
Abstract
Biological evidence suggests that fish use muscles to stiffen their bodies and improve their swimming performance. Inspired by this phenomenon, we propose a planar serial–parallel mechanism with variable stiffness to mimic a swimming fish. Based on Lighthill’s elongated-body theory, we present a general method to design the body stiffness, which is related to morphological parameters and the swimming frequency. The results show that the stiffness profile is directly proportional to the square of the driving frequency. Furthermore, a SimMechanics model of a robotic fish is innovatively built. Numerical results show that the fish with the designed stiffness has the maximum speed when the driving frequency is close to the resonance frequency of fish body, and that the maximum speed is linearly proportional to the resonance frequency. The range of the Strouhal number given by simulations is also consistent with the range 0.25 < St < 0.35 required by the optimal efficiency. All these results agree well with biological observations, indicating that the swimming performance of fish is significantly affected by the body stiffness and the driving frequency.
Introduction
Fish have attracted the interests of researchers because they have far more superior swimming performance to man-made underwater vehicles. 1 Different types of fish have evolved to achieve different swimming performance, such as cruising long distances at a significant speed, maneuvering in tight spaces, and accelerating swiftly. 2 Various bio-inspired swimming robots have been built and applied to seabed exploration, underwater transportation, search and rescue, and so on. 3 These artificial fish-like robots provide innovative solutions to underwater propulsion and maneuvering. More detailed discussions on swimming fish can be found in Colgate and Kevin 2 and Lauder. 4
The synthetic studies of swimming fish are not only of engineering value but also of biological value, because the necessary kinematic and hydrodynamic data are acquired directly from biological experiments.5–7 Recently, many works have focused on the effects of body stiffness on the swimming ability of fish.7,8 For example, Tytell et al. 7 analyzed a two-dimensional model of eel fish and suggested that given the same actuation of the muscles, fish with different body stiffness would have different motions. Lauder et al. 9 demonstrated that flexibility has important effects on vectoring forces and generating thrust in the sunfish motion. Park et al. 10 clarified that the optimal compliance of bionic system could improve the thrust and swimming efficiency. Ahlborn et al. 11 also found that in the fast-start swimming of fish, a slightly flexible tail generated the highest impulse related to thrust generation.
The biological studies suggested that fish can adjust their body stiffness to improve their swimming performance, and some fascinating applications also support this idea. For instance, McHenry et al. 12 found that the live pumpkinseed must increase body stiffness to double the level of passive stiffness in order to achieve their swimming speeds using a sunfish model constructed from vinyl material. One prevalent hypothesis is that fish maximize swimming efficiency or thrust by tuning their natural frequency to the tail-beat frequency.13,14 From a biomechanical perspective, swimming fish use their muscles, collagenous fibers, tendons, and skin to modulate their natural frequency.15,16 In the studies of a robotic dolphin with an actuated joint at the peduncle and a passive compliant joint at the caudal fin, Nakashima and Ono 17 found the correlations of speed, tail-beat frequency, and reduced frequency with stiffness of the compliant joint, providing a possible basis for performance optimization. This is qualitatively consistent with biological data summarized by Bandyopadhyay, 18 who found that the dolphin tail-beat frequency approaches the natural frequency. Furthermore, Harper et al. 19 showed that an appropriate adjustment of the stiffness of the oscillating foil could reduce the energy consumption in heave motions. Similarly, Triantafyllou et al. 20 and Prempraneerach et al. 21 investigated the effects of the passive stiffness of a flapping foil on its thrust-generation capability.
The understanding of these principles of fish swimming has in turn motivated efforts to replicate such superb performance in robotic fish. Traditionally, the motions of the robotic fish have been implemented using complex series mechanisms that employ several discrete, stiff components.22,23 As a result, several actuators are required, along with sophisticated control schemes. Alternatively, Valdivia y Alvarado and colleagues24,25 and Marchese et al. 26 built several soft robotic fish with viscoelastic body. These biomimetic devices have a better swimming performance when the driving frequency is close to the resonance frequency.24,25 However, due to the properties of viscoelastic material (e.g. silicon and rubber), their body stiffness cannot be adjusted as needed, and they swam quickly and efficiently only at a certain driving frequency around resonance frequency. The biological structure of a fish, together with the muscles, skin, muscle tendons, and skeletal system, is a complicated serial–parallel mechanism.27,28 Nevertheless, relatively little work has been done to identify and apply the specific features of the serial–parallel mechanism that benefits the design of robotic fish. Therefore, this article aims to construct a robotic model with a planar serial–parallel mechanism that allows a detailed control over the variable body stiffness, which will contribute to the understanding of fish body stiffness.
While the body stiffness has a significant effect on the swimming performance, 29 there remains huge challenges in building a dynamic model of swimming fish to predict their kinematics. Long et al. 30 built a dynamic model to study the force transmission via axial tendons in undulating fish and predicated four axial tendons functions to produce the most realistic fish motion. In this model, the intervertebral joint acted as a linear spring with a constant Young’s modulus, and the lateral motion, especially the tail’s motion, cannot be fully captured after three beating cycles. Different from the model in Long et al., 30 we design the arbitrary body stiffness of undulating fish based on their realistic fish kinematic and the theory of large-amplitude elongate body. 31 The reactive and resist forces of surrounding fluid are also fully captured in this study. Therefore, the results of our model emerge from the interactions of actuated pattern, passive body properties and the fluid environment. Unlike traditional fish robots,22,23 our model using serial–parallel mechanism is strongly under-actuated with only one single actuator, and the motions of rest parts follow the first actuated serial–parallel unit passively. In summary, the main contribution of this article is to propose an advanced fish model with serial–parallel mechanisms and provide an innovative method to design the body stiffness to achieve fish realistic motion. This study can also be used to guide the design of fish-like robots to obtain an optimal swimming performance. The rest of this article is organized as follows. In section “Design of robotic fish with the serial–parallel mechanism,” the design of robotic fish with the serial–parallel mechanism is proposed, and a numerical example is introduced in section “A numerical example.” The process to build the SimMechanics model of robotic fish and its analysis results are presented in sections “The SimMechanics model of robotic fish” and “Analysis of the robotic fish,” respectively. Finally, the conclusions are summarized in section “Conclusion.”
Design of robotic fish with the serial–parallel mechanism
Structural parameters
In fish swimming, the muscle fibers drive the interconnected vertebrae to rotate a small relative angle and produce thrust by propelling the surrounding water through body waves. 32 In this regard, traditional robotic fish mimic the oscillating motion of real fish using a multi-joint serial mechanism. Meanwhile, the viscoelastic forces or torsional moments are generated by axial tendons and muscular tissues attached to skeleton and ribs through a parallel mechanism.28,32 Inspired by these facts, we propose a planar serial–parallel mechanism to mimic the biological structure of fish.
Figure 1 shows three connected units of serial–parallel mechanical structure in the fish model. Each unit

Serial–parallel mechanism derived from the biological structure.
In nature, the body deformation of swimming fish is determined by its mechanical properties and the fluid environment, and the morphometric measurements help us identify the geometric and physical parameters of the fish model with a serial–parallel mechanism. Figure 2 is the top view of a swimming fish where the view moves at the same velocity as the reference frame fixed at the nose. The total length of fish is l, and the shape of the cross section is approximated by an ellipse. In Figure 2, the x-axis is the direction from rostral to caudal, the y-axis from midline to lateral margin, and the z-axis from horizontal septum to dorsal margin. The spine motions
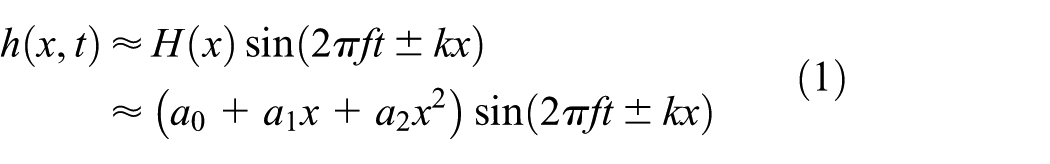
here, x is the distance from the nose in the longitudinal direction; t is the time; f is the flapping frequency;

Variables used to describe kinematics and cross section of swimming fish.
For the mechanical model of swimming fish, some quantitative parameters are needed to describe the structural property, such as the cross-sectional radii, mass, and second moment of inertia. Taking one unit
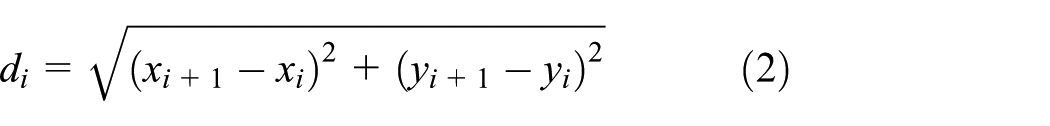
The mass of the rigid body,

where
In general, fish generate thrust by transferring momentum to the surrounding fluids. However, the mechanism of this momentum transfer is quite complex, and simplified models are often used in designing robotic fish (see review paper by Triantafyllou et al.
35
). In this study, the classical model based on Lighthill’s
31
slender-body theory is used to calculate the lateral hydrodynamics around the swimming fish. The lateral force

where
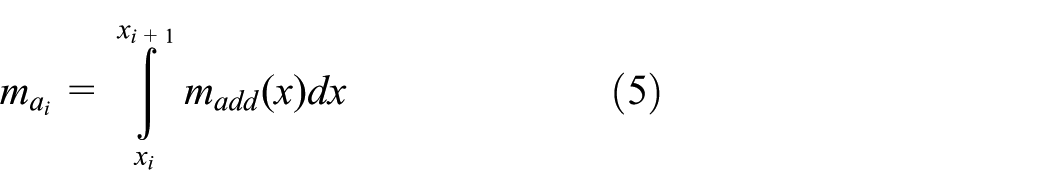
The lumped second moment of inertia

where
Design of the body stiffness
Fish achieve notable swimming performance by modulating their body stiffness actively to match their tail-beat frequency. 36 This oscillatory motion of swimming fish is related to their passive mechanical properties. However, the details of the body stiffness properties and the mechanism that associates them with the morphological parameters and swimming modes are still unknown.
In this work, the amplitude envelope of swimming motions is used to design the body stiffness profile at a certain driving frequency. For the fish model using the serial–parallel mechanism, it deforms to fit this amplitude envelope through the rotation of each serial–parallel unit independently. The amplitude envelope

Therefore, when each serial–parallel unit

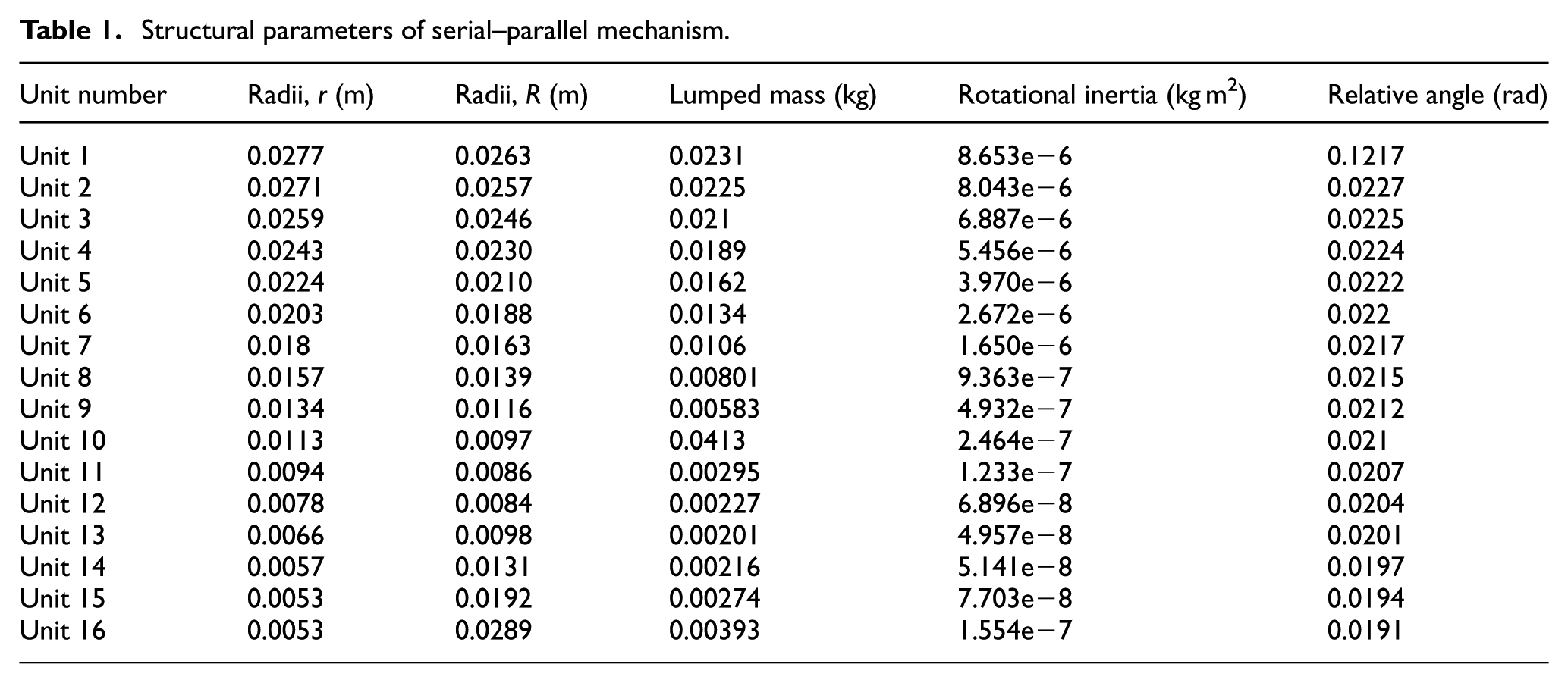
The specific values of structural parameters are calculated by equations (1)–(6) based on the morphological parameters. For the serial–parallel unit

with




here,

Dynamic analysis of the planar serial–parallel unit.
When the first serial–parallel unit is actuated by a sinusoidal moment, the dynamics equation of any other unit

with


here,
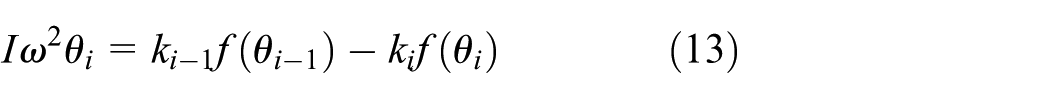
As presented in equation (13), the specific mechanism for the transfer of elastic force to the posterior serial–parallel units is related to the morphological parameters and the swimming mode. The terminal serial–parallel unit is only attached to two springs, and its dynamics equation is written as

The dynamics equations can also be written in matrix form
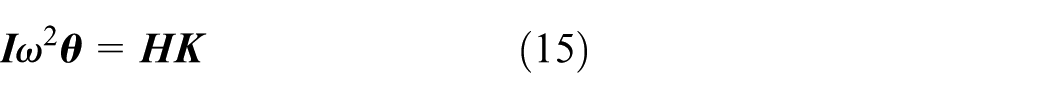
with


The stiffness profile can be solved by backward substitution. For the selected or mimicked fish, the structural parameters of the serial–parallel mechanism, or the matrix
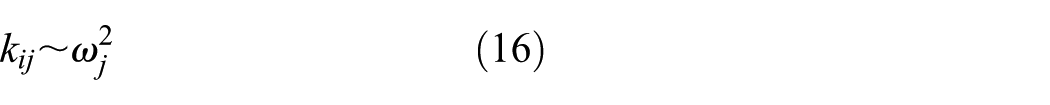
where

This pattern of body stiffness is in a manner consistent with the forced vibrations of oscillating beams. 37 Note that the ratio of the tail-beat frequency to the square root of the body stiffness remains constant, independent of the body mass, geometry (added mass), and damping. This indicates that fish prefers to swim at a frequency around its resonance frequency at the minimum mechanical bending cost. 17 Consequently, if the resonance frequency equates its tail-beat frequency, the swimming fish use their muscles and tendons to stiffen their body passively, and hence increase the tail-beat frequency. Therefore, the swimming thrust or power is controlled by tail-beat frequency.
Here, a general method based on the planar serial–parallel mechanism is proposed to determine the body stiffness at a wide range of swimming frequencies. The number of units and their longitudinal distances are the basic parameters to design the serial–parallel mechanism for the model of swimming fish. In this method, the amplitude envelope is regarded as the design curve. The required body stiffness can be found by minimizing the error between the required deflection and the lateral deflection of selected fish. Certain design guidelines should be followed as a compromise between the computational efficiency and the anatomical reality, including the length of flexible body, wavenumber, and body shape. In general, the length of flexible body varies with the type of body and/or caudal fin (BCF) locomotion. For example, anguilliform swimmers have a large ratio of flexible body, while thunniform fish have a rigid anterior part, and their posterior parts are flexible. However, the wavenumber of midline motion is variable for different types of fish. When the wavenumber is large, it means that the fish body is more flexible, and more units may be needed to mimic the swimming motions.
From a practical view, the more serial–parallel units the robotic fish use, the more energy is consumed, because the dampers connecting serial–parallel units would increase the mechanical friction and reduce the swimming efficiency of the whole system. However, the amplitude envelope can be mimicked very well by more structure units. As mentioned above, the fish model is driven by only one actuator, and the actuator should be embedded into the body to produce the motions. The residue length is replaced by the serial–parallel mechanism right after the actuator’s position. To a large extent, the actuator position determines the residue length of serial–parallel mechanism. The longer the residue length, the more units the serial–parallel mechanism needs.
A numerical example
Real fish provide many useful information, including swimming modes, mechanical properties (e.g. inertia and stiffness), and body shape (morphology),2,5,38 for designing a robotic fish. Here, a thunniform fish with a length of 0.26 m is selected as a numerical example to illustrate the design of robotic fish. In general, the thunniform fish body is stiffer than other BCF locomotion modes, that is, the wavenumber of body motion is small, and the thrust is done by the rear half of the fish body. The spine motion, or the lateral deflection
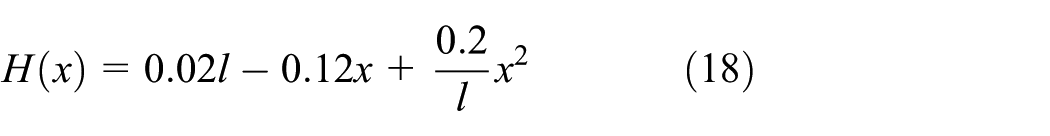
The cross sections of thunniform swimmers 24 can be modeled as


These geometric parameters
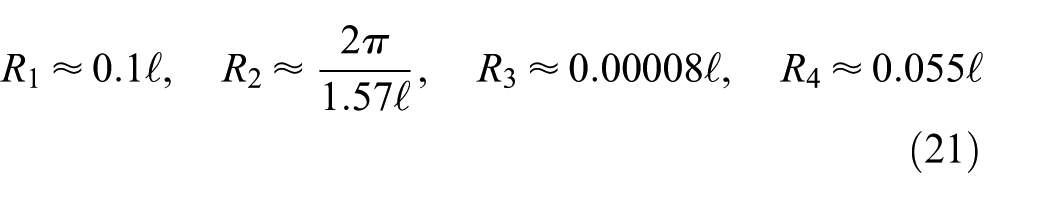

For thunniform fish, the posterior part of fish body and caudal fin are known to generate the principal thrust for the propulsive locomotion, while the anterior part remains relatively rigid. Therefore, the design of fish model focused on the undulation of the tail with defined length of 0.16 m,
Structural parameters of serial–parallel mechanism.
When the modeled fish with the planar serial–parallel mechanism rotates to match the amplitude envelope at a driving frequency of 2 Hz, the relative angle

Figure 4 shows the body stiffness of each serial–parallel unit when actuated at the tail-beat frequencies of 2 and 3 Hz.

Stiffness profile of the serial–parallel units at design frequencies of 2 and 3 Hz.
The SimMechanics model of robotic fish
A novel SimMechanics model of robotic fish is built to mimic its steady rectilinear locomotion based on the biological structure of real fish, as shown in Figure 5. The simulation model consists of two parts: the anterior part (including the head) and the posterior part (including the caudal fin). In the oscillating motion, the anterior part of body is simplified as a rigid body with a lumped mass, while the posterior part consists of 16 serial–parallel units. In the simulation model, the structure parameters are the same as those in the numerical example.

Top view of simulation model with the planar serial–parallel mechanisms.
For simplicity, only three units of the model are used to represent the mechanism, and the schematic diagram of the SimMechanics model is presented in Figure 6. In general, the model relies on six functional modules: (1) the drag module, used to add the drag force; (2) the actuation module, used to add the sinusoidal torque; (3) the vertebra module, using rigid flat plates to mimic the vertebra linked by revolute joints; (4) the fluid module, used to produce the reactive force; (5) the muscle module, using the damper and spring to account for the mechanical property of viscoelastic muscles and produce the relative velocity and the acceleration; and (6) the detection modules, used to monitor and record changes in the lateral amplitude and the forward speed. The forward speed U is further fed back to the model as the drag force D(

Schematic diagram of fish structure in SimMechanics model.
From the aspect of structure, the vertebras are linked in serial by resolute joints, as shown in Figure 7. The joints are located at the midpoint of each vertebra, while the muscles are distributed along two sides, and they interact with the local fluid environment. According to Lighthill’s elongated theory, the fluid force around each unit is different due to the added mass and its acceleration. So, in the muscle module, both the viscoelastic property of muscles and the interactive force of fluid environment are considered in our SimMechanics model, as showed in Figure 7.

Serial vertebra linked by revolute joints.
In Figure 8, at each joint, the speed

Mechanical structure of muscle module.
In a steady rectilinear locomotion, provided a sinusoidal torque at the actuation joint, the virtual fish generates the thrust, which is the vector component of lateral force in forward direction. According to Lighthill’s
31
elongated-body theory, the lateral force for each serial–parallel unit is determined by the product of the second derivative of lateral line
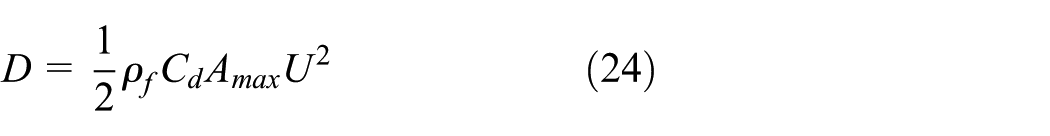
where
As shown in equation (24), when a fish freely swims in the water, the drag coefficient,

where
Analysis of the robotic fish
As mentioned above, the swimming speed and Strouhal number are two useful indexes to evaluate the swimming performance of a robotic fish. In the following analysis, three physical issues are investigated quantitatively to account for the effects of body stiffness: (1) whether the centerline in the simulation can match with that of a real fish by adjusting the body stiffness with only one actuator; (2) how the swimming performance of the robotic fish with the prescribed stiffness changes with various driving frequencies; and (3) how the swimming performance changes when the robotic fish is driven at different resonance frequencies, which correspond to the prescribed stiffness.
Fitting curve of the lateral deflection
A sinusoidal torque with frequency 2 Hz is applied to the first serial–parallel unit, and the flexible body of robotic fish with the design stiffness oscillates in the xy plane. The peak lateral amplitude of each serial–parallel unit is monitored and recorded during the entire simulation period. For example, the time histories of the amplitude evolution of the no. 3, no. 10, and no. 16 unit are shown in Figure 9. The maximum amplitude of each unit is compared with the amplitude envelope in Figure 10, and they are in good agreement with each other.
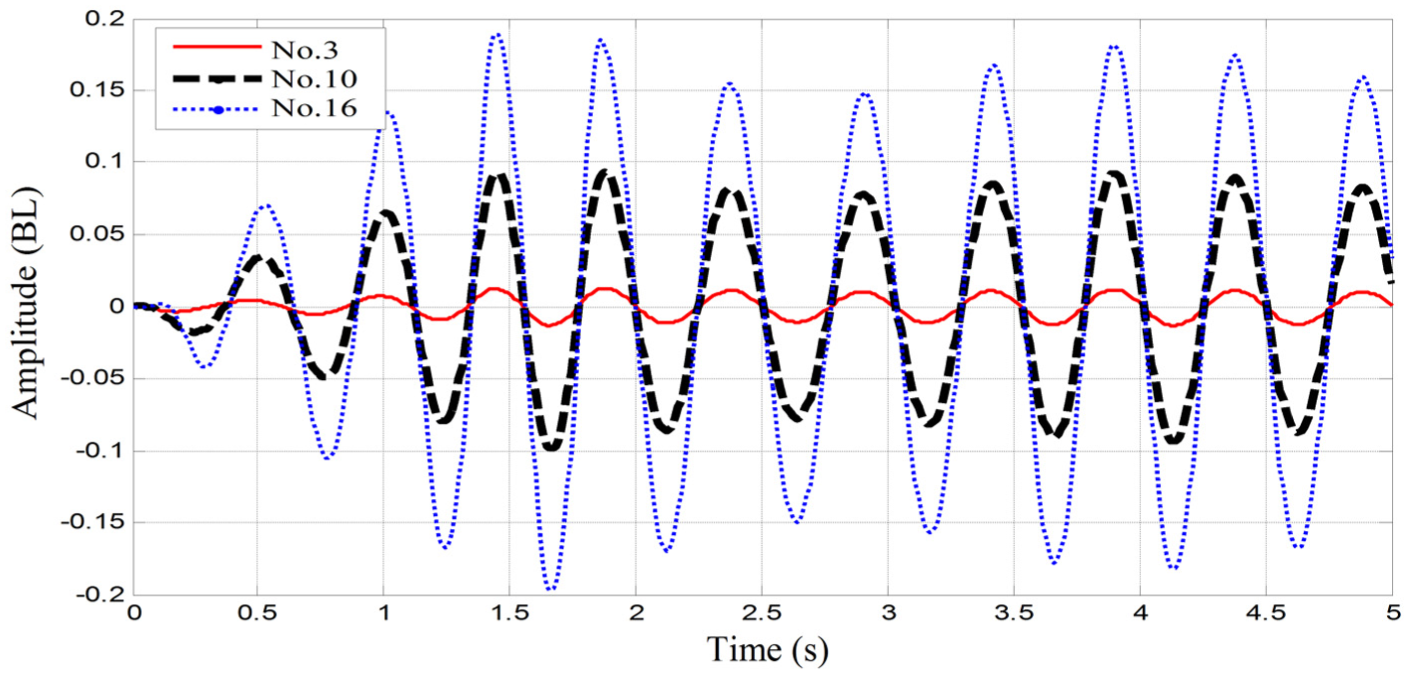
Amplitude history of three different units in SimMechanics model.

Amplitude envelopes of lateral deflection in simulation and design.
Swimming performance versus driving frequency
The robotic fish and the surrounding fluids can be conceptualized as a simple mass–spring system, and the body stiffness is similar to a spring. The robotic fish thus behaves differently when actuated at a frequency much less than, in tune with, or much greater than its resonance frequency. In our analysis, when the resonance frequency is 2 or 3 Hz, the robotic fish with the design stiffness is actuated at different driving frequencies at intervals of 0.5 Hz from 0.5 to 5 Hz. Different forward speeds and maximum oscillating amplitudes of tail are recorded.
Figure 11 shows the relationship between the forward speed and the driving frequency. As expected, when the driving frequency is close to the resonance frequency, the forward speed reaches the highest value. The maximum forward speed of the robotic fish at the design frequencies of 2 and 3 Hz reaches approximately 1.2 and 1.9 body lengths per second (BL/s), respectively. The similar phenomenon is also found in a numerical study for jellyfish. 41 Moreover, the change pattern of the forward speed in Figure 11 is consistent with the experimental results of a compliant fish-like robot made by Valdivia y Alvarado and colleagues.24,25Figure 12 shows the changes in Strouhal number with different diving frequencies. Results show that Strouhal number decreases with the driving frequency. When the driving frequency approaches the design frequency 2 and 3 Hz, the Strouhal number is about 0.28 and 0.29, respectively. These values are consistent with biological results 0.25 < St < 0.35, suggesting the efficient swimming of true fish.

Swimming speed versus driving frequency.

Strouhal number versus driving frequency.
The results in Figures 11 and 12 show that the robotic fish has a better swimming performance when the driving frequency is close to the resonance frequency. They also suggest that the resonance frequency is equal to the design frequency, which is closely related to the body stiffness. In other words, the results verify the effectiveness of the method in designing body stiffness, and the proposition to adjust body stiffness at different driving frequencies.
Swimming performance versus resonance frequency
In nature, the swimming speed of fish is directly proportional to the flapping frequency of tail.34,42 Inspired by this fact, we change the design frequency in intervals of 0.5 Hz from 0.5 to 5 Hz and calculate the body stiffness according to the principle of adjustable stiffness. Then, the model with different body stiffness is simulated at the corresponding design frequency, or resonance frequency. The results of the swimming speed and Strouhal number are presented in Figures 13 and 14, respectively. In Figure 13, the forward speed is linearly proportional to the driving frequency, and the forward speed reaches 2.8 BL/s at a driving frequency of 5 Hz. These forward speeds of the robotic fish are qualitatively consistent with biological data summarized by Videler. 34 The Strouhal numbers in Figure 14 are between 0.28 and 0.34, which are consistent with the range 0.25 < St < 0.35 required for the optimal efficiency. In general, these results show that the robotic fish can achieve superb swimming performance by adjusting the body stiffness and the driving frequency. They also suggest that during swimming, fish, in nature, may change their body stiffness the with tail-beat frequency. This assumption still needs more support from biological experiments although the effects of resonance frequency on swimming fish have been analyzed.13,15,35

Forward speed versus different resonance frequency.
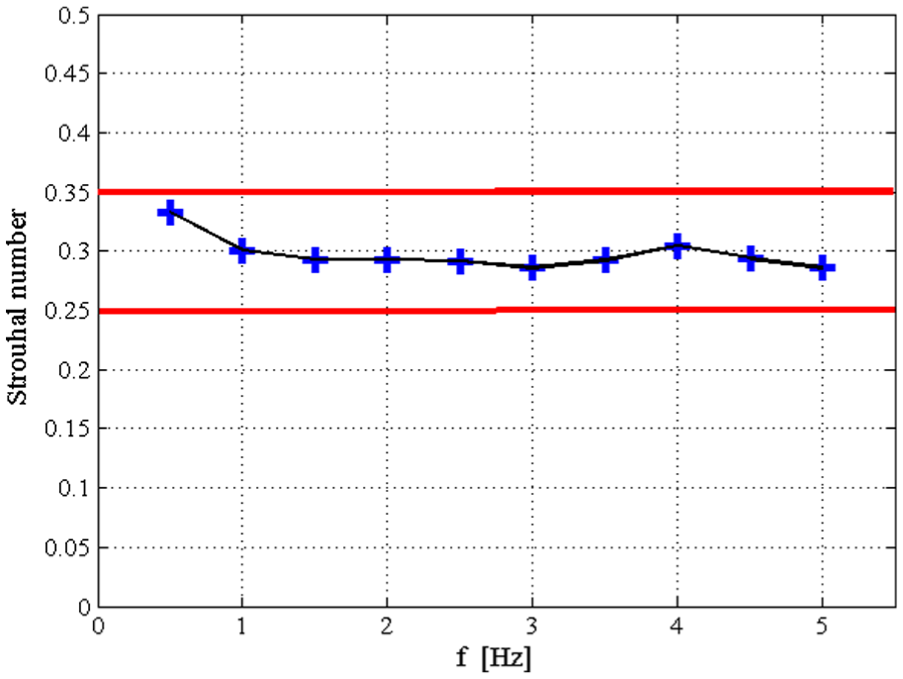
Strouhal number versus different resonance frequency.
Overall, the robotic fish using the planar serial–parallel mechanism has a great potential to mimic the swimming performance of true fish. As seen in Figures 13 and 14, the results validate the proposed method of calculating body stiffness, and verify the way to adjust body stiffness. Although the conceptual design of the robotic fish with a planar serial–parallel mechanism has been presented, there are two main challenges in conducting an experimental verification. The first challenge is that it is difficult to fabricate a robotic fish by assembling the delicate devices used to build a serial–parallel mechanism in a limited space, let alone making the fish swim steadily in water. Another one is lacking a detailed method to change the body stiffness quickly and accurately. Fortunately, many novel applications of the variable impedance actuators (VIAs) are proposed for the devices with adjustable stiffness (see review paper by Vanderborght et al. 43 ). For now, Nakabayashi et al. 44 used a variable effective-length spring to develop a variable-stiffness fin, and Huh et al. 45 built a compliant fin of robotic fish to increase the thrust by adjusting the structure stiffness. These VIAs can be controlled actively, which provide inspirations to build a robotic fish with adjustable stiffness.
Conclusion
Inspired by the biological structure of fish, this article proposes a planar serial–parallel mechanism to design the robotic fish and presents a general method for calculating the body stiffness at a wide range of swimming frequencies. According to Lighthill’s elongated theory, the reactive force from the fluid environment is fully considered in the proposed model. The stiffness profile of fish body is prescribed from the morphological parameters, and the motion of the fish lateral line can be controlled by adjusting the body stiffness with only one actuator. Furthermore, an innovative SimMechanics model of a robotic fish is built to describe the steady rectilinear locomotion. Six functional modules, namely, the drag module, the actuation module, the vertebra module, the fluid module, the detection modules, and the muscle module are included in this model. The swimming speed and Strouhal number are used to evaluate the swimming performance of a robotic fish.
The stiffness profile is prescribed when the tail-beat frequencies are 2 and 3 Hz, which is based on the biological parameters of thunniform fish. The fish is driven with different frequencies at intervals of 0.5 Hz from 0.5 to 5 Hz and has different speeds and Strouhal numbers. The simulations show that when the driving frequency is close to the resonance frequency, the fish with the design stiffness has the maximum speed (1.2 BL/s at 2 Hz and 1.9 BL/s at 3 Hz), and the maximum speed is linearly proportional to the resonance frequency. The range of the Strouhal number (0.28–0.34) is also consistent with the range 0.25 < St < 0.35 required for the optimal efficiency. Both the swimming speed and Strouhal number are consistent with the data from biological observations. These results prove that the fish model using the serial–parallel mechanism is effective. Overall, this work presents a theoretical basis for the design of robotic fish having excellent swimming performance by adjusting the body stiffness so as to match the driving frequency, or resonance frequency. It is also expected to be a useful instrument in the optimal design of fish-like robot.
Footnotes
Academic Editor: ZW Zhong
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This article is financially supported by the National Natural Science Foundation of China (project no. 51275127).