 channel participate in action potential generation in both group III and IV afferents, but NaV1.8 channels, while present, may not have sufficient density to sustain action potential generation under normal conditions.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by a National Institute of Arthritis and Musculoskeletal and Skin Diseases Grant AR-059397 (K. S. Elmslie).