
Abstract
Introduction
Micronutrients, although required in small quantity, their deficiency leads to low productivity in many areas of the world. Among micronutrients, Zn is considered to be the most important as its deficiency causes ubiquitous abiotic stress in Asian countries like Pakistan, India, and Bangladesh and also other countries like Turkey, Syria, Iraq, and also in USA and Africa. 1 Zinc deficiency in agriculture not only affects crop yields but also nutritional quality of the crops, and crop products are frequently impaired that causes nutritional and human health issue, especially in children.2,3 In plants, zinc plays a key role as a structural constituent or as a regulatory co-factor of a wide range of different enzymes and proteins involved in many important biochemical pathways, mainly concerned with photosynthesis, in the conversion of sugars to starch, protein metabolism, auxin (growth regulator) metabolism, pollen formation, and the maintenance of the integrity of biological membranes. 4 Zinc deficiency is exhibited by calcareous, alkaline, and sodic soil having low organic matter and high pH which lead to drying of the upper soil horizon.5,6 Meanwhile, intensive cropping with poor drainage is one of the reasons of zinc deficiency in soil. 4 Synthetic fertilizers in the form of zinc sulfate or Zn EDTA appear as an easy and quick way to rectify the Zn deficiency, but it is non-sustainable and their use puts an economic burden.7-12 Meanwhile, to increase zinc availability, fertilization of food crops with Zn represents a short-term and complementary strategy, which is necessary to build the Zn pool for uptake or translocation. Unfortunately, application of chemical fertilizers often has low use efficiency that means plants can uptake just a small portion of applied nutrients as only a small part of Zn is present in soluble form in soil. To combat soil Zn deficiency, in long term, is not possible only with chemical fertilizers as 96–99% of the applied Zn is converted to unavailable form due to formation of carbonates, oxides, or phosphates depending on soil type.13,14
However, the worldwide Zn deficiency in crops is not only due to low Zn level in soil but rather due to low solubility of Zn in soil and conversion of Zn in insoluble form. 15 It is reported that low amount of bioavailable Zn in soil not only leads to low productivity of crops but also lower nutritional quality of the plants. 16 Zn fertilizers applied in the fields as soluble form, like zinc sulphate, is transformed into various insoluble forms such as ZnS in reducing conditions predominantly in flooding, zinc phosphate in neutral to alkali soils having number of applications of phosphate fertilizers, ZnCO3 in calcium-rich alkali soils, and Zn(OH)2 in high pH soil, thus supporting the evidence of Zn deficiency in soil and ultimately in plants due to insoluble forms. 17
In Pakistan, soils are normally of alkaline and calcareous nature that is more prone to Zn deficiency, as in these soils, Zn could precipitate into the unavailable forms. 18 Inorganic fertilizers are recommended as good source of Zn but they are quickly fixed on soil medium, causing poor availability to plants.19,20 The application of zinc sulfate (ZnSO4) in the form of fertilizer might decrease Zn deficiency and increase plant yield. This unavailable form of zinc can be reverted back to available form by inoculating a bacterial strain capable of zinc-solubilizing potential.
Therefore, an attractive alternate would be to exploit the microorganisms having potential to solubilize the fixed forms of Zn present in soil to bioavailable forms to cater plant Zn requirement. Many bacterial genera like Pseudomonas sp., Burkholderia, Gluconacetobacter sp., Thiobacillus sp., Bacillus sp., and Acinetobacter sp. have been reported in vitro to solubilize the Zn insoluble forms,21-24 although to solubilize ZnO, ZnCO3, or ZnPO4 (insoluble forms of Zn) is not a common characteristic of cultivable bacteria. Zn-solubilizing bacterial strains have the ability to enhance growth and yield of crops due to diverse direct and indirect mechanisms involved to effect the growth. The direct mechanisms involve solubilization of nutrient/minerals, stimulation of biological nitrogen fixation, siderophores production, ACC deaminase activity, and production of phytohormones, while indirect mechanisms involve antibiotics production, exopolysaccharides, and antifungal activity. The Zn-solubilizing bacterial strains solubilize the unavailable form of zinc by producing chelating ligands; secreting organic acids, vitamins, and phytohormones; and through oxido-reductive systems and proton extrusion. 23 Organic acid poduction by bacterial strains is a major mechanism used for Zn solubilization. Among organic acids, 2-ketogluconic acid and gluconic acid production by PGPR is responsible for solubilization of Zn. 22 Use of Zn-solubilizing strains in crops could not only be an alternate to chemical Zn fertilizer application but also deal with the malnutrition problem through increased nutrient concentration in grains.
Keeping in view the above facts, this present study was designed to isolate, screen, characterize, and identify the zinc-solubilizing bacterium and to evaluate the its impact (if any) on the growth parameters and zinc content of chickpea plants through pot experiments. In addition to evaluating the Zn-solubilizing potential, the selected isolates were also screened for their multiple abilities such as solubilization of P, K, Si, production of indole-3-acetic acid (IAA), NH3, HCN, and siderophore.
Materials and Methods
Isolation of Zinc-Solubilizing Bacterial Strains
For isolation and screening of Zn-solubilizing bacteria, 50 soil samples were collected from agricultural fields of maize in the vicinity of industrial area (Paharang drain) of Faisalabad (Pakistan), which had been irrigated with waste water from industries. The samples were collected in the sterile, dry polythene bags and transported to the laboratory for further analysis within 6–8 hours of collection. Soil samples were air dried ground until they pass through a 2 mm sieve. Then, sterile water was added in soil samples and shaken at 150 rpm for 2 hours. After that, samples were settled for 5 minutes and 1 mL of the suspension was serially diluted up to 10−6. Each dilution was spread onto nutrient agar plate supplemented with 1 mM Zn and incubated at 30°C. The growing colonies were observed and 10 out of 35 bacterial colonies with different morphology were selected and purified by repeated streaking on nutrient agar. 25
Qualitative and Quantitative Zn Solubilization Assay
The zinc-solubilizing ability of these 10 strains was evaluated in mineral salt medium (MSM: NaCl 1g, CaCl2 0.1 g, MgSO4 0.5 g, KH2PO4 1 g, K2HPO4 1 g, yeast extract 4 g, agar 16–18 g in 1 L, and pH was maintained at 7.2). 26 Different sources of insoluble zinc salts like zinc oxide (1.244 g/L = 15.23 mM), zinc phosphate (1.3882 g/L = 4.0 mM), zinc carbonate (1.4728 g/L 11.74 mM), and zinc sulfide (1.124 g/L 11.54 mM) at final concentration of .1% were added in the medium individually and autoclaved at 121°C for 30 minutes. 27 A loop full of bacterial overnight growth in nutrient broth was spread on MS medium. Plates were incubated at 30°C for 7 days. Strains showing a clear zone around the colony are considered as zinc-solubilizing strains. The halo zone was measured to determine the zinc solubilization efficiency (ZnE) of the strains, using method 28 :
ZnE = (HZ/C) × 100, where
ZnE is the Zn solubilization efficiency, HZ the diameter of the solubilization halo zone, and C is the diameter of the colony.
For quantitative Zn solubilization assay, MSM broth was prepared and supplemented with .1% insoluble zinc compounds. One ml of overnight grown culture was inoculated into each flask containing 50 mL of MSM broth and then incubated at 30oC for 15 days on shaking at 150 rpm (Thermo Scientific™, UK). 10 mL of samples were withdrawn after 0, 2, 5, 10, and 15 days of incubation and centrifuged at 10 000 rpm for 10 minutes (Thermo Scientific™, UK). After that, 1 mL of supernatant was transferred to 50 mL flask and distilled water was used to make volume up to 50 mL. This sample was used to determine the available Zn using atomic absorption spectrophotometer (AAS) (Shimatzu AAS-7000) at 213.9 nm. 29 Highly zinc-soluble strain was selected. Medium without culture was maintained as control. pH of each culture and control was also determined by using pH meter. Experiments were performed in triplicate.
In Vitro Screening of Zn-Solubilizing Isolate for Their Growth Promoting Characteristics
Indole-3-Acetic acid production
For IAA production, Zn-solubilizing bacterial strains were inoculated in LB broth with the addition of .1% L-tryptophan as precursor for IAA and incubated at 28oC for 96 hours at 120 rpm. After incubation, the broth was centrifuged at 10 000 rpm for 10 minutes and supernatant was collected. One mL of Salkowski’s reagent 30 was added in the 1 mL of supernatant in 2 mL Eppendorf tube and samples were kept in dark for 25 minutes to observe color change. The absorbance was taken at 530 nm (Halo DB-20 UV–vis double beam spectrophotometer). 31 Calibration curve of pure IAA was used as a standard.
Siderophore Production
For determination of siderophore production, the chrome azurol-S (CAS) method was used as described by Perez-Miranda et al. 32 Bacterial culture was inoculated on agar medium and incubated at 28°C for 24 hours. Siderophore production was determined by production of orange halo zone around the bacterial isolate on blue. 33
HCN Production
Qualitative method of Kloppers et al. 34 was used for determination of HCN production with few modifications. Bacterial strains were inoculated in DF minimal salt medium supplemented with 4.4 g L−1 glycine. Filter paper strips soaked in picric acid (.5%) and sodium carbonate (2%) solution were placed in half of the vials and sealed with parafilm. Samples were incubated for 96 hours at 28°C for 72 hours in a mechanical shaker. Change in color of the filter paper from yellow to orange brown is indicator for HCN producing strain.
ACC Deaminase
The production of α-ketobutyrate (α-KB) by the enzymatic cleavage of ACC for 10 selected bacterial strains was determined by measuring its absorbance in bacterial culture at 540 nm and comparing with the absorbance of known concentration of pure α-KB0020. 35 Protein concentration in bacterial culture was estimated by the method of Bradford 36 and then the enzymatic activity was expressed as 1 M αKB mg 1 h1. 37
NH3 Production
NH3 production was measured by inoculating the bacterial strains in peptone broth (4%) and incubated at 25°C for 96 hours. After incubation, 1 mL of Nessler’s reagent was added. Development of yellow to dark brown color was considered as positive indicator for NH3 producing strains. 38
Solubilization of P, K, and Si
For measurement of solubilization of P, bacterial strains culture was inoculated on Pikovskaya’s broth for 7 days at 150 rpm. After incubation, culture was centrifuged at 6000 rpm and supernatant was used to determine P solubilization using the calorimetric method. Color reagent (500 mL ammonium molybdate, sulfuric acid solution, and 125 μL of ferrous sulfate solution) was prepared and 750 mL added to the obtained supernatant and phosphomolybdate production was measured at 700 nm (UV–vis double beam spectrophotometer).
For the measurement of K solubilization, bacterial strains were inoculated in Aleksandrov medium 39 and clear zone was observed after incubation at 30°C for 96 hours.
For the measurement of Si solubilization, basal medium (20 g glucose, .1 g magnesium sulphate, .2 g potassium chloride, 1 g ammonium sulphate, and .1 g potassium dihydrogen phosphate in 1 L) was used containing 5.0 g of potassium alumina silicate (insoluble silicate). After solidification, well was made in the middle of Petri plates and .1 mL of the freshly prepared bacterial strains suspension was poured into the well followed by and incubated for 5 days at 28°C. Afterward, plates were observed for clear zone (mm) around the wells. 40
Identification of Selected Strain
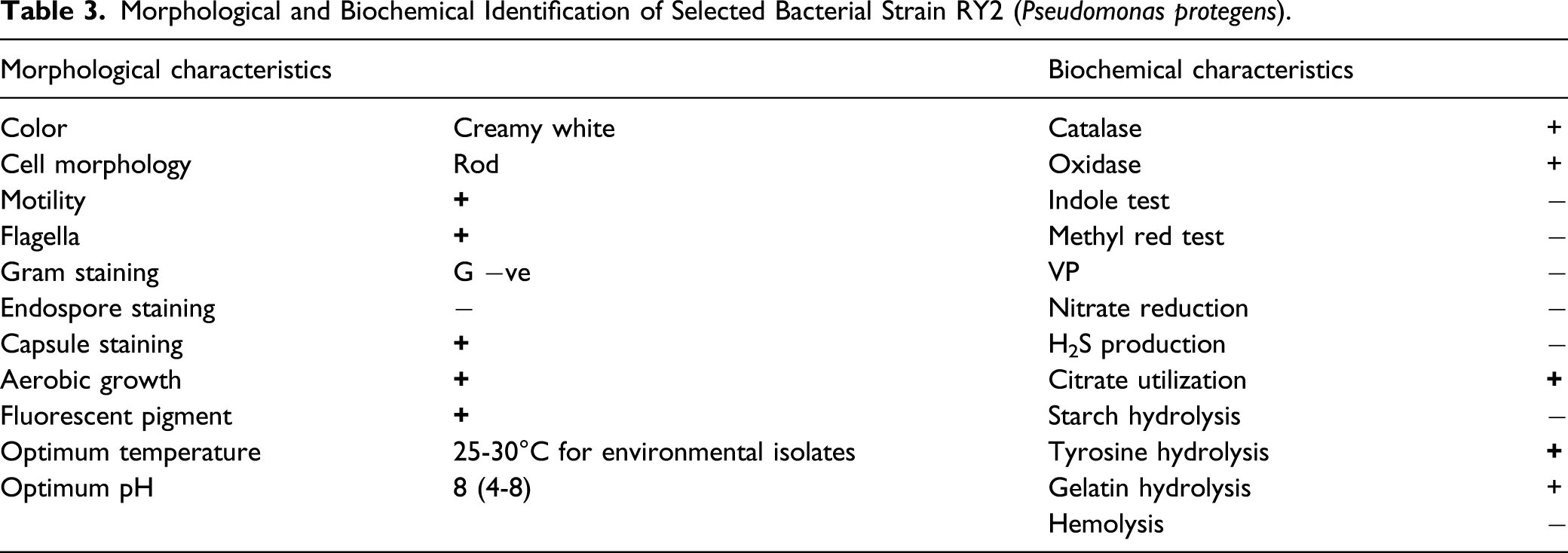
On the basis of above traits, strain RY2 was selected for further study in plants. Bacterial strain RY2 identification was done on the basis of cultural characteristics, gram’s staining, and biochemical characteristics like oxidase reaction, citrate utilization, catalase reaction, etc.
The selected bacterial strain was identified through 16S rRNA gene sequencing. The standard protocol of Cheneby et al. 41 was followed to extract bacterial DNA. Universal primers 785F (5′ − GGATTAGATACCCTGGTA −3′) and 907R (5′ − CCGTCAATTCMTTTRAGTTT −3′) were used. 42 PCR was carried out with the help of thermo cycler (Advance Primus 96) using initial denaturation at 94oC for 3 minutes, 35 cycles of denaturation at 94°C for 1 minute, primer annealing at 53°C for 30 seconds, DNA extension at 72°C for 2 minutes, and final extension at 72°C.
For ribotyping, PCR products were sequenced using commercial service of Macrogen, Korea. The 1425 bp sequences obtained were checked for base calling using FinchTV and contigs made using NCBI BLAST (2 sequence alignments). After amplification, 16S rRNA sequence was aligned using ClustalW and compared with known sequences in the GenBank database. The phylogenetic analysis was performed as described by Roohi et al. 43 using MEGA version 7.0.14.
ZnSB-Mediated Zn Release in Soil
To determine the Zn release, soil was treated with RY2 strain. For the study, soil was sieved and sterilized. Then, .06 g of ZnO was added to 60 g soil (1000 mg/kg), and transferred to pots. After that, 3 mL of freshly grown culture of RY2 was inoculated and pots were kept in incubator and moisture was maintained. Control was also maintained without RY2 strain inoculation. Four replicates were maintained for each treatment for sampling. Available Zn was determined after 2, 5, 8, 12, 15, and 20 days. At each day of sampling, 10 g of soil from each pot was taken and mixed with 20 mL of diethylenetriaminepentaacetic acid (DTPA) extract, shaken for 2 hours, and filtered through Whatman 42 filter paper. The zinc concentration was analyzed in the filtrate by atomic absorption spectrophotometer (Shimatzu AAS-7000) as described by Wenzel et al. 44
Plant Experiments
To analyze the effect of RY2 on chickpea growth, pot experiment study was conducted using different treatments as follows: T1 (Control: soil without Zn), T2 (soil without Zn + strain RY2), T3 (soil + ZnSO4.), T4 (soil + ZnO), T5 (soil + ZnO + strain RY2), T6 (soil + manure + ZnO), and T7 (Soil + manure + ZnO + strain RY2). For RY2 application, 20 mL of freshly grown culture (∼×108 cfu/mL) was applied. Chickpea (C arietinum) seeds were obtained from Ayub Agriculture Research Institute, Faisalabad, Pakistan. Seeds were surface sterilized with 70% ethanol for 1 minute. Then, they were rinsed with sodium hypochlorite solution for 3 minutes and washed with distilled water. Seeds were germinated in sterilized soil, and after 1 week, seedlings were transferred to earthen pots containing 2.5 kg of soil using sterilized forceps. 200 g/pot animal manure has been used that normally contains essential nutrients like N, P, K, etc.
Pots were placed in a growth room with controlled conditions, that is, temperature: 28 ± 2°C, photoperiod: 16 hour-/8 hour-light/dark, and relative humidity: 52 ± 2% in a randomized complete block design (RCBD). 45 To provide moisture, plants were watered at alternate days using autoclaved distilled water. Two sets of experiments (each with 3 replicates) were carried out. After 60 days of germination, chickpea seedlings were harvested and data related to root length, shoot length, root fresh and dry weight, shoot fresh and dry weight, and total fresh and dry biomass were recorded.
Analysis of Zn Content in Soil and Plant Using Atomic Absorption Spectrophotometer (AAS)
Zinc concentration in roots, shoots, and leaves of 4 plants per treatment was analyzed for zinc estimation. 1 g of plant powder was digested in di-acid mixture (HNO3: H2SO4) in the ratio of 2.5:1). Sample was heated at 70°C and diluted with 50 mL distilled water. Then, extract was filtered through a Whatman’s filter paper and filtrate was analyzed for available zinc content using AAS. 46 For soil, extraction was done with DTPA and Zn content was measured in the extract using AAS. Zn content in soil was determined at 5, 15, 30, and 45 days in each treatment. 47
Statistical Analysis
In this study, ANOVA was employed to analyze the data, and in case F values were significant, the least significance test at the .05 probability level was applied.
Results
Isolation of Zinc-Solubilizing Bacterial Strains
A total of thirty-five strains were isolated from the samples. These strains were purified and coded as RY1 to RY35. Ten isolated strains (RY2, RY7, RY10, RY12, RY14, RY19, RY24, RY25, RY26, and RY27) were selected (based on different morphology) for further experimentations. The texture of soil was sandy loam with good internal drainage. The pH of soil was <8, 20 with 160 mg/kg of Zn content.
Qualitative and Quantitative Zn Solubilization Assay
Efficacy of Rhizobacterial Strains to Solubilize Zinc Insoluble Salts Under Laboratory Conditions.
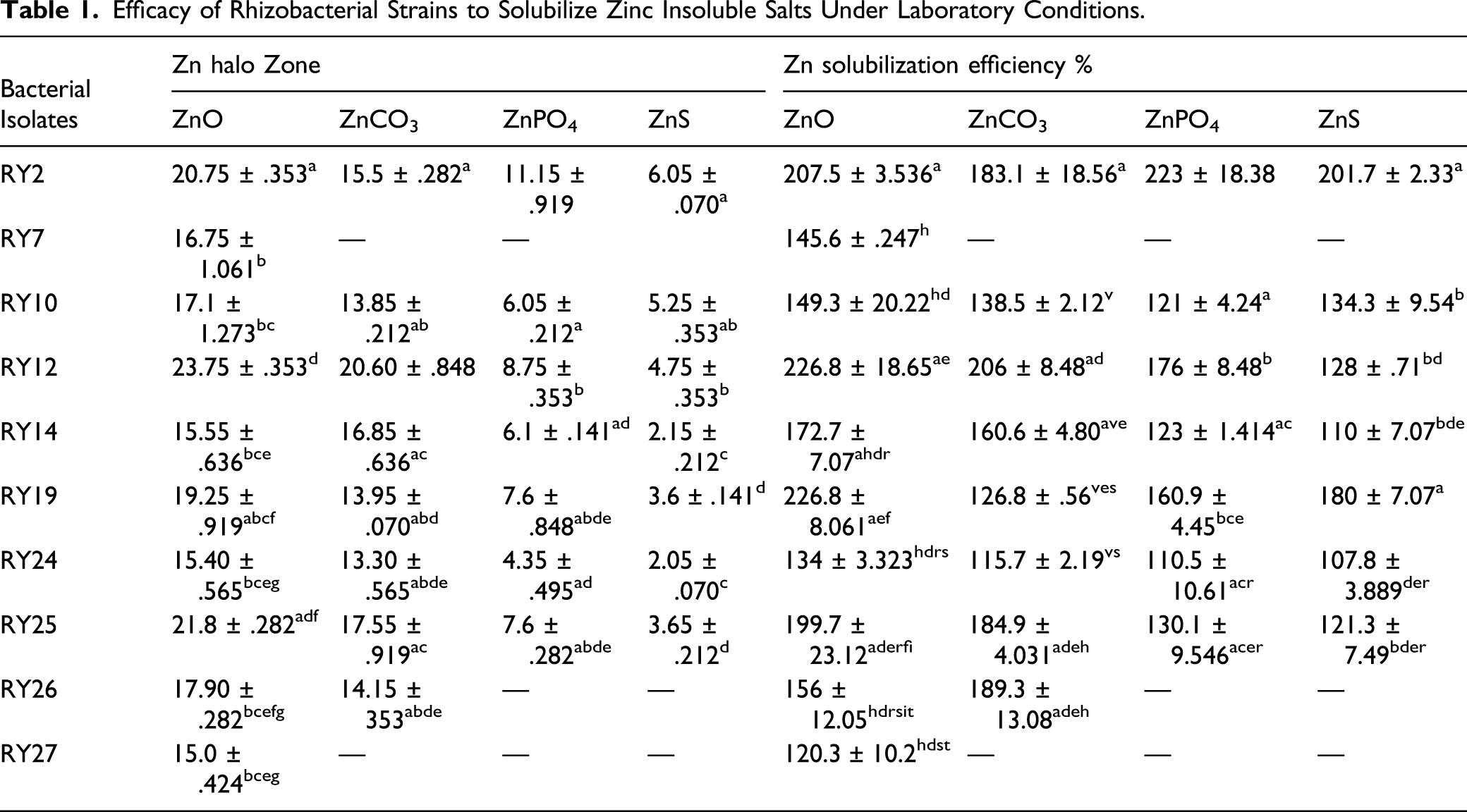
Bacterial strains were further assessed for their Zn solubilization potential in liquid medium at different time intervals. RY2 registered significantly higher (P < .05) available Zn at all days of incubation for insoluble salts. In case of all insoluble salts, the solubilization was peaked at day 15 and on day 20, and it was decreased in case of zinc oxide; maximum available Zn registered by RY2 was 49.45 mg/L, 90 mg/L, 113.13 mg/L, 142.35 mg/L, and 134.65 mg/L on the 2nd, 5th, 10th, 15th, and 20th day, which peaked to 142.35 mg/L on 15th day of incubation, and on 20th day, there was a reduction in zinc content (Figure 1A). In case of zinc sulphide, the maximum solubilization was shown by RY12 where maximum zinc content was shown as 48.85 mg/L at day 15 and on 20th day it reduced to 38.60 mg/L (Figure 1B). There were, however, no significant differences in the amount of Zn solubilized by the other bacterial strains, irrespective of the days of sampling. Other strains except RY2 and RY12 showed non-significant changes in zinc content in different days of incubation. Some strains showed solubilization potential in the 2nd day, and after that in all other days, these strains showed no increase in zinc content and some strains started solubilization after 10th day of incubation (Figures 1C and D). Compared to control, there was a slight decrease in pH of the medium with RY12 and the decrease was perceptible in the treatment with RY2 where they ranged from 5.20 (day 10) to 4.41 (day 15). In fact, the highest level of soluble Zn (142.35 mg/L) was registered by RY2 when the pH was 4.41 during the 15th day. The pH in other ZnSB treatments ranged between 6.40 (RY25) and 7.71 (RY10). Quantitative solubilization of zinc insoluble salts: (A) ZnO (B) Zn3(PO4)2 (C) ZnS (D) ZnCO3 by different bacterial strains at different incubation period.
In Vitro Screening of Zn-Solubilizing Isolate for Their Growth Promoting Characteristics
Growth Promoting Characteristics of Isolated Bacterial Strains.

ACC, 1-aminocyclopropane-1-carboxylate; HCN, hydrogen cyanide.
The symbol, − shows the absence of the trait, + represents the presence of the traits while ++ shows very good activity.
Identification of Selected Bacterial Strain RY2
Morphological and Biochemical Identification of Selected Bacterial Strain RY2 (Pseudomonas protegens).
Molecular characterization of selected strain was carried out using 16S rRNA gene amplification. The strain RY2 confirmed as Pseudomonas protegens. The nucleotide sequence of 16S rRNA genes of Pseudomonas strain RY2 was submitted to the GenBank under the Accession no. MF351762. Finally, evolutionary status of strain RY2 was confirmed by constructing phylogenetic tree using the maximum likelihood method (Figure 2). Phylogenetic tree deduced from the 16S rRNA gene sequences of Pseudomonas protegens RY 2 (Accession no. MF351762) by neighbor joining linear method, conducted with MEGA 7 software.
ZnSB-Mediated Zn Release in Soil
The study was performed to investigate the rate of Zn release in soils treated with the selected ZnSB strain RY2. An increase in available Zn level in soil treated with ZnSB RY2 has been observed up to the day 12 and after that decrease in available Zn. Net available Zn (difference between available zinc in the treated soil and untreated soil) in the soil was 39.1, 59.4, 115.8, 153.5, 137.5, and 122.5 mg/kg at day 2, 5, 8, 10, 12, and 15, respectively. It is observed that rate of available Zn release in the soil treated with RY2 was 26.18%, 38.6%, 74.39, 96.5%, 88.9%, and 81.6% greater than untreated soil (Figure 3). Zn release rate by zinc solubilzation strain RY2 in soil at different incubation time (days).
Plant Experiments
Effect of Zinc-solubilizing Rhizobacteria on Chickpea Growth in Pot Experiment.

Shoot length in (cm). Root length in (cm). Fresh and dry weights are in (g).
aT1 (control: soil without Zn), T2 (soil without Zn + strain RY2), T3 (soil + ZnSO4.), T4 (soil + ZnO), T5 (soil + ZnO + strain RY2), T6 (soil + manure + ZnO), and T7 (Soil + manure + ZnO + strain RY2).
Zn Content in Soil and Plant Using Atomic Absorption Spectrophotometer
Available zinc in soil was determined. It was observed that Zn content was gradually increasing from day 5 to day 30, while at day 45 it was high than day 15 but less than day 30. Zinc level was highest 13.01 ± .289 mgkg−1 in T7 at day 30 as compared to all other treatments where T1 (control) was 2.228 ± .412 mgkg−1. At day 15, non-significant differences were observed between T3 and T5 but a significant difference between T3 and T7. In case of T2 and T3, a non-significant difference was observed in all days of incubation (Figure 4). While in case of T2 and T5, a significant result was obtained at day 5 and day 15, while non-significant differences were observed at day 30 and 45. Zinc level was greater in T5 (12.06 mgkg−1) and T7 (13.01 mgkg−1) at day 30. Available Zn in the treatment T3 was lower at day 30 (10.42mgkg−1), though significantly (P < .05%) higher than T1 (2.228 mgkg−1). While comparing T3 at day 15 and day 30, a significant increase in available Zn was not observed in T3 up to day 15, but at day 30, no further increase was observed. Zinc in soil after at 45th day of chickpea growth.
Zn uptake was significantly higher in roots (P < .05) in the treatment T7 (1.717 mg/kg), T5 (1.35 mg/kg), and T2 (.927 mg/kg) as compared to T1 (.247 mg/kg) and T3 (1.493 mg/kg). There was significant increase in zinc uptake in T2, T5, and T7 when compared with T1 (control) but non-significant difference was observed when compared T5 and T7 to T3 (1.493 mg/kg), whereas when compared T2 and T3, there was significant increase in T3 rather than T2. Highest amounts of zinc were detected in the roots in T7 followed by T5 which was 1.717 mg/kg and 1.35 mg/kg, respectively. Minimum shoot length, shoot fresh weight, and shoot dry biomass were observed in T1 (control). Comparison to T1 (control) and T3, considerable increase in zinc content of shoots was recorded while comparison to T5 with T3 non-significant difference was found in shoots. While in comparison with T3 and T7 (P < .05), a significant increase was observed in T7. The Zn content present in shoot for T3, T5, and T7 was 1.327, 1.220, and 1.55 mg/kg, respectively. Similar pattern was observed for Zn contents in leaves (Figure 5). Concentration of zinc in roots, shoots and leaves of the chickpea plant at 45 days.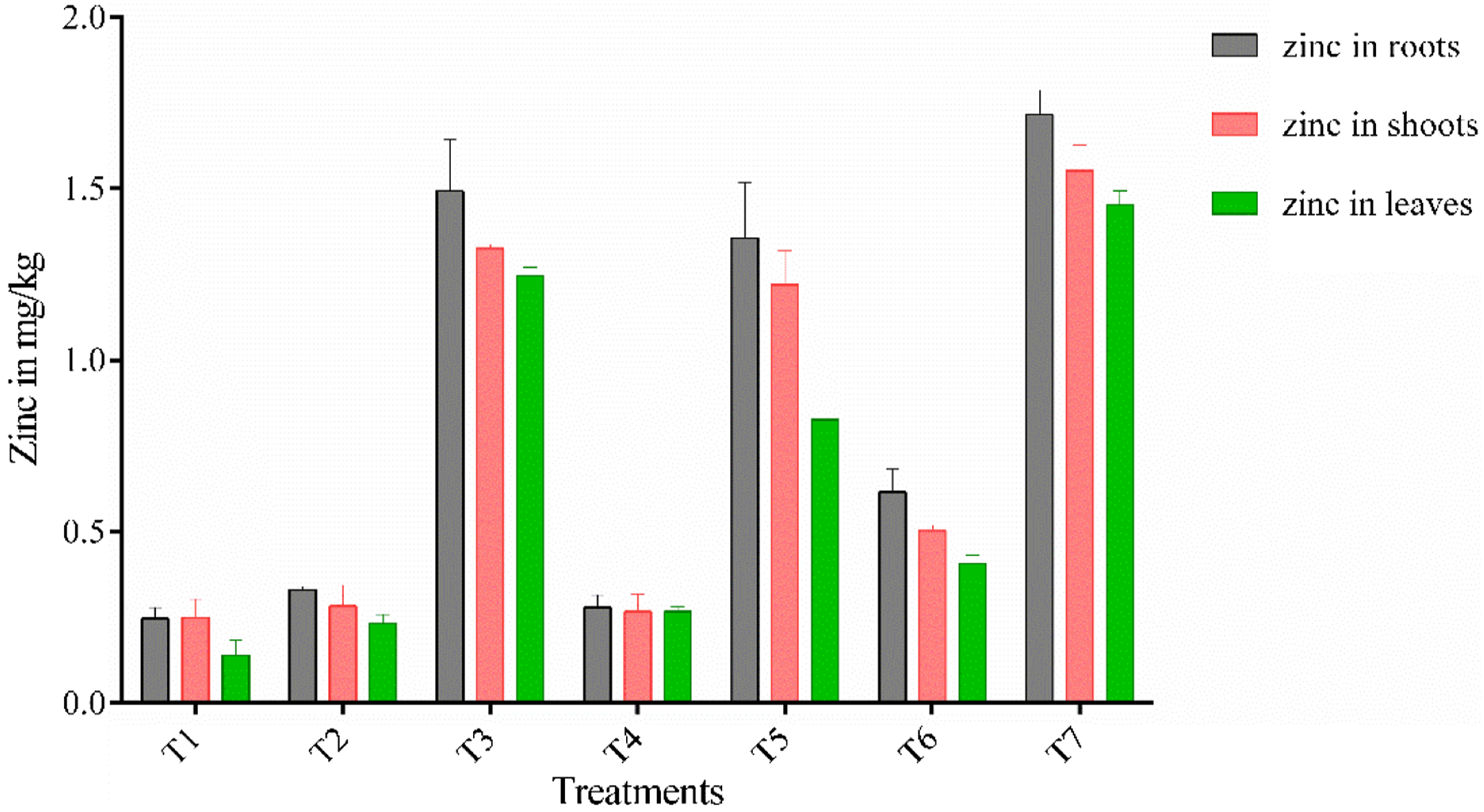
Discussion
Zinc is one of the key constituents of plants and is very crucial for their growth and development. Zinc fertilizers applied in the soil become unavailable soon after applications. Soil bacteria are reported to exert a beneficial effect on plant growth and excellent alternate to chemical fertilizers due to their zinc-solubilizing potential. These bacteria are of great interest as they have been proposed as inoculants for agriculture.19,48 In this present study, efforts were made to isolate zinc-solubilizing bacteria. These bacterial strains were screened on mineral salt medium amended with insoluble zinc salts to check their ability to solubilize zinc. Results revealed that these ten strains showed different solubilization efficiency with different insoluble zinc salts. Halo zone appearance around the bacteria colonies on solid medium was indication of solubilization potential which ranged from 15 to 23.75 mm for ZnO and 13 to 20 mm for ZnCO3. Maximum solubilization of zinc was obtained with strain RY2 and RY12 followed by RY25 and RY19. This indicates that isolated strains were able to solubilize zinc salts of which only a few isolates were able to solubilize all the 4 insoluble zinc salts. Several studies showed isolated bacterial evaluation for their zinc-solubilizing activity with various zinc sources.49,50 Variation among the bacterial strains is common when many different bacterial isolates were tested for zinc solubilization. Formation of halo zone on ZnO-amended medium using B. sp. and zinc-solubilizing efficiency (ZSE) 2.23 to 3.88 was observed in Mumtaz et al. 51 Zinc solubilization efficiency results indicated that no single strain showed uniform efficiency on all 4 zinc salts.
As the plate assay is not true indicator of zinc solubilization capacity of the bacterial strains, 52 all strains were assessed further for qualitative zinc solubilization in liquid medium supplemented with insoluble zinc salts at a concentration of .1% in order to assess soluble Zn release in the medium. Qualitative assay also confirmed the differential abilities of the bacterial strains to solubilize the zinc, just RY2 consistently showing higher zinc solubilization at all days. Maximum zinc solubilization was observed for zinc oxide followed by zinc phosphate and zinc carbonate. These findings are accordance with Saravanan et al. 48 and Sunithakumari et al. 53 who found maximum solubilization for ZnO but contrary to Ramesh et al. 54 who reported maximum solubilization for zinc phosphate. Increase in available zinc in medium was also accompanied by a reduction in pH (data not shown) that could be correlated with organic acids production such as gluconic acid, citric acid, etc. Khande et al. 55 and Dinesh et al. 29 findings support the higher zinc solubilization with lower pH due to gluconic acid production. The ability of bacteria to solubilize all the 4 inorganic zinc compounds might be interesting as zinc is known to occur in soils in discrete chemical forms varying in their solubility and availability to crop plants. 54 With the advantage of zinc solubilization to make micronutrients availability to plants, plant growth promoting characterization was also necessary to confirm plant growth promotion. For siderophore production, 8 out of 10 strains were positive. As concerned to P solubilization, almost all strains were found positive. Bechtaoui et al. 56 showed that the application of plants with insoluble phosphate and PGP bacterial strain improved the wheat and faba beans growth. Abou-el-Seoud and Abdel-Megeed 57 improved the wheat growth with the application of P and K solubilizing B. sp. Some strains showed IAA production at different level, while all strains were not able to solubilize silicate. In contrast, Kang et al. 58 found Burkholderia eburnea CS4-2 as Si solubilizer with IAA production that enhanced the rice plant growth.
On the basis of solubilization and PGP characteristics, selected strain RY2 was identified as P. protegens by molecular characterization. Bacillus, Enterobactor, and Pseudomonas genera are reported for the growth production of different crops like wheat, chickpea, mung, etc.59,60 In literature, Pseudomonas spp. such as Pseudomonas fragi (EPS1), Pseudomonas fluorescens, and Pseudomonas aeruginosa have been extensively reported for their potential to plant growth and crop yield.61,62 However, according to our knowledge, zinc solubilization and beneficial effects of P. protegens on plants were not found.
Further, the selected Zn solubilizing strain was assessed for their plant growth promoting ability of chickpea seedlings in axenic condition. The results showed that P. protegens RY2 significantly enhanced the chickpea growth parameters compared to control. Our results are accordance to Hussain et al. 50 and Ramesh et al. 54 that reported increase in growth parameters such as root and shoot length, root and shoot fresh and dry mass with inoculation of Zn solubilizing B. sp. (AZ6), and Bacillus aryabhattai strains. This increase in plant growth parameters could be related to ability of Zn-solubilizing bacteria to solubilize the nutrient and make them available to the plants. Nadeem et al. 63 reported that IAA and ACC deaminase activity by bacterial isolates improve the root growth and increase root surface area more suitable for nutrient uptake by decreasing ethylene level. In literature, Pseudomonas genus has been reported as plant growth promoters with zinc solubilization properties, IAA production, and phytohormone production. 62 Our findings showed that the Zn contents in root and shoot was significantly (P < .05) higher in the treatment involving Zn-solubilizing bacterium P. protegens RY2 with ZnO as zinc source compared to control. This was expected as with increase available Zn in soil could result in higher zinc in the plants. 64 This increase in zinc content in root and shoot is also supported in many studies with the use of PGPRs. PGPRs have been reported to overcome nutrient deficiency in many crops as Lefèvre et al. 65 revealed 12% more zinc translocation in wheat grains as compared to chemical zinc.
Zinc solubilization by using PGPR is relatively a newer approach and has lots of unexplored efficient zinc-solubilizing strains. This study indicates the potential of P. protegen to overcome zinc deficiency in countries like Pakistan where zinc fertilizers are under-used and not cost-effective. Zinc-solubilizing bacterium P. protegen strain RY2 used in present study exhibited beneficial effect on Zn uptake in chickpea plant and having the plant growth properties it could qualify as efficient strain to apply as bio-fertilizer under field conditions.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.