
Abstract
Atherosclerosis is a chronic inflammatory disease in the vessel wall. As a representative inflammatory cytokine, C-reactive protein (CRP) participates in the formation and development of atherosclerosis. It is demonstrated that aldosterone induces CRP generation in vascular smooth muscle cells (VSMCs). This study explored the inhibitory effect of eplerenone on aldosterone-induced CRP expression in VSMCs and mechanism. In the in vitro experiments, rat VSMCs were cultured and aldosterone (10 nM) was used as a stimulant for CRP generation. VSMCs were pretreated with eplerenone for 1 h prior to the stimulation. Messenger RNA (mRNA) and protein expression were identified by reverse transcription quantitative polymerase chain reaction (RT-qPCR) and western blot, respectively. The production of reactive oxygen species (ROS) was observed by a fluorescence microscope. In the in vivo experiment, a model of hyperaldosteronism was established by the subcutaneous administration of aldosterone to rats with the osmotic minipumps for 4 weeks. Serum aldosterone and CRP levels were determined with a radioimmunoassay and ELISA (enzyme-linked immunosorbent assay), respectively. The results showed that eplerenone inhibited aldosterone-induced mRNA and protein expression of CRP in VSMCs in vitro and in vivo, and decreased the circulating CRP level of hyperaldosteronism rats. Meanwhile, eplerenone reduced aldosterone-stimulated ROS generation and aldosterone-activated ERK1/2 phosphorylation in VSMCs. In summary, eplerenone inhibits aldosterone-induced CRP generation in VSMCs by regulating the MR-ROS-ERK1/2 signal pathway. These results provide new evidence for the potential anti-inflammatory effect of eplerenone.
Introduction
Atherosclerosis is a common cardiovascular disease and the main pathological basis of ischemic cardiocerebrovascular diseases. The traditional viewpoints think that atherosclerosis is a progressive narrowing of the artery lumen caused by hyperlipidemia. However, increasing epidemiological evidence and experimental data support that atherosclerosis is a chronic inflammatory disease in the vessel walls,1,2 and inflammation takes part in the pathological process of atherosclerosis.
As a classic inflammatory cytokine, C-reactive protein (CRP) is not only a predictor but also an important risk factor for the cardiovascular diseases. 3 CRP promotes the formation and development of atherosclerosis by inducing the vascular endothelial dysfunction, promoting the monocyte/macrophage adhesion to the vascular endothelium, stimulating the proliferation of VSMCs, activating the complement system, and facilitating thrombosis formation. 4
Aldosterone is a final product of the renin–angiotensin–aldosterone system. With the development of diagnostic technology, the morbidity of hyperaldosteronism increases recently. The abnormally elevated aldosterone level in plasma is considered to be a risk factor of many cardiovascular diseases.5,6 Aldosterone causes the vascular contraction and the endothelial dysfunction, and stimulates the expressions of inflammatory cytokines. 6 Our previous study demonstrated that aldosterone stimulates CRP generation in rat VSMCs via the mineralocorticoid receptor (MR), reactive oxygen species (ROS), and extracellular signal-regulated kinase (ERK1/2) signal pathway. 7
Eplerenone is a novel selective MR antagonist and used for the treatment of hypertension and heart failure in clinic practice. Multiple studies suggest that eplerenone inhibits the expressions of the inflammatory cytokines, decreases the oxidative stress level, and improves myocardial hypertrophy and injury.8–10 However, it is unknown whether eplerenone produces an anti-inflammation by inhibiting CRP generation in VSMCs. Therefore, this study examined the inhibitory effect of eplerenone on aldosterone-induced CRP generation in VSMCs and the related molecular mechanisms.
Materials and methods
Reagents
Fetal bovine serum (FBS) and Dulbecco’s High Glucose Modified Eagle’s Medium (DMEM) were ordered from HyClone (UT, USA). Eplerenone and aldosterone were from Sigma–Aldrich (St. Louis, MO, USA). Rabbit polyclonal CRP antibody was provided by Abcam (Cambridge, MA, USA). Mouse monoclonal glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody was purchased from CoWin Biotech (Beijing, China). Mouse monoclonal ERK1/2, phospho-ERK1/2, α-actin antibodies, and 2′, 7′-dichlorodihydrofluororescein diacetate (H2DCF-DA) were obtained from Beyotime (Jiangsu, China). ELISA (enzyme-linked immunosorbent assay) kit for the detection of CRP was produced by Westtang (Shanghai, China).
Culture of rat VSMCs
The primary VSMCs from the thoracic aorta of male Sprague Dawley (SD) rats were cultured in DMEM containing 10% FBS, 100 U/mL penicillin, and 100 U/mL streptomycin at 37°C in a humidified atmosphere of 5% CO2 and 95% air until the cells exhibited the typical “hill and valley” growth pattern. 11 Identity of VSMCs was confirmed by the morphological examination and the immunocytochemical staining for α-actin. The confluent cells at passages 3–8 were used for the study. Before the experiments, the cells were incubated in the serum-free medium for an additional 24 h.
Reverse transcription quantitative polymerase chain reaction
Total RNA was purified from VSMCs using RNAfast200 purification kit. Complementary DNA (cDNA) was synthesized from total RNA by Revert Aid™ First Strand cDNA synthesis kit according to the procedures provided by manufacturer. The specific cDNA was amplified using primer pairs specific for rat CRP (sense primer: 5′-CATCTGTGCCACCTGGGAGTC-3′; anti-sense primer: 5′-AAGCCACCGCCATACGAGTC-3′). GAPDH was amplified as an internal control for normalization (sense primer: 5′-GCAAGTTCAACGGCACAGTCAAG-3′; anti-sense primer: 5′-ACATACTCAGCACCAGCATCACC-3′). PCR products were separated by agarose gel electrophoresis. Expression of mRNA was quantified as relative to internal-control GAPDH.
Western blot
After the treatments, VSMCs were lysed with the lysis buffer containing the protease inhibitors, cocktail, and the phosphatase inhibitors. The protein level was detected by the BCA (bicinchoninic acid assay) protein assay kit. Protein extract (35 μg) was loaded, separated by 10% SDS-PAGE and blotted onto nitrocellulose membrane. Then, the membranes were incubated with anti-CRP (1:150 dilution), anti-GAPDH (1:500 dilution), anti-ERK1/2 (1:500 dilution), and anti-phospho-ERK1/2 (1:500 dilution) antibodies overnight at 4°C. After washed, the membranes were incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit or anti-mouse second antibody (1:6000 dilution) followed by the enhanced chemiluminescence. Optical density of the bands was scanned and quantified with Gel Doc 2000 (Bio-Rad). GAPDH was used as loading control.
Measurement of ROS in VSMCs
The intracellular ROS were measured using the H2DCF-DA fluorescent labeling method. 12 VSMCs were pretreated with eplerenone at 0.1, 1, and 10 μM for 1 h prior to the exposure to 10 nM aldosterone for 2 h. Then, the cells were loaded with 10 μM H2DCF-DA for 1 h. Fluorescence images were acquired at the excitation wavelength of 488 nm and the emission wavelength of 525 nm with fluorescence microscope (Olympus BX51, Japan). Fluorescence intensity was determined and analyzed from the fluorescence images with the Image-pro plus software. The relative fluorescence intensity was taken as the average of values from three independent experiments.
Animal model
To investigate whether eplerenone reduced aldosterone-induced CRP generation in vascular smooth muscle tissue, a model of hyperaldosteronism was established by the subchronic administration of aldosterone to rats. 13 Male SD rats (n = 24, weight = 180–220 g) were randomly assigned to control, hyperaldosteronism model, and two eplerenone-treated groups. Rats in the model and eplerenone-treated groups were subcutaneously delivered with 0.75 μg/h aldosterone for a period of 4 weeks via the implanted osmotic minipumps. Rats in control group received vehicle (PEG 400) in an identical fashion to the model group. Meanwhile, eplerenone at the doses of 50 and 100 mg/kg/day was intragastrically administered to rats in eplerenone-treated groups for 4 weeks, and physiological saline was given to rats in control and the model groups instead. Systolic blood pressure (SBP) was measured with a non-invasive tail cuff method in the beginning and the end of the experiment. In the end of the experiment, blood was collected for the detection of aldosterone and CRP concentrations in serum by a radioimmunoassay (RADIM Diagnostics, Italy) and ELISA, respectively. Then, the thoracic aorta of rats was immediately removed for the determination of CRP mRNA expression by reverse transcription quantitative polymerase chain reaction (RT-qPCR) and protein expressions of CRP, ERK1/2, and p-ERK1/2 with western blot. All rats were from the Experimental Animal Center of Xi’an Jiaotong University Health Center. Animal care and procedures were performed in accordance with the Laboratory Animal Care Guidelines approved by Medical Ethics Committee of Xi’an Jiaotong University.
Statistical analysis
All data were presented as mean ± standard error of mean (SEM). Statistical significance between groups was determined by one-way analysis of variance (ANOVA) with the SPSS 13.0 program (SPSS Inc., Chicago, IL, USA). A value of P < 0.05 was considered significant.
Results
Eplerenone inhibits aldosterone-induced CRP expression in VSMCs
Figure 1 showed that mRNA and protein expression of CRP was significantly increased after VSMCs were stimulated with 10 nM aldosterone for 24 h (P < 0.01 vs control). The pretreatment of the cells with eplerenone at 0.1, 1, and 10 μM prior to aldosterone stimulation significantly inhibited aldosterone-induced mRNA and protein expression of CRP in VSMCs in a concentration-dependent way (P < 0.05 or P < 0.01 vs aldosterone alone). In addition, eplerenone alone did not affect the basal CRP expression in VSMCs.

Effect of eplerenone on the ALD-induced CRP expression in VSMCs. VSMCs were preincubated with the different concentrations of eplerenone (EPL) for 1 h before the stimulation for 24 h with 10 nM aldosterone (ALD). (a) mRNA and (b) protein expression of CRP was identified by RT-qPCR and western blot, respectively. EPL alone (10 μM) was used as drug control. Results from three independent experiments were expressed as means ± SEM (n = 3).
Eplerenone reduces aldosterone-stimulated ROS generation in VSMCs
To probe whether the inhibition by eplerenone of aldosterone-induced CRP expression is related to interfering with ROS generation, the intracellular ROS generation was detected using ROS fluorescent probe H2DCF-DA. As seen from Figure 2, aldosterone at 10 nM increased ROS generation in VSMCs (P < 0.001 vs control), and the pretreatment of the cells with eplerenone at 0.1, 1, and 10 μM reduced aldosterone-stimulated ROS generation in VSMCs in a concentration-dependent manner (P < 0.001 vs aldosterone alone).

Effect of eplerenone on the ALD-stimulated ROS generation in VSMCs. VSMCs were pretreated with the different concentrations of eplerenone (EPL) for 1 h prior to the stimulation for 2 h with 10 nM aldosterone (ALD), and then incubated with H2DCF-DA (10 μM) for 1 h in the serum-free DMEM. Finally, the cells were observed and the pictures were captured under fluorescence microscope. The fluorescent intensity was measured by a fluorescence microscope. (a) Representative fluorescence images: (a) control, (b) ALD alone, (c) ALD + 0.1 μM EPL, (d) ALD + 1 μM EPL, and (e) ALD + 10 μM EPL. (b) Relative fluorescence intensity quantified from the fluorescence images. Results from three independent experiments were expressed as mean ± SEM (n = 5).
Eplerenone suppresses aldosterone-activated ERK1/2 phosphorylation in VSMCs
We previously reported that aldosterone activates ERK1/2, which mediates aldosterone-induced CRP expression in VSMCs. 7 In the present research, the effect of eplerenone on aldosterone-activated ERK1/2 phosphorylation was assayed with western blot. The result in Figure 3 illustrated that the phosphorylated ERK1/2 expression increased after the stimulation of the cells for 30 min with aldosterone (P < 0.001 vs control), while the pretreatment of the cells with eplerenone for 1 h before the exposure to aldosterone inhibited aldosterone-activated ERK1/2 phosphorylation (P < 0.01 or P < 0.001 vs aldosterone alone).
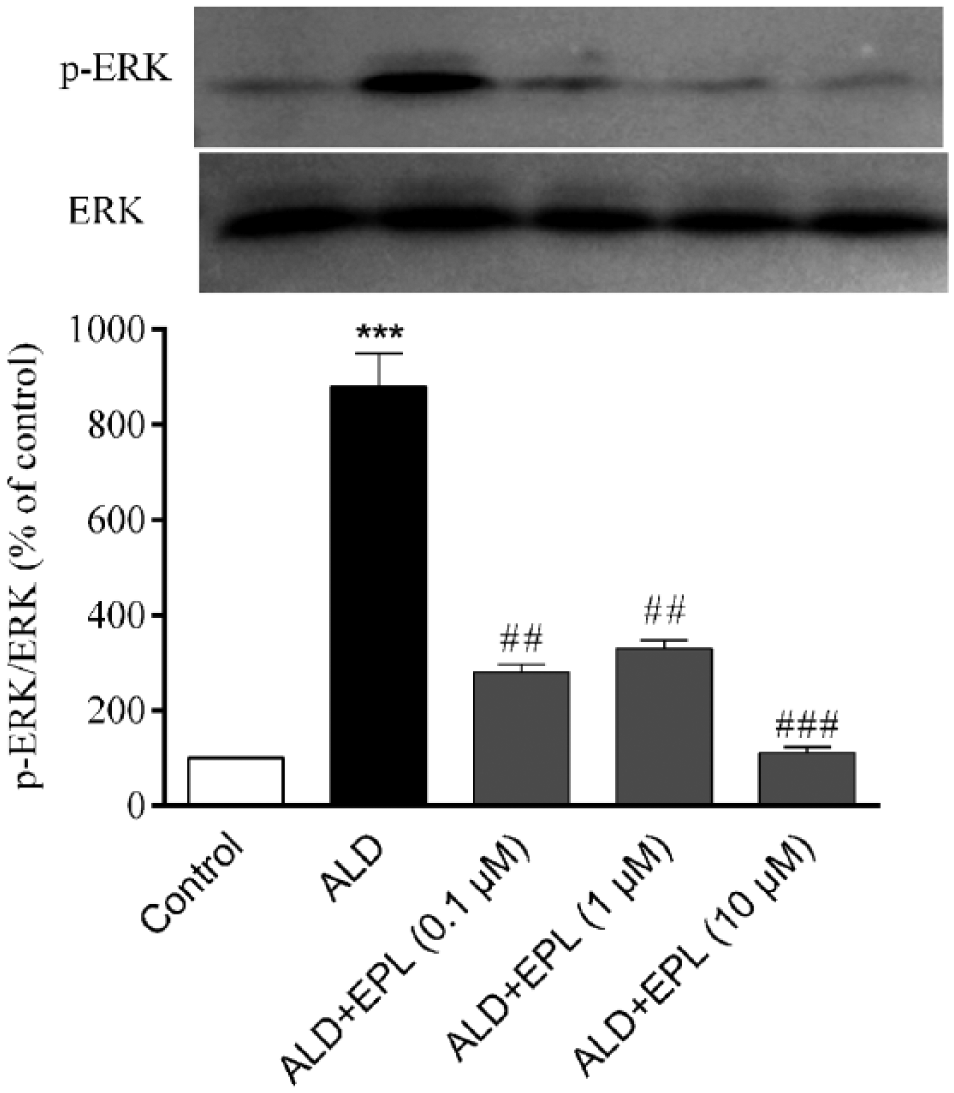
Effect of eplerenone on the ALD-activated ERK1/2 phosphorylation in VSMCs. After the pretreatment with the different concentrations of eplerenone (EPL) for 1 h, VSMCs were stimulated with 10 nM aldosterone (ALD) for 30 min. Then, ERK1/2 phosphorylation was detected by western blot. Results from three independent experiments were expressed as mean ± SEM (n = 3).
Eplerenone decreases serum CRP level in hyperaldosteronism rats
To observe the effect of eplerenone on aldosterone-stimulated CRP production in vivo, a hyperaldosteronism model of rats was established. Infusion of aldosterone to rats for 4 weeks did not affect the body weight and SBP (data not shown) and however, elevated aldosterone and CRP levels in serum (P < 0.001 and P < 0.05 vs control). In comparison with the model rats, rats receiving eplerenone exhibited the lower serum CRP level (P < 0.05 or P < 0.01 vs model), but not aldosterone (Figure 4).

Effects of eplerenone on ALD and CRP levels in serum of hyperaldosteronism rats. Rats were treated by the subcutaneous administration of aldosterone (ALD, 0.75 μg/h) with an osmotic minipump for 4 weeks, and simultaneously received eplerenone (EPL). In the end of the experiment, blood was collected for the detection of serum ALD level (a) by a radioimmunoassay and CRP concentration (b) with ELISA. Data were expressed as mean ± SEM (n = 5).
Eplerenone down-regulates CRP expression in the thoracic aorta of hyperaldosteronism rats
The sustained infusion of aldosterone to rats increased both mRNA and protein expression of CRP in the thoracic aorta (P < 0.001 vs control). However, the simultaneous treatment of rats with eplerenone down-regulated CRP expression in the aortic walls of hyperaldosteronism rats in mRNA and protein levels (Figure 5; P < 0.001 vs model).

Effect of eplerenone on CRP expression in the thoracic aorta of hyperaldosteronism rats. Rats were treated by the subcutaneous administration of aldosterone (ALD, 0.75 μg/h) with an osmotic minipump for 4 weeks, and simultaneously received eplerenone (EPL). In the end of the experiment, the thoracic aorta was removed for the examination of (a) mRNA and (b) protein expression of CRP by RT-qPCR and western blot, respectively. Data were expressed as mean ± SEM (n = 5).
Eplerenone diminishes ERK1/2 phosphorylation in the thoracic aorta of hyperaldosteronism rats
The in vivo experiment also showed that ERK1/2 phosphorylation increased in the thoracic aorta of hyperaldosteronism rats (P < 0.001 vs control). ERK1/2 phosphorylation reduced in the thoracic aorta of eplerenone-treated rats as compared to the model rats, (Figure 6; P < 0.01 vs model).

Effect of eplerenone on ERK1/2 phosphorylation in the thoracic aorta of hyperaldosteronism rats. Rats were treated by the subcutaneous administration of aldosterone (ALD, 0.75 μg/h) with an osmotic minipump for 4 weeks, and simultaneously received eplerenone (EPL). In the end of the experiment, the thoracic aorta was removed for the determination of ERK1/2 phosphorylation by western blot. Data were expressed as mean ± SEM (n = 5).
Discussion
Aldosterone is an important mineralocorticoid hormone regulating sodium and potassium transports in the epithelial cells of renal tubules. Excess aldosterone results in the abnormal water homeostasis and increases the risk of cardiovascular events independent of hypertension.5,6 Aldosterone exerts its actions by activating MR in cytoplasm. As an MR antagonist, eplerenone is able to attenuate aldosterone-induced inflammation and oxidative stress. Rocha et al. 14 report that eplerenone down-regulates aldosterone-induced expressions of osteopontin, COX-2, MCP-1, and TGF-β1 in the rat heart. In the apolipoprotein E-deficient mice fed with high-cholesterol diet, eplerenone reduces the expressions of NAD(P)H oxidase, TNF-α, and MCP-1 to prevent the formation of atherosclerosis. 8 Our previous study demonstrated that aldosterone induces CRP expression in VSMCs in vitro and in vivo. 7 In this experiment, we observed the effect of eplerenone on aldosterone-induced CRP generation in VSMCs. The results displayed that eplerenone inhibited aldosterone-induced mRNA and protein expression of CRP in VSMCs in vitro and in vivo, and lowered serum CRP level in hyperaldosteronism rats without affecting blood pressure. These results confirm that eplerenone has the ability to alleviate the vascular inflammation by suppressing CRP generation in VSMCs, which provides a protection on the cardiovascular system independent of its hypotensive effect. Our result also showed that eplerenone did not alter aldosterone concentration in the serum of hyperaldosteronism rats, which implies that eplerenone does not affect aldosterone metabolism and eplerenone-inhibited CRP expression is not accomplished though regulating aldosterone metabolism. Comparison of the results in Figures 4 and 5 found that two doses of eplerenone both lowered serum CRP level; however, only high dose of eplerenone inhibited CRP expression in the aortic walls of hyperaldosteronism rats. Since plasma CRP is from multiple tissues, this suggests that eplerenone possibly inhibits CRP generation in other tissues, which merits further investigation.
ROS are both the intracellular second messenger and the direct participant of oxidative stress. 15 The recent researches have shown that ROS play the important roles in the aldosterone-elicited cardiovascular pathological changes. 16 In the aldosterone-induced hypertensive rats, NAD(P)H oxidase activity and ROS generation increased.17,18 NAD(P)H oxidase inhibitor apocynin decreases blood pressure and relieves cardiac hypertrophy in aldosterone-infused rats. 19 Rajagopalan et al. 20 report that eplerenone improves the endothelial function and reduces the superoxide generation in the diet-induced atherosclerosis rabbits. Callera et al. 21 find that eplerenone inhibits the aldosterone-induced increase of NAD(P)H oxidase activity. Our previous experiment showed that the intracellular ROS are essential for the aldosterone-induced CRP expression in VSMCs since antioxidant N-acetylcysteine abolishes the aldosterone-induced CRP protein expression. 7 The present result indicated that eplerenone diminished the aldosterone-stimulated ROS generation in VSMCs. It is known that aldosterone-stimulated ROS generation via activating MR 7 and eplerenone is a selective MR antagonism. This suggests that eplerenone reduces the aldosterone-stimulated ROS generation through blocking MR.
Mitogen-activated protein kinase (MAPK) signaling mediates the generation of inflammatory cytokines. Eplerenone down-regulates the expressions of inflammatory molecules like ICAM-1, VCAM-1, and eNOS in the renal cortex of Dahl salt-sensitive hypertensive rats by inhibiting MAPK signal pathway. 22 Nagai et al. 23 document that eplerenone attenuates the aldosterone-induced ERK1/2 phosphorylation in rat renal fibroblasts. The results from our previous experiment also implicated that aldosterone activates ERK1/2 phosphorylation via MR-ROS pathway. 7 This study displayed that eplerenone inhibited the aldosterone-activated ERK1/2 phosphorylation in VSMCs in vitro and in vivo. In combination of the results with the previous reports, it is inferred that eplerenone inhibits the aldosterone-activated ERK1/2 phosphorylation by interfering with MR-ROS signal pathway in VSMCs.
In summary, this study demonstrates that eplerenone inhibits the aldosterone-induced CRP generation in VSMCs by regulating the MR-ROS-ERK1/2 signal pathway. These results provide new experimental evidence for the potential anti-inflammatory effect of eplerenone.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was financially supported in part by the Doctoral Fund of Ministry of Education of China (no. 20100201110053).