
Abstract
Background
Papillary thyroid carcinoma is the most frequent histological subtype of thyroid cancer with a high incidence. We aimed to explore the function of circular RNA_0039411 (circ_0039411) and its associated mechanism in papillary thyroid carcinoma progression.
Methods
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and Western blot assay were conducted to determine the expression of RNA and protein, respectively. The colony formation ability, migration, invasion, and apoptosis were analyzed by colony formation assay, transwell migration assay, transwell invasion assay, and flow cytometry. Cell glycolytic metabolism was analyzed using fluorescence-based glucose assay kit and fluorescence-based lactate assay kit. Dual-luciferase reporter assay and RNA-Pull-Down Assay were performed to validate the binding between microRNA-423-5p (miR-423-5p) and circ_0039411 or SRY-box transcription factor 4 (SOX4). The xenograft tumor model was used to assess the role of circ_0039411 in the tumor growth in vivo.
Results
Circ_0039411 was highly expressed in papillary thyroid carcinoma tissues and cell lines compared with adjacent normal tissues and NTHY-ORI3.1 cells. Circ_0039411 interference suppressed the colony formation ability, migration, invasion, and glycolysis but promoted the apoptosis of papillary thyroid carcinoma cells. MiR-423-5p was a target of circ_0039411 in papillary thyroid carcinoma cells. Circ_0039411 knockdown-mediated effects in papillary thyroid carcinoma cells were largely overturned by the silence of miR-423-5p. MiR-423-5p bound to the 3′ untranslated region (3′UTR) of SOX4. SOX4 overexpression largely reversed circ_0039411 silencing-mediated effects in papillary thyroid carcinoma cells. Circ_0039411 positively regulated SOX4 expression by sponging miR-423-5p in papillary thyroid carcinoma cells. Circ_0039411 silencing notably suppressed the growth of xenograft tumors in vivo.
Conclusion
Circ_0039411 promoted the malignant behaviors of papillary thyroid carcinoma cells partly depending on the regulation of the miR-423-5p/SOX4 axis.
Introduction
Papillary thyroid carcinoma (PTC) is the most frequent histological subtype of thyroid cancer, and is the most prevalent malignancy of the endocrine system.1,2 The prognosis of PTC patients is usually satisfactory because of the steady growth rate of the tumor and the retention of the patient's iodine uptake. 3 However, some PTC patients with invasiveness and metastasis in the early stage are prone to have a dismal outcome after surgery. 4 Accumulating studies have focused on the clinical diagnosis of PTC metastasis.5‐7 In this study, we explored the potential mechanism behind PTC pathology using approaches in vitro and in vivo.
Non-coding genes make up the majority of the human genome, 8 and circular RNAs (circRNAs) are a special class of non-coding transcripts with a closed-loop structure. 9 With the development of sequencing techniques, an increasing number of circRNAs have been found to be dysregulated in human malignancies.10,11 CircRNAs are reported to regulate various biological processes related to PTC progression including cell proliferation, migration, invasion, and glycolysis.12‐15 A previous study reported that circ_0039411 was up-regulated in PTC, and it aggravated PTC progression. 16 In this study, the working mechanism of circ_0039411 in PTC development was further explored.
CircRNAs can regulate tumor progression by acting as miRNA sponges. 17 MiRNAs have shown important regulatory roles in cancer development. 18 Based on the bioinformatic analysis, we found that miR-423-5p possessed the potential binding sites with circ_0039411. A previous study found that miR-423-5p expression was reduced in PTC cells, and low expression of miR-423-5p restrained the malignant behaviors of PTC cells. 19 In this study, we examined the binding between circ_0039411 and miR-423-5p in PTC cells and explored their functional association in regulating PTC progression.
MiRNAs regulate gene expression in post-transcriptional level by binding to target mRNAs at 3′ untranslated region (3′UTR). 20 The binding between miR-423-5p and SRY-box transcription factor 4 (SOX4) was predicted by bioinformatic database. SOX4 is a transcription factor that activates or suppresses gene transcription by functioning alone or together with other transcription factors.21,22 Previous studies have reported the oncogenic role of SOX4 in several malignancies.23‐25 However, the role of SOX4 in PTC development has never been clarified. In the present study, the target relation between miR-423-5p and SOX4 in PTC and their functional association were explored.
In this study, we first analyzed the expression pattern and function of circ_0039411 in PTC cells. Subsequently, the working mechanism of circ_0039411 was explored via bioinformatic analysis and rescue experiments. Finally, the in vivo role of circ_0039411 in the tumor growth was assessed using the xenograft tumor model.
Materials and methods
Clinical samples
A total of 51 pairs of PTC tissue specimens and adjacent healthy tissue specimens were collected from PTC patients at Beijing Chao-yang Hospital, Capital Medical University. All tissues were immediately frozen in liquid nitrogen and stored at −80°C. Tumor tissues were utilized to determine the expression of circ_0039411, miR-423-5p and SOX4 messenger RNA (mRNA) and protein via reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and Western blot assay. All participants have signed written informed consent. This clinical study obtained permission from the Ethics Committee of Beijing Chao-yang Hospital, Capital Medical University.
Cell lines
PTC cell lines (TPC-1 and BHT-101) and human normal thyroid cell line NTHY-ORI3.1 purchased from BeNa Culture Collection (Beijing, China) were cultured with Dulbecco's modified Eagle's medium (DMEM) medium (Gibco, Carlsbad, CA, USA) plus 10% fetal bovine serum (FBS, Gibco) and 1% antibiotic reagent (Sangon Biotech, Shanghai, China) at 37°C with 5% CO2.
RT-qPCR
RNA samples were isolated from PTC tissues and cells using Trizol reagent (Invitrogen, Carlsbad, CA, USA). Complementary DNA (cDNA) was synthesized using TaqMan Reverse Transcription Reagent (Invitrogen). Thermal-cycling reaction was conducted using Real Master Mix (SYBR Green) (Tiangen, Beijing, China) and specific primers (Supplementary Table 1) on BioRad CFX96TM (Bio-Rad, Hercules, CA, USA). The relative expression of circ_0039411 and SOX4 was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and miR-423-5p level was normalized to U6. The relative expression was analyzed by the 2−ΔΔCt method.
Cell transfection
Small interfering (si)RNA targeting circ_0039411 (si-circ_0039411), siRNA negative control (si-NC), short hairpin (sh)RNA targeting circ_0039411 (sh-circ_0039411), shRNA negative control (sh-NC), miR-423-5p mimics (miR-423-5p), negative control of miRNA (miR-NC), miR-423-5p inhibitor (anti-miR-423-5p), anti-NC, SOX4 overexpression plasmid (SOX4), and empty vector (Vector) were purchased from GenePharma (Shanghai, China) and Ribobio (Guangzhou, China). The oligonucleotides or plasmids were transfected into PTC cells using the Lipofectamine 3000 (Invitrogen).
Colony formation assay
PTC cells were plated at 200 cells/well in 12-well plates in triplicate, and were then incubated for 2 weeks to form colonies. The colonies were stained with 10% Giemsa solution (Sangon Biotech), and colonies that contained more than 50 cells were counted.
Transwell migration and invasion assays
Diluted Matrigel solution (Sigma, St. Louis, MO, USA) was added to the upper chambers for solidification to mimic the extracellular matrix to analyze cell invasion ability, and un-coated upper chambers were utilized to analyze cell migration ability. PTC cells were starved for 12 h. PTC cells suspended in 100 μL serum-free medium were seeded into the upper chambers, and the bottom chambers were filled with 600 μL culture medium with 10% FBS. Cells passed through the membrane were stained with 10% Giemsa solution (Sangon Biotech) and migrated, and invading cells were counted under an optical microscope (Olympus, Osaka, Japan) at 100×.
Flow cytometry
PTC cells were collected and suspended in the binding buffer (BD Biosciences, Franklin, NJ, USA). Subsequently, Annexin V-fluorescein isothiocyanate (Annexin V-FITC; BD Biosciences) and propidium iodide (PI; BD Biosciences) were added to simultaneously incubate with PTC cells in the dark. The apoptosis rate (FITC+ and PI+/−) was analyzed by the FACS CantoII flow cytometer (BD Biosciences).
Western blot assay
PTC tissues and cells were lysed using the Radioimmunoprecipitation assay (RIPA) buffer (Beyotime, Haimen, China). Equal amounts of protein samples (30 μg) were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and were then blotted onto a polyvinylidene difluoride (PVDF) membrane (Bio-Rad). After sealing with 5% non-fat milk, the membrane was incubated with the primary antibodies including anti-Cyclin D1 (ab16663; Abcam, Cambridge, MA, USA), anti-Vimentin (ab92547; Abcam), anti-BCL2 associated X, apoptosis regulator (anti-Bax, ab32503; Abcam), anti-SOX4 (ab70598; Abcam), and anti-GAPDH (ab8245; Abcam) overnight. Horseradish peroxidase (HRP)-labeled secondary antibody (Abcam) at the dilution of 1:5000 was then incubated with the membrane for 2 h. The protein signals were visualized using the enhanced chemiluminescent (ECL) kit (Thermo Fisher Scientific, Waltham, MA, USA). The protein expression was quantified using the Image Lab analysis software (Bio-Rad).
Glycolytic measurement
The intracellular level of glucose and the level of lactate in culture medium were measured using fluorescence-based glucose assay kit (BioVision, Milpitas, California, USA) and fluorescence-based lactate assay kit (BioVision) according to the manufacturer's instructions.
Dual-luciferase reporter assay
Luciferase plasmids (circ_0039411 WT, circ_0039411 MUT, SOX4 3′UTR WT, and SOX4 3′UTR MUT) were constructed through inserting the wild-type or mutant type fragment of circ_0039411 or SOX4 3′UTR into pmirGLO plasmid (Promega, Madison, WI, USA). MiR-423-5p or its control miR-NC was co-transfected into PTC cells with the luciferase plasmids. After transfection for 24 h, luciferase intensities were examined using the Dual-Luciferase Reporter Assay System kit (Promega).
RNA-Protein Pull-Down assay
RNA-Protein Pull-Down Kit (Thermo Fisher Scientific) was used to verify the binding between miR-423-5p and circ_0039411. PTC cells were transfected with Bio-miR-NC or Bio-miR-423-5p. Cell lysates were incubated with dynabeads, and enriched RNAs were analyzed by RT-qPCR.
Xenograft tumor model
A total of ten BALB/c male nude mice purchased from Vital River Laboratory Animal Technology (Beijing, China) were arbitrarily divided into sh-NC group (n = 5) and sh-circ_0039411 group (n = 5). TPC-1 cell line stably transfected with sh-NC or sh-circ_0039411 was established. A total of 2 × 106 TPC-1 cells were subcutaneously injected into the right flank of the mice. Tumor volume was measured every week as length × width2 × 0.5. At 35 d post-injection, the tumors were resected and weighed. Immunohistochemistry (IHC) assay was conducted to measure the level of Ki-67 using its antibody (ab15580; Abcam) at the concentration of 1 μg/mL. 26 Animal experiments were approved by the Animal Research Ethics Committee of Beijing Chaoyang Hospital, Capital Medical University.
Data analysis
The data from three independent experiments were analyzed using Graphad Prism 7.0 software (GraphPad, La Jolla, CA, USA) and were represented as mean ± SD. Differences in the two groups were analyzed using the Student's t-test, and differences in multiple groups were analyzed using one-way analysis of variance (ANOVA). P < 0.05 was considered statistically significant.
Results
Circ_0039411 expression is up-regulated in PTC tissues and cell lines
Circ_0039411 is derived from the host gene matrix metallopeptidase 2 (MMP2), and its position in chromosome was shown in Figure 1(a). The expression pattern of circ_0039411 in PTC was analyzed. Circ_0039411 expression was significantly up-regulated in PTC tissues (n = 51) compared with paracancer normal tissues (n = 51) (Figure 1(b)). We found that circ_0039411 level was up-regulated in two PTC cell lines (TPC-1 and BHT-101) compared with normal human thyroid cell line NTHY-ORI3.1 (Figure 1(c)). Aberrant expression of circ_0039411 in PTC tissues and cell lines might imply its vital regulatory role in PTC progression.
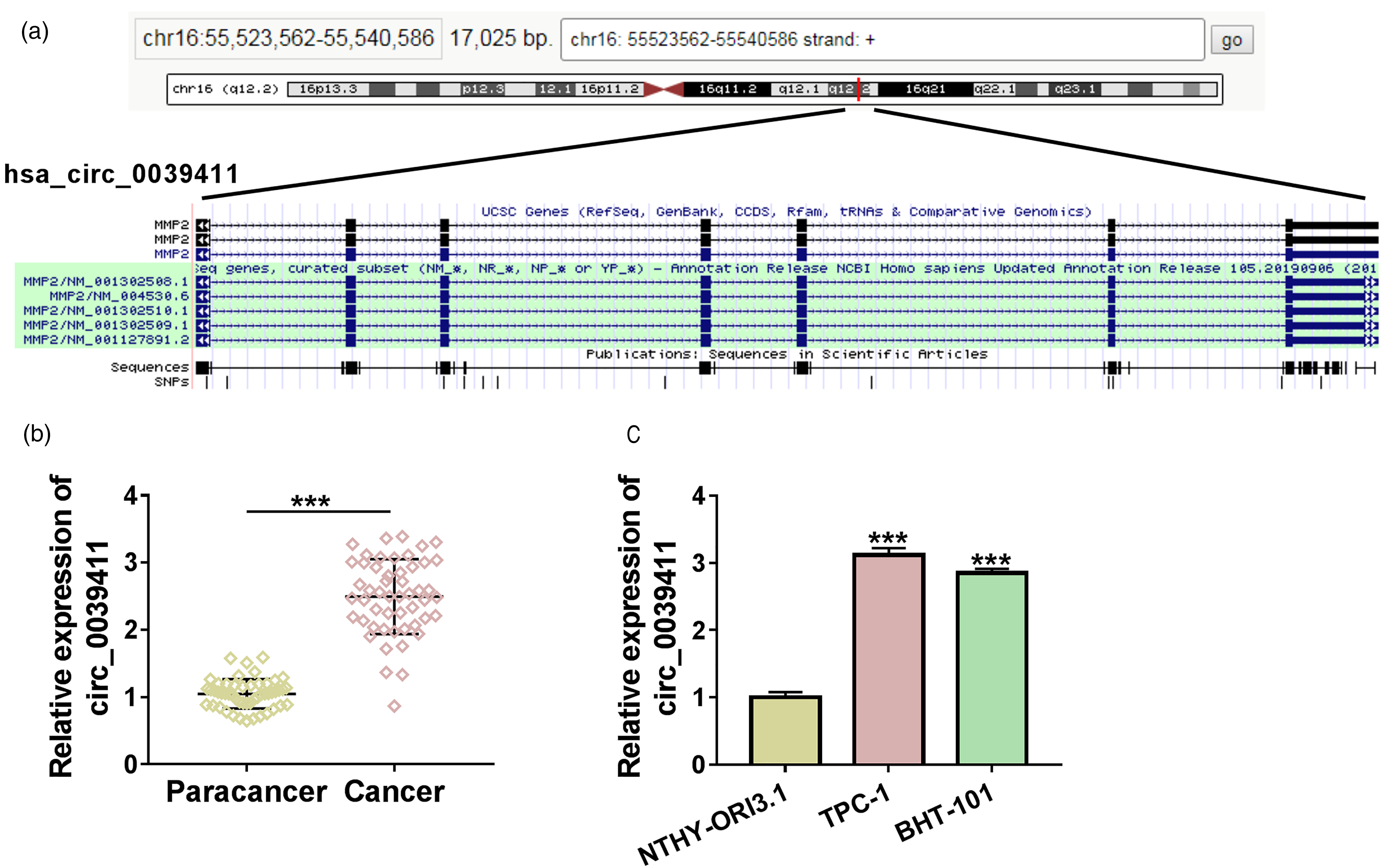
Circ_0039411 expression is up-regulated in PTC tissues and cell lines. (a) Circ_0039411 is derived from the back-splicing of MMP2 gene, and its chromosomal position is shown. (b) RT-qPCR was applied to determine the expression of circ_0039411 in PTC tissues (n = 51) and paracancer tissues (n = 51). (c) The level of circ_0039411 was examined in two PTC cell lines (TPC-1 and BHT-101) and normal human thyroid cells NTHY-ORI3.1 by RT-qPCR. ***P < 0.001.
Circ_0039411 interference restrains the colony formation ability, migration, invasion, and glycolysis and induces the apoptosis of PTC cells
Loss-of-function experiments were conducted to analyze the functions of circ_0039411 in PTC cells. High knockdown efficiency of si-circ_0039411 in PTC cells was confirmed via RT-qPCR assay (Figure 2(a) and (b)). Colony formation assay revealed that circ_0039411 silencing markedly suppressed the colony formation ability of PTC cells, as evidenced by the decreased number of colonies (Figure 2(c) and (d)). Transwell migration and invasion assays uncovered that circ_0039411 interference reduced the numbers of migrated and invading cells in PTC cells (Figure 2(e)–(h)), demonstrating that circ_0039411 interference suppressed the motility of PTC cells. Flow cytometry showed that silencing circ_0039411 notably induced the apoptosis of PTC cells (Figure 2(i) and (j)). The expression of the proliferation-associated marker (cyclin D1), the motility-associated marker (vimentin), and the pro-apoptotic protein (Bax) was measured to further analyze the functions of circ_0039411 in PTC cells. Circ_0039411 interference reduced the expression of cyclin D1 and vimentin and increased the level of Bax in PTC cells (Figure 2(k) and (l)). As shown in Figure 2(m)–(p), circ_0039411 knockdown suppressed the glucose consumption and the lactate production in PTC cells, suggesting that circ_0039411 knockdown suppressed the glycolytic metabolism of PTC cells. Overall, circ_0039411 knockdown suppressed the malignant behaviors of PTC cells.

Circ_0039411 interference restrains the colony formation ability, migration, invasion, and glycolysis and induces the apoptosis of PTC cells. (a–p) TPC-1 and BHT-101 cells were transfected with si-NC or si-circ_0039411. (a and b) RT-qPCR was conducted to analyze the transfection efficiency of si-circ_0039411. (c and d) Cell colony formation ability was measured by colony formation assay. (e and f) Transwell migration assay was conducted to assess cell migration capacity. (g and h) Cell invasion ability was analyzed by transwell invasion ability. (i and j) Flow cytometry was performed to assess cell apoptosis rate. (k and l) Western blot assay was conducted to measure the expression of the proliferation-associated marker (cyclin D1), the motility-associated marker (vimentin), and the pro-apoptotic protein (Bax) in PTC cells. (m and n) The uptake of glucose was analyzed using Fluorescence-based Glucose Assay Kit. (o and p) The production of lactate was assessed using Fluorescence-based Lactate Assay Kit . ***P < 0.001.
MiR-423-5p is a target of circ_0039411 in PTC cells
MiR-423-5p was notably down-regulated in PTC tissues (n = 51) compared with that in paracancer tissues (n = 51) (Figure 3(a)). We found that miR-423-5p expression was down-regulated in PTC cell lines than that in NTHY-ORI3.1 cell line (Figure 3(b)). CircRNAs can modulate cell phenotypes by serving as miRNA sponges. 17 Through bioinformatic analysis using the StarBase database, we found that miR-423-5p harbored the potential binding sites with circ_0039411 (Figure 3(c)). To analyze whether circ_0039411 interacted with miR-423-5p through the predicted sites, we mutated the putative binding sites in circ_0039411 into “GACGGGGAG” (Figure 3(c)). MiR-423-5p overexpression significantly reduced the luciferase activity of wild type plasmid (circ_0039411 WT) but not that of mutant type plasmid (circ_0039411 MUT) (Figure 3(d) and (e)), suggesting that circ_0039411 interacted with miR-423-5p via the predicted sites. The RNA pull-down assay was conducted to further confirm the binding between circ_0039411 and miR-423-5p. As shown in Figure 3(f), circ_0039411 was significantly enriched in the Bio-miR-423-5p group, suggesting interaction between circ_0039411 and miR-423-5p. Circ_0039411 silencing markedly increased miR-423-5p level in PTC cells (Figure 3(g)). Taken together, circ_0039411 negatively regulated miR-423-5p in PTC cells by directly binding to it.

MiR-423-5p is a target of circ_0039411 in PTC cells. (a) The level of miR-423-5p was determined in 51 pairs of PTC tissues and paracancer tissues by RT-qPCR. (b) MiR-423-5p expression in NTHY-ORI3.1, TPC-1, and BHT-101 cells was measured by RT-qPCR. (c) The binding sites between circ_0039411 and miR-423-5p predicted by StarBase database were shown. In the Dual-Luciferase Reporter Assay, we mutated the predicted miR-423-5p-binding sites in circ_0039411 into “GACGGGGAG,” and the wild-type or mutant type fragment of circ_0039411 was inserted into luciferase vector to obtain circ_0039411 WT or circ_0039411 MUT. (d and e) Dual-Luciferase Reporter Assay was conducted to test whether miR-423-5p was a target of circ_0039411 in PTC cells. (f) RNA-Pull-Down Assay was conducted to confirm the target relation between miR-423-5p and circ_0039411. (g) The expression of miR-423-5p was determined in PTC cells transfected with si-NC or si-circ_0039411 by RT-qPCR. ***P < 0.001.
Circ_0039411 silencing suppresses the malignant behaviors of PTC cells partly by up-regulating miR-423-5p
RT-qPCR assay confirmed the high silencing efficiency of anti-miR-423-5p in PTC cells (Figure 4(a) and (b)). To explore whether circ_0039411 functioned by sponging miR-423-5p, we performed rescue experiments through co-transfecting PTC cells with si-circ_0039411 and anti-miR-423-5p. The addition of anti-miR-423-5p largely rescued the number of colonies and the numbers of migrated and invading cells (Figure 4(c)–(h)), demonstrating that circ_0039411 interference restrained the colony formation ability, migration, and invasion of PTC cells partly by up-regulating miR-423-5p. Circ_0039411 interference-triggered apoptosis in PTC cells was also largely attenuated by the addition of anti-miR-423-5p (Figure 4(i) and (j)). The introduction of anti-miR-423-5p largely recovered the levels of cyclin D1 and vimentin and reduced the expression of Bax (Figure 4(k) and (l)). Circ_0039411 interference reduced the uptake of glucose and the production of lactate, which were largely overturned by the silence of miR-423-5p (Figure 4(m)–(p)). These results demonstrated that circ_0039411 interference restrained the malignant phenotypes of PTC cells partly by up-regulating miR-423-5p.

Circ_0039411 silencing suppresses the malignant behaviors of PTC cells partly by up-regulating miR-423-5p. (a and b) RT-qPCR was applied to analyze the interference efficiency of anti-miR-423-5p in PTC cells. (c–p) TPC-1 and BHT-101 cells were transfected with si-circ_0039411 alone or together with anti-miR-423-5p. (c and d) Colony formation assay was carried out to analyze the colony formation ability of PTC cells. (e and f) The number of migrated cells was analyzed by transwell migration assay. (g and h) Invading cells were counted by transwell invasion assay. (i and j) Cell apoptosis rate was evaluated by flow cytometry. (k and l) The Western blot assay was utilized to measure the protein levels of cyclin D1, vimentin and Bax in PTC cells. (m–p) Cell glycolysis was evaluated using the Fluorescence-based Glucose Assay Kit and the Fluorescence-based Lactate Assay Kit. **P < 0.01, ***P < 0.001.
SOX4 is a target of miR-423-5p in PTC cells
The mRNA and protein expression of SOX4 was up-regulated in PTC tissues compared with that in adjacent normal tissues (Figure 5(a) and (b)). The mRNA and protein levels of SOX4 were markedly elevated in two PTC cell lines than that in NTHY-ORI3.1 cell line (Figure 5(c) and (d)). MiRNAs can regulate gene expression at the post-transcriptional level by binding to the 3′UTR of target mRNAs. 20 The putative binding sites between miR-423-5p and SOX4 3′UTR predicted by the StarBase database are shown in Figure 5(e). Luciferase activity of wild type plasmid (SOX4 3′UTR-WT) was notably reduced by the transfection of miR-423-5p rather than miR-NC, and luciferase activity of mutant type plasmid (SOX4 3′UTR MUT) was unchanged by the transfection of miR-NC or miR-423-5p (Figure 5(f) and (g)), suggesting that miR-423-5p bound to the 3′UTR of SOX4 in PTC cells. High overexpression efficiency of miR-423-5p was validated by RT-qPCR in PTC cells (Figure 5(h)). MiR-423-5p overexpression markedly decreased the protein level of SOX4 in PTC cells (Figure 5(i)). Considering the interaction and negative regulatory relationship between miR-423-5p and circ_0039411 or SOX4, we then analyzed whether circ_0039411 indirectly regulated SOX4 expression by targeting miR-423-5p in PTC cells. Circ_0039411 silencing reduced the protein expression of SOX4, and the addition of anti-miR-423-5p largely rescued the SOX4 protein level in PTC cells (Figure 5(j) and (k)), suggesting that circ_0039411 interference down-regulated SOX4 expression partly by up-regulating miR-423-5p in PTC cells. These results suggested that SOX4 was a target of miR-423-5p, and that SOX4 was regulated by the circ_0039411/miR-423-5p axis in PTC cells.

SOX4 is a target of miR-423-5p in PTC cells. (a and b) RT-qPCR and Western blot assay were conducted to determine the mRNA and protein levels of SOX4 in PTC tissues (n = 51) and in paracancer normal tissues (n = 51). (c and d) The mRNA and protein expression of SOX4 was examined in NTHY-ORI3.1, TPC-1 and BHT-101 cells by RT-qPCR and Western blot assay. (e) The binding sequence between miR-423-5p and SOX4 predicted by StarBase database was shown. The fragment of SOX4 3′UTR, containing the wild-type (“CCCCUC”) or mutant type (“GGGGAG”) binding sites with miR-423-5p, was inserted into the luciferase vector to obtain SOX4 3′UTR WT or SOX4 3′UTR MUT. (f and g) The target relationship between miR-423-5p and SOX4 was analyzed by Dual-Luciferase Reporter Assay. (h) The overexpression efficiency of miR-423-5p was analyzed by RT-qPCR in PTC cells. (i) Western blot assay was conducted to measure the protein level of SOX4 in miR-423-5p-overexpressed PTC cells. (j and k) The Western blot assay was conducted to measure the protein expression of SOX4 in PTC cells in the following four groups: si-NC, si-circ_0039411, si-circ_0039411 + anti-NC, si-circ_0039411 + anti-miR-423-5p. ***P < 0.001.
SOX4 overexpression largely rescues the malignant behaviors in circ_0039411-silenced PTC cells
Western blot assay verified the high overexpression efficiency of SOX4 plasmid in PTC cells (Supplementary Figure 1(a) and (b)). To investigate whether circ_0039411 knockdown restrained the malignant behaviors of PTC cells by regulating SOX4, rescue experiments were conducted through co-transfecting PTC cells with si-circ_0039411 and SOX4 plasmid. SOX4 overexpression largely rescued the colony formation ability in PTC cells (Supplementary Figure 1(c) and Supplementary 1(d)). The numbers of migrated and invading PTC cells were reduced by circ_0039411 interference, and were largely recovered by the addition of SOX4 plasmid (Supplementary Figure 1(e)–(h)). Circ_0039411 interference-induced apoptosis in PTC cells was also largely diminished by the overexpression of SOX4 (Supplementary Figure 1(i) and Supplementary 1(j)). SOX4 overexpression rescued the expression of cyclin D1 and vimentin and decreased the level of Bax in circ_0039411-silenced PTC cells (Supplementary Figure 1(k) and Supplementary 1(l)). SOX4 overexpression also largely rescued the glycolytic rate in circ_0039411-silenced PTC cells (Supplementary Figure 1(m)–(p)), as evidenced by the increased glucose consumption and lactate production in si-circ_0039411 and the SOX4 co-transfected group. Taken together, circ_0039411 interference restrained the malignant behaviors of PTC cells partly by down-regulating SOX4 expression.
Circ_0039411 interference restrains the tumor growth in vivo
Given the oncogenic role of circ_0039411 in PTC cells in vitro, we subsequently analyzed whether circ_0039411 exerted a similar role in vivo. TPC-1 cells stably expressing sh-NC or sh-circ_0039411 were subcutaneously injected in the mice to establish the xenograft tumor model. Circ_0039411 interference significantly restrained the tumor growth in vivo (Supplementary Figure 2(a) and (b)). We analyzed the level of proliferation-marker Ki-67 in tumor tissues by IHC assay. The results showed that tumors formed from circ_0039411-silenced TPC-1 cells displayed a lower intensity of Ki-67 staining (Supplementary Figure 2(c)). The expression of circ_0039411 and SOX4 protein was decreased in sh-circ_0039411 group (Supplementary Figure 2(d) and Supplementary 2(e)), and circ_0039411 silencing up-regulated the miR-423-5p level in tumor tissues in vivo (Supplementary Figure 2(d)). Overall, circ_0039411 silencing inhibited the tumor growth in vivo.
Discussion
Although a large number of circRNAs have been identified to be dysregulated in human malignancies, 27 only a few circRNAs have been studied. CircRNAs play pivotal roles in the initiation and development of cancers by serving as oncogenes or tumor suppressors. 28 In this study, the function and the miRNA sponge role of circ_0039411 in PTC were uncovered. We demonstrated that circ_0039411 facilitated the malignant behaviors of PTC cells by sponging miR-423-5p to induce SOX4 expression. The binding between miR-423-5p and circ_0039411 or SOX4 was found for the first time in this study, and the role of SOX4 in PTC was also reported for the first time.
Circ_0039411 has shown an oncogenic role in lung adenocarcinoma, 29 hepatocellular carcinoma, 30 and PTC. 16 Lv et al. demonstrated that circ_0039411 facilitated the proliferation and migration capacities of lung adenocarcinoma cells. 29 Yang et al. found that circ_0039411 expression was elevated in PTC tissues and cells, and it contributed to PTC development by targeting both the miR-1179/ABCA9 signaling and the miR-1205/MTA1 signaling. 16 We found that circ_0039411 expression was significantly up-regulated in PTC tissues and cell lines, and it promoted the colony formation ability, motility, and glycolysis and inhibited the apoptosis of PTC cells. The oncogenic role of circ_0039411 was consistent with a former article. 16
Increasing studies have shown that circRNAs can function as miRNA sponges, which results in the up-regulation of downstream genes.31,32 For instance, circ_0004458 facilitated PTC development through elevating RAC1 expression via absorbing miR-885-5p. 33 Circ-PSD3 promoted the malignant behaviors of PTC cells through up-regulating HEMGN via sponging miR-637. 34 MiR-423-5p was confirmed as a target of circ_0039411 in PTC cells. MiR-423-5p exhibited a tumor suppressor role in various malignancies, including PTC.19,35‐37 For example, miR-423-5p suppressed colon cancer progression through inducing cell apoptosis. 38 MiR-423-5p restrained the malignant properties of ovarian cancer cells. 36 Yang et al. found that NR2F1-AS1 contributed to the progression of PTC through reducing the miR-423-3p level and up-regulating SOX12, 19 suggesting the tumor suppressor role of miR-423-5p in PTC cells. We found that circ_0039411 negatively regulated the miR-423-5p level in PTC cells. To test if circ_0039411 silencing-mediated effects were dependent on the up-regulation of miR-423-5p, we performed rescue experiments. The results revealed that circ_0039411 silencing restrained the malignant phenotypes of PTC cells partly by up-regulating miR-423-5p.
MiRNAs suppress the translation or induce the degradation of target mRNAs by binding to their 3′UTR. 20 We predicted the downstream targets of miR-423-5p using a bioinformatic database. Through the Dual-Luciferase Reporter Assay, SOX4 was verified as a target of miR-423-5p in PTC cells. SOX4 is a member of the SOX transcription factor family, and exhibits a promoting or inhibitory role in gene transcription. 39 Convincing evidence has confirmed the oncogenic role of SOX4 in several cancers. For instance, miR-129-2 inhibited the development of esophageal carcinoma through reducing the SOX4 level. 40 Zhang et al. demonstrated that SOX4 facilitated the epithelial-mesenchymal transition of breast cancer cells. 41 SOX4 expression was elevated in PTC tissue samples and cell lines. MiR-423-5p overexpression reduced SOX4 expression in PTC cells. Through rescue experiments, we found that circ_0039411 interference restrained the malignant properties of PTC cells partly by reducing SOX4 expression. Circ_0039411 positively regulated SOX4 expression partly by acting as an miR-423-5p sponge in PTC cells.
Given the pro-tumor role of circ_0039411 in vitro, we assessed whether circ_0039411 exhibited a similar function in the tumor growth in vivo. Circ_0039411 knockdown markedly restrained the tumor growth, suggesting that circ_0039411 promoted PTC progression in vivo.
In conclusion, our data suggested that circ_0039411 was notably up-regulated in PTC tissues and cell lines. Circ_0039411 interference suppressed the colony-formation ability, migration, invasion, and glycolysis and triggered the apoptosis of PTC cells. Mechanistic analysis revealed that circ_0039411 functioned partly by sponging miR-423-5p and up-regulating SOX4, which provided novel potential targets for PTC therapy.
Supplemental Material
sj-doc-1-jbm-10.1177_17246008211043128 - Supplemental material for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling
Supplemental material, sj-doc-1-jbm-10.1177_17246008211043128 for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling by Xiaohui Wen, Jingyan Du and Xun Wang in The International Journal of Biological Markers
Supplemental Material
sj-tif-2-jbm-10.1177_17246008211043128 - Supplemental material for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling
Supplemental material, sj-tif-2-jbm-10.1177_17246008211043128 for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling by Xiaohui Wen, Jingyan Du and Xun Wang in The International Journal of Biological Markers
Supplemental Material
sj-tif-3-jbm-10.1177_17246008211043128 - Supplemental material for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling
Supplemental material, sj-tif-3-jbm-10.1177_17246008211043128 for Circ_0039411 promotes papillary thyroid carcinoma development through mediating the miR-423-5p/SOX4 signaling by Xiaohui Wen, Jingyan Du and Xun Wang in The International Journal of Biological Markers
Footnotes
Ethics approval and consent participate
Written informed consent was obtained from patients with approval by the Institutional Review Board in Beijing Chaoyang Hospital, Capital Medical University.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.
Availability of data and materials
Please contact the correspondence author for the data request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.