
Abstract
Gastrointestinal (GI) microbiota maintains a symbiotic relationship with the host and plays a key role in modulating many important biological processes and functions of the host, such as metabolism, inflammation, immune and stress response. It is becoming increasingly apparent that GI microbiota is susceptible to a wide range of environmental factors and insults, for examples, geographic location of birth, diet, use of antibiotics, and exposure to radiation. Alterations in GI microbiota link to various diseases, including radiation-induced disorders. In addition, GI microbiota composition could be used as a biomarker to estimate radiosusceptibility and radiation health risk in the host. In this minireview, we summarized the documented studies on radiation-induced alterations in GI microbiota and the relationship between GI microbiota and radiosusceptibility of the host, and mainly discussed the possible mechanisms underlying GI microbiota influencing the outcome of radiation response in humans and animal models. Furthermore, we proposed that GI microbiota manipulation may be used to reduce radiation injury and improve the health of the host.
Introduction
The microbiota refers to the community of all microorganisms, including bacteria, archaea, fungi, protozoans and viruses that exist at numerous sits within the host body, from skin, oral, to gastrointestinal (GI) track. 1 Humans harbor at least 100 trillion microbial cells, mainly bacteria and the number of genes of GI microbiota exceeds that of human genome by 150 times. 2 The largest number of microbes was found in the GI tract. 3 Increasing evidence demonstrated that GI microbiota involved in many biological processes in humans, from harvest and storage of energy, supply of nutrients and vitamins, to prevention against pathogenesis, promotion of immune system development and maturation, and maintenance of epithelia mucosa homeostasis. 4 -6 GI microbiota is the largest dynamic and complex ecosystems. 7 Alteration of the GI microbiota, termed as dysbiosis, linked with various diseases in humans. 3,4,8,9 It is known that GI microbiota has fundamental association with inflammatory and autoimmune diseases in intestinal tract, such as inflammatory bowel disease, 10 celiac disease, 11 and extra-intestinal diseases, such as obesity, diabetes, 12 rheumatic arthritis, cardiovascular disease, neurological and psychiatric diseases 8 and cancer. 9
Different factors could change the composition and subsequent functional status of the GI microbiota, including diet, antibiotics, geography and other exposure. 13 -15 Radiation is naturally present in the environment and encountered in daily life. Over the course of a lifetime, humans may be exposed to different types of radiation, especially for the cancer patients who receive radiotherapy. 16 Radiation could not only directly influence the exposed tissues of the body, but also cause indirect effects on distal tissues through gut-organ axes, such as gut-brain axes. 16 Although the connection between radiation and changes of GI microbiota has been widely investigated, little is known about whether and how the GI microbiota regulates the response to radiation in host. In this minireview, we summarized the literatures on radiation-induced alterations in GI microbiota, and the relationship between GI microbiota and radiosusceptibility of the host, and mainly discussed the possible mechanisms underlying GI microbiota influencing the outcome of radiation response in humans and in particular in experimental animal models.
Radiation Alters GI Microbial Composition
As early as 1950s-1960s, several studies had already shown that radiation could alter the numbers of some specific bacteria that present in feces, and different bacteria exhibited a large variation in sensitivity to radiation. 17 -20 For example, Vincent et al. identified an increase in Proteobacteria in rats received total-body irradiation (TBI) with 10.2 Gy X-rays, and a 4-fold decrease in Lactobacilli and a 1000- fold increase in Pseudomonas in rats received small intestine irradiation with 6.3 Gy. 20 These changes were maximal at 5-7 days after radiation, suggesting that radiation exposure may lead to a prolonged change in the GI microbiota. In another study, TBI (13.6 Gy) or local intestinal radiation alone (19.4 Gy) in the rat resulted in a 100-10,000-fold increase in Proteobacteria, and a slightly decreases in the normally predominant Lactobacilli. Maximal changes were observed at 3 days after irradiation. 20 In addition, radiation-induced alteration in the abundance of specific bacteria was also reported in mice, 21 dogs, 22 pigs, 23 and human. 24
It should be noticed that early studies relied on the cultivation of fecal bacteria followed by manual inspection and classification. However, most bacteria colonizing in the intestines are not readily cultivatable by current techniques. With the development of high throughput molecular approaches based on 16s rRNA sequencing and metagenomic sequencing, and expanding computational and bioinformatics support, it is feasible to characterize the entire microbial communities, including both pathogenic and commensal microbes that have not yet been cultured or otherwise detected. 25
In a pioneered study by Lam et al., high throughput molecular approaches based on 16S rRNA sequencing was applied to measure the shifts in intestinal microbiota populations in rat feces collected pre- and post-irradiation. 26 In this study, authors demonstrated that radiation exposure could increase Bacteroidales, Lactobacillaceae and Streptococcaceae, and decrease Clostridiaceae members at the family level. Soon afterward, many studies showed that radiation could disrupt the GI microbiata composition and promote dysbiosis (Table 1). For example, Kim et al. characterized the mouse gut microbiota and revealed that radiation could cause significant alteration in both abundance and diversity of microbiota with an increase in Alistipes and a decrease in Mucispirillum at the genus level. 27 Ritchie et al. found increased Lactobacillales and Clostridiales and decreased Firmicutes at the order level in male Sprague-Dawley rats received γ-ray irradiation. 28 In addition, exposure to high-linear energy transfer (LET) radiation (such as heavy particles in spaceflight and heavy ions in radiotherapy) could also induce alterations in the GI microbiota (Table 1). For instance, Casero et al. showed that in mice received 16O ion TBI, there was an increase in the amount of A. Muciniphila at the family level in GI track. 29 Meanwhile, the authors also observed the effect of different dose of 16O ion (0.1, 0.25 and 1 Gy) irradiation on GI microbiota. A significant increase in alpha diversity (i.e., the mean species diversity) and marked difference in beta diversity (viz., the microbiomes compositional heterogeneity) were observed between 10 and 30 days after radiation, regardless of dose. 29 Overall, these studies in laboratory animal models demonstrated that exposure to low-LET or high-LET irradiation could alter the composition of the GI microbiome. Generally, the most significant alterations in the GI microbiota associated with radiation exposure in mice are the increase in Lactobacillaceae and Bacteroidaceae, and the decrease in Lachnospiraceas/Ruminococcaceae at the family level (Table 1).
Experimental Studies on Radiation-Induced Alterations in GI Microbiota in Animal Models.

Moreover, the effect of radiation on GI microbiota has also been investigated in clinical studies in humans (Table 2). For example, Nam et al. observed a significant decrease in Firmicutes and an increase in Fusobacteria at the phylum level during pelvic radiotherapy. 36 Although there were discrepancies regarding the nature of these alterations between studies, all these 5 studies 35 -39 concluded that there was a significant difference in gut microbiota between cancer patients prior to and post pelvic radiotherapy, and radiation could result in a significant alteration in gut microbiota community. GI microbiota dysbiosis could be a useful biomarker to predict and prevent radiation-induced bowel toxicity. 26,35,38 Importantly, Kumagai et al. 40 and AI-Qadami et al. 41 have given reviews on the interplay between GI microbiota and radiation induced bowel injury.
Clinical Studies on Radiation-Induced Alterations in GI Microbiota in Cancer Patients and Healthy Control.

GI Microbiota Influences the Radiosusceptibility of the Host
In contrast to the intensive studies about the radiation-induced effects on GI microbiota, relatively little is known about whether and how the GI microbiota modulates the response of the host to radiation. The role of GI microbiota in modulating radiosusceptibility of the host is of great concern, and several studies in mouse models has linked the GI microbiota to radioresistance. Back in 1964, McLaughlin et al. observed that germ-free (GF) mice were more radioresistant than either E.coli-monocolonized or conventional mice. 18 In another study, Crawford and Gordon also observed that GF mice were markedly resistant to lethal radiation enteritis. 42 Where, GF mice had fewer apoptotic endothelial cells in the intestinal mucosa and less lymphocyte infiltration than conventionally raised mice 4 h after TBI with a higher dose (16 Gy γ-rays from a 137Cs source). The conventionally raised mice still exhibited earlier death than GF mice even though hematopoietic rescue performed by bone marrow transplantation. The resistance of GF mice to the enteric toxicity of TBI could be due to the absence of gut microbiota that existed in the conventionally raised animals. 42 Moreover, the role of the microbiota in imparting this increased radiosusceptiblity was confirmed by colonizing GF mice with gut microbiota harvested from conventionally raised mice. 43
Recently, Cui et al. investigated the effect of gut microbiota on radiosusceptibility using 16s rRNA sequencing. 44 When mice were exposed to TBI of 5 Gy γ-rays, it was found that mice with normal of the circadian rhythm (12 h dark/12 h light cycle) had a significantly better survival than mice with disruption of the circadian rhythm (8 h dark/16 h light or 16 h light/8 h dark cycle). The mice housed in aberrant light cycles harbored a reduction of intestinal bacterial species and shifts of gut bacterial composition compared with those kept under normal light cycles. These findings suggested that circadian rhythm could shape the gut microbiota which could affect the radiosusceptibility of the host. 44 In another study by Cui et al., they described the association between gut microbiota community and radiosusceptibility on an antibiotic-treated mouse model. 45 They found that the enteric bacterial composition of treated mice was significantly different from that of control group, and the survival rate of antibiotic-treated mice was significantly higher than untreated mice after irradiation. These observations indicated that changes in gut microbiota community were able to influence the radiosusceptibility of host mice. In addition, Maier et al. reported that mice with restricted microbial composition (RM) were more susceptible to high-energy protons radiation-induced DNA damage than the conventionally raised mice. Their work indicated a dysbiotic intestinal microbiota could regulate the radiosusceptibility of RM mice. 46,47 Furthermore, several clinical studies in humans also implied that the initial GI microbiota colonization before pelvic radiotherapy could impact radiosensitivity of the normal tissues. 35,37,39
Once colonized in the intestine, microbiota influences gut barrier functions and host stress response through metabolic activities such as short chain fatty acids (SCFAs) from indigestible carbohydrates of the host and tryptophan metabolites from amino acid metabolism. 48,49 It has been demonstrated that several microbial metabolites involved in the response to radiation of the host. For example, Vernia et al 50 and Hille et al 51 reported that butyrate, a SCFA, has protective effect against radiation-induced proctitis owing to reducing of radiosusceptibility, leading to the mitigation of inflammation. Recently, Li et al demonstrated that gut microbiota produced-valeric acid (VA) exerted the most significant radioprotection among the SCFAs. Oral replenishment of VA significantly attenuated the response of the host intestine, improved GI tract function and intestinal epithelial integrity, and elevated the survival rate of irradiated mice. 52 Besides SCFAs, another important molecules derived from microbial metabolites are some amino acids, such as cysteine and tryptophan. Cysteine can be converted into glutathione, which can help to protect the host from reactive oxygen species (ROS). Indeed, treatment with N-acetyl cysteine (NAC) could ameliorate radiation-induced oral mucosa damage. 53 And indole 3-propionic acid, a bacterial-mediated production from tryptophan, protected mice against radiation toxicity. 54 The studies demonstrated gut microbiota involved in the responses to radiation in host and influence the susceptibility to radiation exposure directly through their metabolites.
Together, these works mentioned above provided solid evidence that the GI microbiota and microbiota-produced metabolites could influence the radiosusceptibility of the host. Some species of the GI microbiota at right abundance could influence the outcome of radiation responses in the host at local tissues and even at a systemic level.
Possible Mechanisms Underlying GI Microbiota Influencing Radiosusceptibility of the Host
Involvement of the microbiota in the integrity of the mucosal barrier
GI microbiota plays a critical role in maintaining intestinal homoeostasis, and has some protective effect on intestinal integrity. In physiological conditions, the commensal bacteria in the intestinal lumen are prevented from translocating through the intestinal mucosa by an intact epithelial barrier, which covered by a mucus layer poorly permeable to microorganisms. 55 After exposure to radiation, both gut microbial cells and host cells (i.e., intestinal epithelial cells, IECs) are influenced by direct and indirect actions of radiation. On one hand, radiation can induce cell apoptosis in crypt and villous atrophy, resulting in an increase in intestinal permeability. On the other hand, as summarized in the previous section, radiation can change the GI microbiota compositions, resulting in dysbiosis, which aggravated radiation-caused intestinal damage. 55,56
It showed more than a decade ago that the small bowel of GF mice was markedly resistant to lethal radiation enteritis, 42 due to the decreased apoptosis of endothelial cells and lymphocytes in the mesenchymal cores of the small intestinal villi in GF mice compared to that in the conventional mice. Mechanistically, fasting-induced adipose factor (Fiaf) expression was selectively and significantly downregulated by the GI microbiota in the gut epithelium. Fiaf -/- mice are susceptible to radiation-induced lethal enteritis. 42 While Fiaf has pleiotropic effects on lipid metabolism, angiogenesis, tissue repair and restoring integrity of the mucosal barrier. 57 Although some intestinal bacteria, such as Bifidobacterium, Lactobacillus and Streptococcus, could induce Fiaf expression in IECs and mediate protection for mucositis and colitis. 58 -60 However, due to the potential influence of radiation-induced dysbiosis, these bacteria may be altered, and the protective effect of Fiaf expression can be greatly diminished.
In addition, a large pilot study by Ferreira et al. showed that there was a significant increase in Phascolarctobacterium, Roseburia, Propionibacterium, and Clostridium IV at the genus level in patients suffering from radiation enteropathy which was related to the poor intestinal barrier function. 39 Similarly, Wang et al. also showed an elevated Proteobacteria, Gammaproteobacteria (primarily Enterobacteriaceae), and a decreased Bacteroides at the family level in patients with radiation-induced diarrhea that was mainly due to mucosal integrity damage. 37 Furthermore, GI microbiota also regulates intestinal barrier function by modulating the expression and distribution of tight junction proteins. 61 These studies indicated that the microbiota disruption after radiation exposure is responsible for the alterations of intestinal permeability, and at least, partly involve in the integrity of the intestinal mucosal barrier (Figure 1).
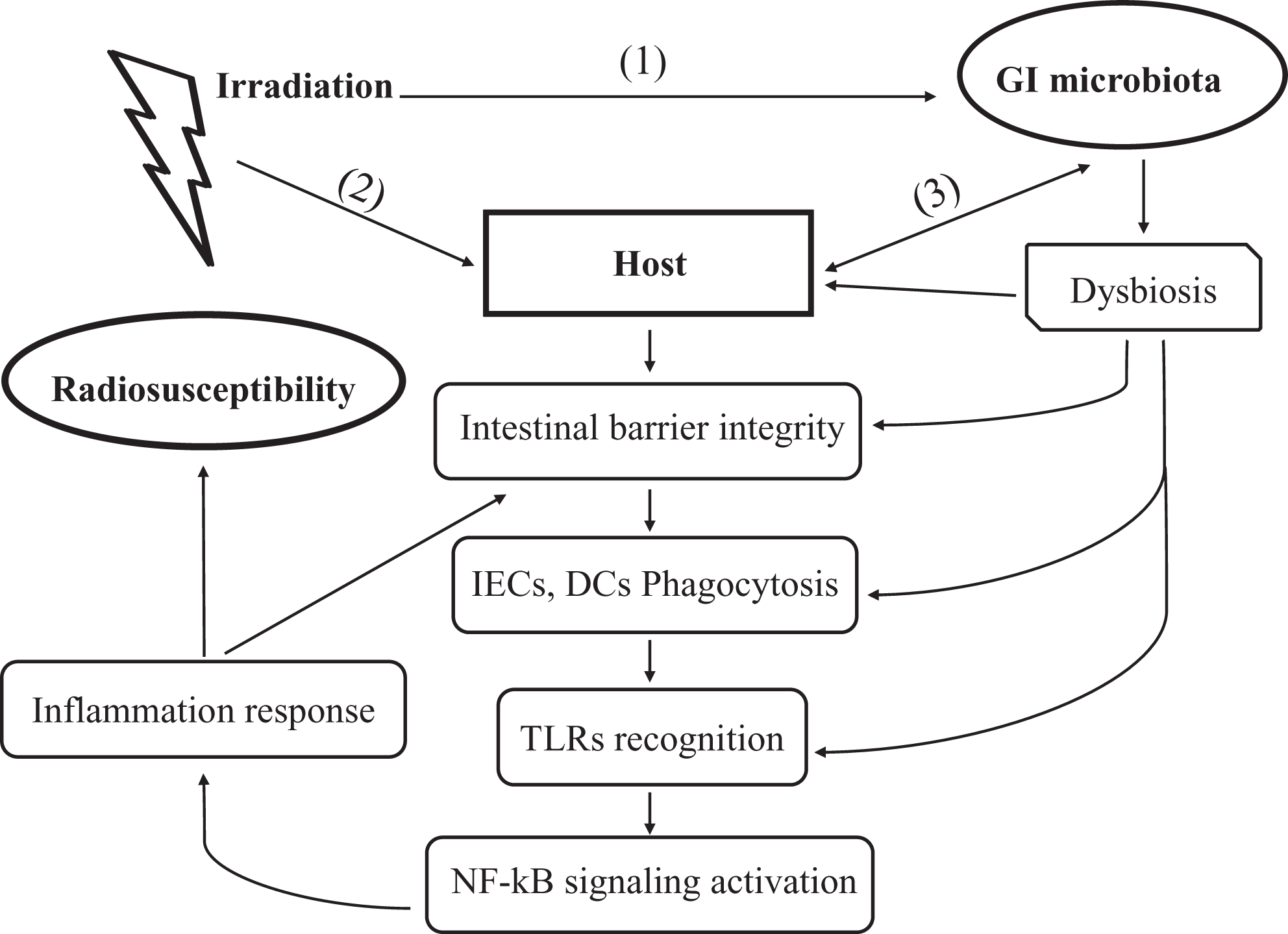
GI microbiota influences radiosusceptibility of the host through modulating intestinal immune reaction. (1) Irradiation alters the GI microbiota composition and causes gut microbiota translocation and dysbiosis that consequently disrupt intestinal homeostasis, and induce release of inflammatory cytokines; (2) irradiation causes cell loss and damage to the host that lead to increased intestinal permeability. These changes further trigger response of IECs and innate immune cells (such as macrophages and DCs) to radiation-induced “danger signals” from microbe- or damage-associated molecular patterns (MAMPs/DAMPs) by releasing inflammatory cytokines and activating NF-κB signaling; (3) bidirectional interaction between GI microbiota and the host. The GI microbiota in normal circumstance could balance between anti-inflammatory and pro-inflammatory signals to prevent bacterial dissemination and avoid host tissue damage; alterations in GI microbiota could exert directly pro-inflammatory effects on the epithelial cells, damage intestinal barriers and enhance radiosensitivity of host gut track. Collectively, effect of radiation on the host is a combined outcome of the complex interactions among GI microbiota, the host, and irradiation. Of note, microbiota could influence radiosusceptibility of the host through modulating intestinal immune reaction. IECs: intestinal epithelial cells; DCs: dendritic cells; NF-κB: nuclear factor κB.
Microbiota as a cause of inflammation
Increasing evidence links radiation effects with inflammation-related mechanisms 62,63 and the relationship between GI microbiota and inflammation has been well elucidated. 64 While there is limited empirical evidence about the effects from radiation-induced alterations in GI microbiota composition on the inflammation response of the host. Recently, Gerassy-Vainberg et al. demonstrated that GI microbiota exerts directly pro-inflammatory effects on the epithelial cells and increases the intestinal permeability. In their study, localized internal rectal irradiation was used to investigate the effects from radiation-induced dysbiosis on mouse proctitis. 30 When human HT29 colonic epithelial cells were co-cultured with fecal bacteria from the mice with radiation proctitis, expression of pro-inflammatory cytokines such as TNF-α and IL-1β was upregulated. In contrast, fecal bacteria from naïve control mice were unable to elevate expression of pro-inflammatory cytokines. In addition, GF mice that received fecal bacteria from mice with radiation proctitis showed a more intense inflammatory reaction than that from naïve control mice. 30 This work for the first time linked the alteration of GI microbiota caused by radiation to the inflammation in the GI tract. They posited that, compared to those with non-irradiated microbiota, radiation induced GI microbiota dysbiosis could exert directly pro-inflammatory effects on the epithelial cells, increasing their expression of pro-inflammatory cytokines such as TNF-α and IL-1β. 30 Furthermore, the pro-inflammatory cytokines such as TNFα and IL-1β could also impair tight junction barrier and increase the intestinal permeability. 65,66 On the one hand, the disruption of intestinal epithelial permeability could greatly increase the chance of gut bacterial translocation and pathogen invasion. On the other hand, the translocation and invasion of bacteria, in turn, would further aggravate loss of the radiosensitive cells and intensify the outcome of a “leaky” mucosa, expose mucosa immune cells to luminal antigens and amplify the inflammatory cascades via pattern-recognition receptors (PRRs) signaling. 67 (Figure 1). Besides, it seems that radiation is more detrimental to the commensals than to the pathogens, leading to a further dysbiosis state. 68 Microbes that are capable of surviving from radiation exposure overgrow to dominant bacterial community. 69 The dislocated and entero-invasive microbiota could directly induce the production of ROS and chemokines in IECs, further exacerbating the inflammatory dysbiosis. 55
Activation of toll-like receptors (TLRs)
TLRs are a class of PRRs that are expressed on epithelial cells and immune cells such as macrophages and dendritic cells (DCs). Recognition of intestinal commensal bacteria or their products by TLRs is necessary for the regulation of intestinal homeostasis. 70 TLRs recognize microbial components and induce the innate immune response against pathogens. GI microbiota might control the TLRs by secreting these natural ligands to activate the innate immune response of the host in radiation response. For example, one study showed that injection of a polypeptide derivative of flagellin, a TLR5 ligand, before exposure to lethal doses of radiation, could enhance proliferative response of the intestinal crypts cells and decrease mortality in mice and non-human primates. 71 Another study demonstrated that pretreatment with TLR9 agonist (CpG DNA, typically found in viruses) attenuated radiation-induced intestinal injury and improved the survival of mice. Additionally, administration of lipopolysaccharide (LPS) (the composition of outer membrane of Gram-negative bacteria), a TLR4 ligand, to mice before radiation, could protect IECs against radiation-induced apoptosis and increase the survival of crypts through induction of COX-2 and the production of prostaglandins. 72 LPS could active TLR4-expressing cells, leading to the release of TNF-α and production of prostaglandins that could decrease radiation-induced epithelial stem cells apoptosis. 73 Conversely, injection with polyinosinic: polycytidylic acid (Poly I:C), a TLR3 agonist, which is naturally found in viruses, aggravated radiation-induced GI symptoms in a TLR3-dependent manner. 74 The activation of TLR3 appears to exacerbate radiation injury to the bowel. It is well known that bacterial flagellin, LPS, CpG DNA, and Poly I: C, are all derived from GI microbiota, thus it is possible that the GI microbiota themselves could autonomously control the TLRs by secreting these natural ligands to activate the innate immune response of host after radiation exposure. The potential mechanisms by which TLR involves in radiation response was associated with the subsequently activation of the nuclear factor-kappa B (NF-kB) pathway (Figure 1). 75,76
Initiating immune reaction
The GI microbiota plays an essential role in secondary and tertiary lymphoid structure. Notably, the interaction of gut microbiota and the host IECs is critical in the maturation and maintenance of the mucosal immune function. 77 GF mice lack of microbiota have smaller Peyer’s patches and a reduced number of CD4+ T cells and IgA-producing plasma cells. 78 The presence of specific bacteria in the gut microbiota is directly correlated with T cell development and differentiation. 41 In the normal state, GI microbiota and its components (e.g., LPS) or metabolites (e.g., short chain fatty acids, SCFAs) actively dampen host immune response by inhibiting NF-κB inflammatory pathway or certain inflammasome, promoting the conversion of CD4+ T cells into Foxp3+ regulatory T cells (Tregs) and increasing the production of anti-inflammatory cytokines such as Interleukin-10 (IL-10). 55 The GI microbiota is a key orchestrator of intestinal homeostasis by a balance between anti-inflammatory and pro-inflammatory signals that prevent bacterial dissemination while avoiding host tissue damage. 79 However, the intestinal homeostasis is lost when exposure to radiation. Besides reacting to the damage signals directly induced by radiation, the host immune system is vigorously responding to pathogens invasion. First of all, NF-κB signaling and inflammasome pathways are activated both in IECs and innate immune cells at the elevated level of ROS, initiating immune response and amplifying radiation effect. 64,80,81 Especially, innate immune cells such as macrophages, neutrophils and DCs, sense and respond to the radiation-induced “danger signals” from microbe- or damage-associated molecular patterns (MAMPs/DAMPs), leading to release of inflammatory cytokines (TNFα, IL-1β, and IL-6), and hence initiating inflammatory response. 67,80,82 The activation of inflammatory responses prolongs the radiation response by generating more ROS, cytokines and growth factors. 55 In addition, expression or release of these MAMPs/DAMPs leads to the activation of antigen-presenting cells (APCs), particularly DCs. The activated APCs engulf these damaged, death cells or pathogens, process and present them to cytotoxic T lymphocytes and subsequently induce immune response. Despite a large number of evidences focusing on the impact of radiation on immune cells, there is limited empirical evidence on the combined effects from radiation and microbes on the host immune activation. The mucosal barrier, inflammation and Toll-like receptors are all important links in the influence of GI microbiota on immune response after radiation (Figure 1). Given the role of GI microbiota in modulating immune cells in the response to radiation, these studies also indicated that GI microbiota could fulfill a role in the response to radiation by initiating immune reaction.
The microbiota lives in GI tract and is closely interacting with the host IECs, when exposure to radiation, both the GI microbial cells and host IECs are influenced by direct and indirect actions of radiation. Therefore, the effect of radiation on host is a combined outcome of the stress responses from the gut microbes, host IECs and immune cells with multiple interactions among each other (Figure 1). Although whether and how the GI microbiota influence the response to radiation of the host is still largely unknown, cumulative evidence indicates that the mechanisms underlying GI microbiota influencing radiosusceptibility are complex, with dynamic changes in microbial communities interacting with various components of enteric systems.
Interestingly, it has been found that gut microbiota contributed to cancer treatment through modulating host intestinal and/or system immune reaction. Though the host immune response influences both tumor response to radiation (efficacy) and normal gut tissues response to radiation (toxicity), the mechanisms underlying microbiota modulating the efficacy and toxicity of radiation appear to be different. Gut microbiota potentially impacts the efficacy of radiotherapy through anti-tumor immune response, which is mediated by Th17 and CD8+ cells. While gut microbiota influence the radiation-induced toxicity via translocation and dysbiosis, which disrupt intestinal immune homeostasis and release of pro-inflammatory cytokines. 41 This revealed that gut microbiota functions with different biological effects in different microenvironment, even in the same radiation exposure. Therefore, an in-depth understanding of the mechanisms requires a careful and systematical parsing of the multivariate processes.
GI Microbiota Manipulation in the Exposure to Radiation
GI microbiota interacts with host IECs and/or innate immune cells in a bidirectional manner. Radiation disrupts GI microbiota composition and promotes dysbiosis, which, in turn, affects radiation response of host. Thus, application of GI microbiome manipulation to reduce and/or prevent radiation-induced systemic toxicity and improve health of host is of great concern. Several strategies have been used in preclinical or clinical experiments to test the effect of GI microbiota on radiation response. For example, administration of antibiotics is part of the standard treatment plan in the event of acute radiation syndrome. 83,84 Yet, some intestinal commensal bacterial species provide protective functions in the response to radiation. 85 Hence, it is preferable that administration of the antibiotics could uniquely target bacterial organisms with a proclivity to translocate after radiation, while spare bacterial species that are benefit in mitigating the dysbiosis and detrimental effect. In addition, administration of probiotics (such as Lactobacillus and Bifidobacterium) and/or prebiotics (such as SCFAs) is another strategy for improving health of the host and reducing radiation toxicity. Both pre- and probiotics showed beneficial effects on the GI tract, including antimicrobial peptide production, increased mucus production by Goblet cells, SCFA production, antioxidant effects, barrier enhancement and prevention of pathogenic biofilm formation. 86 -89 However, there are discrepancies regarding the benefit functions of pre- and probiotics for reducing the radiation-caused intestinal toxicity, 89 -93 which suggests that future research should focus on pairing the GI toxicities with certain microbial phenotypes to allow targeted microbiota manipulation.
Fecal microbiota transplant (FMT) is also considered as an effective method to manipulate the GI microbiota. FMT was first used in fourth-century China to treat an assortment of GI symptoms, now is primarily treated recurrent Clostridium difficile infection. 94 Recently, Cui et al. demonstrated that FMT could alleviate radiation-induced toxicity in mice. They found that the GI microbiota of male and female mice was different, and the difference was associated with the susceptibility to radiation-induced toxicity. When FMT from sex-matched healthy donor stool was performed immediately after radiation, which increased the survival rate of FMT-responded mice. 45 In humans, a pilot study of FMT for chronic radiation enteritis (CRE) was performed by Ding et al. Five female patients underwent FMT, 3 patients responded to FMT and experienced improvement in diarrhea, rectal hemorrhage, abdominal/rectal pain and fecal incontinence and Karnofsky Performance Status score. 95 Although the preclinical and clinical relevance of FMT seems promising, FMT still come with a lot of unknowns and uncertainties. It still should be cautious regarding FMT trials or experiments and looking forward standardized approaches.
Conclusions
An increasing number of studies showed that GI microbiome is significantly altered by radiation. Meanwhile, accumulative evidence highlighted the key role of the GI microbiome in influencing the outcome of host radiation response. Given the role of GI microbiota in modulating immune cells known to involve in the response to radiation, it is suggested that GI microbiota involve in the response to radiation at least partly through modulating the host immune reaction (Figure 1). Although the underlying mechanisms are still to be well identified, with the development of promising tools to optimize prevention and therapeutic strategies, GI microbiota composition is expected to be used as a biomarker to estimate radiosusceptibility and radiation health risk in the host.
At present, our knowledge of GI microbiota is restricted to bacterial information owing to technical limitations, while future advancements will enable us a better and comprehensive understanding of the GI microbiota, the complex interactions among GI microbiota, the host, and irradiation. The ultimate goal would be to identify the most favorable composition of GI microbiota and its productions and/or metabolites, and develop the preventive and therapeutic strategies through such as modulation and manipulation of the microbiota to reduce radiation-induced health risk and improve the health of the host.
Footnotes
Acknowledgments
The authors thank Ms. Mikiko Nakajima, Ms. Hiromi Arai, and Ms. Yasuko Morimoto (QST-NIRS, Japan), for their expert technical assistance and administrative support. Thanks, are also due to the anonymous peer reviewers for providing the constructive comments that strengthened the presentation of this work.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interests with respect to the research, authorship, and publication of this paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article. The first author was a fellow of The Japan-China Sasakawa Medical Fellowship (fiscal year 2020) from The Japan China Medical Association supported by The Nippon Foundation, Japan.