
Abstract
Adenosine triphosphate (ATP) serves as a signaling molecule for adaptive responses to a variety of cytotoxic agents and plays an important role in mediating the radiation stress-induced responses that serve to mitigate or repair the injurious effects of γ radiation on the body. Indeed, low doses of radiation may have a net beneficial effect by activating a variety of protective mechanisms, including antitumor immune responses. On the other hand, ATP signaling may be involved in the radiation resistance of cancer cells. Here, focusing on our previous work, we review the evidence that low-dose γ irradiation (0.25-0.5 Gy) induces release of extracellular ATP, and that the released ATP mediates multiple radiation-induced responses, including increased intracellular antioxidant synthesis, cell-mediated immune responses, induction of DNA damage repair systems, and differentiation of regulatory T cells.
Keywords
Induction of Antioxidant Synthesis and Immune Function by Low Doses of Ionizing Radiation
It is well known that induction of endogenous antioxidants in organisms is one of the adaptive responses to various kinds of cytotoxic agents, including reactive oxygen species (ROS), quinone-based anticancer agents, metals, ultraviolet, and nanoparticles, as well as ionizing radiation. 1 -8 Many reports have described the responses to ionizing radiation, 9 -12 including our studies on the effects of low-dose γ rays on the enhancement of antioxidant potency in mouse and cultured cells. 13 -15
We examined induction of endogenous antioxidants in mice exposed once over the whole body to γ rays. Total intracellular reduced form of glutathione (GSH) in splenocytes was significantly elevated from 2 to 6 hours after 0.5 Gy postirradiation, reaching a maximum at around 4 hours and declining to the 0-time level within 24 hours. Other antioxidants, such as glutathione peroxidase (GPX), superoxide dismutase (SOD), catalase (CAT), and thioredoxin (TRX), were also similarly induced, 13 as was expression of the corresponding genes. 14 The induction of these antioxidants is considered as one of the adaptive responses against ROS, since low-dose radiation induces ROS formation, for example, through radiolysis of water molecules.
We also examined natural killer (NK) activity, which is an index of cell-mediated immunity. The activity in mouse splenocytes after whole-body single γ irradiation at a dose of 0.5 Gy was also increased significantly from 4 to 6 hours and then declined to the nonirradiation control level by 24 hours. 16
Third, we examined the effect of exogenous GSH and precursors of de novo GSH synthesis, such as cysteine (Cys), N-acetylcysteine (NAC), glycine, and glutamine, on NK activity, using splenocytes obtained from nonirradiated normal mice, to see whether GSH induced by low-dose γ irradiation was involved in the enhancement of the immune function.17 The NK activity was elevated dose dependently at GSH concentrations ranging from 0 to 5 mM. Exogenous GSH increased the cellular GSH level in a similar way. Other precursors such as Cys and NAC elevated NK activity more effectively than GSH. Furthermore, we investigated the effect of buthionine sulfoximine (BSO), a specific inhibitor of γ-glutamylcysteinyl synthetase, which is the rate-limiting enzyme of the de novo GSH synthetic pathway, on the elevated NK activity induced by the precursors. The elevation was blocked by BSO, and the extent of inhibition was in good agreement with the changes of cellular total GSH content. 15,17 These results indicated that the de novo pathway of GSH synthesis is involved in modulating NK activity in mice irradiated with low-dose γ rays.
Adenosine Triphosphate Signaling
Intracellular adenosine triphosphate (ATP) is released into the extracellular space in response to various stimuli, including shear stress, stretch, hypoxia, inflammation, osmotic swelling, pH, ionizing and nonionizing radiations, ROS, nanoparticles, and cell death, followed by activation of purine 2 (P2) receptors. 7,18 -20 The P2 receptors are classified into 2 major subtypes, that is ionotropic P2X receptors and metabotropic P2Y receptors, and various physiological functions are modulated through activation of these receptors (Figure 1). 21 The released ATP is rapidly metabolized to adenosine diphosphate, adenosine monophosphate, and adenosine by ectonucleotidases. 22 These metabolites bind with P2X, P2Y, and/or P1 receptors with different affinities.

Schematic overview of purinergic/ATP signaling. Intracellular ATP is released into the extracellular space in response to various stimuli. The released ATP is rapidly metabolized to ADP, AMP, and adenosine, and these metabolites bind with P2X, P2Y, and/or P1 receptors, respectively, and activate receptors with a different concentration, leading to downstream signaling. ATP indicates adenosine triphosphate; ADP, adenosine diphosphate; AMP, adenosine monophosphate; P2, purine 2.
Various ATP release pathways have been reported, including a maxi-anion channel, P2X7 receptor/pore, a volume-sensitive outwardly rectifying chloride channel, a member of the ATP-binding cassette protein family, a gap junction hemichannel, and vesicular exocytosis (Figure 2). 19,23 -30 The ATP released from cells through these pathways activates P2 receptors in an autocrine or a paracrine manner, 18 leading to activation of downstream purinergic signaling/ATP signaling pathways. Outcomes include activation of DNA repair, induction of antioxidants, apoptotic cell death, and so on. 31 -34 Though the latter phenomena are also observed in cells exposed to γ rays, involvement of ATP signaling in the radiation-induced effects was not clear.

Pathways/channels of ATP release from cells. The pathways include a maxi-anion channel, P2X7 receptor/pore, a volume-sensitive outwardly rectifying chloride channel, a member of the ATP-binding cassette protein family, a gap junction hemichannel, and vesicular exocytosis. ATP indicates adenosine triphosphate; P2, purine 2.
ATP Release Through P2X7 Receptor From Cells Exposed to γ Rays
Therefore, we investigated whether radiation-induced release of ATP contributes to induction of the cellular antioxidant systems. 35,36 First, dose-dependent release of ATP was investigated in B16 melanoma cells exposed to γ rays at doses ranging from 0 to 8 Gy. The ATP release was greatest at the dose of 0.5 Gy. Doses of more than 1.0 Gy resulted in lower ATP release, suggesting that oxidative damage to membrane proteins at higher doses of γ irradiation might cause impairment of ATP-release mechanisms. It is noteworthy that the dose of 0.5 Gy that gives the greatest ATP release is in accordance well with that we used in animal experiment so far. 13,14,16,37 The concentration of ATP in culture medium of irradiated cells exposed to 0.5 Gy reached a peak at 5 minutes and then declined to the basal level within 30 minutes. We examined which pathways might be involved in the ATP release using various inhibitors of the maxi-anion channel, volume-sensitive outwardly rectifying chloride channel, P2X7 receptor channel, and so on. Glibenclamide, flufenamic acid (FFA), GdCl3, and arachidonic acid, which are inhibitors of anion channels or transporters, did not block radiation-induced ATP release, but the release was significantly inhibited by a highly selective antagonist of P2X7 receptor, A438079, 38 suggesting involvement of P2X7 receptor in radiation-induced ATP release in B16 melanoma cells. 38 This was confirmed by means of studies with P2X7 receptor-knockdown B16 melanoma cells. These results strongly support the involvement of P2X7 receptor in ionizing irradiation-induced ATP release in the melanoma cells used in our study. However, it should be noted that the contributions of ATP-releasing pathways may vary depending upon the type of cell examined.
Induction of TRX-1 by γ Ray Irradiation and Exogenously Added ATP
The role of thioredoxin-1 (TRX-1) in the induction of endogenous antioxidants by γ irradiation was examined in detail. The change of TRX-1 expression was evaluated in mouse macrophage-like RAW264.7 cells exposed to 0.5 Gy γ rays by means of immunoblotting. The TRX-1 expression increased time dependently from 1 hour after irradiation, peaking between 3 and 6 hours, and remained elevated until 24 hours after irradiation. The increase was almost completely blocked by pretreatment with the ectonucleotidase apyrase, suggesting that the γ ray irradiation-induced ATP release is involved in mediating the increase of TRX-1 expression. Therefore, we examined the effect of ATP itself on TRX-1 expression. Exogenously added ATP (100 μM) also increased TRX-1 expression time dependently, with a maximum at approximately 6 hours posttreatment, and the extent of the increase was dose dependent. Again, the increase was suppressed by pretreatment with apyrase. Taken together, these results indicate that extracellular ATP and activation of purinergic signaling are involved in the elevation of TRX-1 induced by γ irradiation. 39
Next, selective purinergic-receptor antagonists were used to identify the specific pathway(s) through which extracellular ATP induces increased TRX-1-expression of RAW264.7 cells. The induction of TRX-1 and increase of cellular ATP by γ rays were both blocked by pretreatment with apyrase, P2X7 receptor-specific antagonist A438079 and P2Y6 receptor-specific antagonist MRS2578, suggesting the involvement of both P2Y6 and P2X7 receptors in TRX induction.
Overall, these results indicate that ATP released from cells exposed to γ rays mediates activation of cellular antioxidant systems as a defense against newly released intracellular ROS, including those generated by direct action of ionizing radiation on water molecules (Figure 3).

ROS production in RAW264.7 cells via ATP signaling. ATP is released from the irradiated cells and subsequently may produce ROS by the activation of cell membrane nicotiamide adenine dinucleotide (NADPH) oxidase via purinergic signaling. Antioxidants, such as TRX-1, SOD, GSH and so on, are thought to be induced as an adaptive response newly released intracellular ROS, including those generated by direct action of ionizing radiation on water molecules. ATP indicates adenosine triphosphate; NADPH, TRX-1, thioredoxin 1; SOD, superoxide dismutase; GSH, glutathione; ROS, reactive oxygen species.
Suppression of Solid Tumor Growth by Low-Dose Whole-Body γ Irradiation and Its Mechanism
It is now known that low-dose irradiation can have beneficial effects in living organisms, including increase of endogenous antioxidants and cell-mediated immunity, activation of DNA repair, and inhibitory effects on inflammation. 17,40,41 In addition, low-dose ionizing radiation suppresses tumor growth and metastasis both clinically and in animal studies. 12,42 -51 In these articles, it is suggested that the antitumor effect is due to activation of the immune system, especially elevation of cytotoxic immune responses. We have examined this issue. First, we confirmed the inhibitory effect of low-dose whole-body γ irradiation on solid tumor growth in Ehrlich tumor-bearing mice. Male Institute of Cancer Research (ICR) mice were exposed to a whole-body γ irradiation dose of 0.5 Gy. At 6 hours after the irradiation, Ehrlich tumor cells (2 × 105 cells) were subcutaneously implanted into the right leg. The mice were irradiated 3 times more (0.5 Gy/each) at 4-day intervals (total dose: 2.0 Gy). The tumor size was measured from day 9 through day 30. Gamma irradiation suppressed solid tumor growth at each time point. 37
Next, we examined the mechanisms involved. The NK cells, cytotoxic T lymphocytes (CTLs), and macrophages are essential immune cells for cell-mediated antitumor immunity. It has been reported that NK cells were responsible for the suppression of pulmonary metastasis by X-ray irradiation at 0.1 to 0.2 Gy. 52,53 Also, we had previously shown that γ irradiation increased the amount of antioxidants such as GSH in tissues and cultured cells, leading to upregulation of cell-mediated antitumor immunity. 16,17 These results suggest that elevated levels of antioxidants in immune cells of irradiated mice might be involved in the enhancement of cell-mediated antitumor immunity, and it seemed plausible that irradiation would activate a signaling cascade involved in the T-cell differentiation processes, such as interleukin 12 (IL-12) production by macrophages as antigen-presenting cells (APCs), thereby promoting a T helper 1 (Th1)-dominant state. Thus, Th1- and Th2-type cytokines were assayed to examine the Th1/Th2 balance. Tumor-bearing mice were irradiated with a single dose of 0.5 Gy γ rays, and splenocytes were isolated 48 hours later. Interferon-γ (IFN-γ) and IL-4 (Th1 and Th2 cytokines, respectively) were assayed in irradiated and control mice. The splenocytes from the irradiated mice produced more IFN-γ than those from control mice, whereas little difference was found in IL-4 production between the 2 groups of mice; consequently, the IFN-γ/IL-4 ratio was significantly elevated in splenocytes of the irradiated group, suggesting that the balance of immunity was skewed toward Th1. To confirm this, we measured production of IL-12 in macrophages isolated from the abdomen of mice at 48 hours postirradiation with γ rays (0.5 Gy). Indeed, IL-12 production by these macrophages was increased. The irradiation also increased the cellular GSH level of the macrophages. The increase in the GSH level was sustained for a few days after irradiation. Thus, elevation of GSH in macrophages may also be involved in the shift of the immunity balance. It is noteworthy that NK and CTL activity were both reportedly enhanced with increase of intracellular GSH content. 54 Further, in order to see whether the enhanced IL-12 production and elevated GSH level are due to a direct effect of irradiation on macrophages, we isolated macrophages from nonirradiated mice and irradiated them in vitro. No significant differences were seen between the irradiated and nonirradiated groups, suggesting that the enhanced production of IL-12 and the elevated GSH level of macrophages are induced indirectly, not by a direct effect of irradiation. As mentioned elsewhere, the intracellular GSH level in APC (including macrophages) has been found to play a critical role in the determination of immunity; a higher level of GSH favors Th1 55 -57 by increasing IL-12 production. Though elevation of IFN-γ was confirmed in γ-irradiated mice, IFN-γ alone cannot stimulate macrophages to produce IL-12. However, production of IL-12 by macrophages is required for stimulation with lipopolysaccharide. 58 -60 The IFN-γ may potentiate macrophages to produce more IL-12 by increasing GSH.
It has been shown that IL-1β production is increased in immune cells exposed to radiation, leading to the production of IFN-γ. An increase of IL-1β expression in macrophages was induced by in vitro irradiation at 0.04 Gy, as well as in vivo irradiation; the initial release of IL-1β from irradiated macrophages activates T cells to produce cytokines such as IFN-γ, which in turn potentiate macrophages or APCs to produce IL-12. 61,62 Thus, an APC’s T-cell activation loop may be initiated. On the other hand, tumor-bearing B-cell–deficient mice showed a marked increase in the tumor-free fraction of mice. 63 The elimination of B cells from the splenocytes of tumor-bearing mice resulted in enhanced IFN-γ production, leading to increased IL-12 production. These effects were mediated by APC T-cell interaction via CD40. 64 Thus, B cells favor a Th2-dominant state by interfering with the interaction between APCs and antitumor T cells. They also have potential to skew immunity toward Th2 by inducing IL-4 and IL-10 or by presenting antigens. 65 -67 These facts suggest that a decrease in B-cell population may also facilitate Th1 polarization. It is well established that the B-cell lineage is especially radiosensitive among lymphocytes and hemopoietic cells, 68 and whole-body irradiation selectively decreases the percentage of B-cell population with a concomitant increase in the helper T-cell population in blood, as well as in the spleen. 15,69 Thus, selective killing of B cells by radiation throughout the body may provide conditions that favor Th1 polarization. These results indicate that low-dose radiation shifts the immunity balance toward Th1, resulting in enhancement of cell-mediated antitumor activity.
Suppression of Tumor Metastasis by Low-Dose Whole-Body γ Irradiation: Role of Matrix Metalloprotease
As already mentioned, low-dose whole-body γ irradiation (0.5 Gy × 4 times) suppressed solid tumor growth due at least in part to the elevation of intracellular antioxidants followed by activation of cell-mediated immunity. 17 We next examined the effect of γ irradiation on tumor metastasis in a mouse model of pulmonary metastasis. C57BL/6 mice were first γ irradiated at the dose of 0.5 Gy and then intravenously injected with B16F10 melanoma cells (2.0 × 105 cells) via the tail vein at 6 hours after irradiation. The mice then received 3 more irradiations (0.5 Gy/each) at 4-day intervals (total dose: 2.0 Gy). After 14 days, the lungs were excised and melanoma colonies were counted. The repeated γ irradiation significantly inhibited colony formation in the lung tissues, suppressing tumor metastasis. A previous report had also shown that 0.15 to 0.2 Gy X-ray single-dose irradiation inhibited tumor metastasis. 70 Though many factors may contribute to this effect, we focused on changes in matrix metalloproteases (MMPs) and tissue inhibitors of metalloproteases (TIMPs), which are both important factors in tumor metastasis, in order to clarify the mechanism of the inhibitory effect. Several reports show that high levels of MMP-2, as well as other MMPs, exacerbate tumor metastasis both directly and indirectly via degradation of the extracellular matrix and activation of various growth factors. 71 -73 On the other hand, tumor metastasis was inhibited in MMP-2 knockout (KO) mice. 74 There is also much evidence that redox modulation of the progression of various diseases, such as tumors, cardiac ischemic injuries, and so on, involves down/upregulation of a subset of MMPs. 75 -83
First, we investigated the changes of MMP-2 in lungs of metastasis model mice at 14 days after the first γ irradiation. Protein expression and activity of MMP-2 were both significantly reduced. A similar reduction was seen in lungs of normal mice 24 hours after a single γ irradiation. These results suggest that radiation-induced decrease of MMP-2 in lung tissues may contribute to the suppression of lung tumor metastasis.
On the other hand, TIMP-2 suppresses tumor metastasis by inhibiting the gelatinolytic activity of MMP-2 and blocking MMP-2–mediated activation of various other factors. Alteration of TIMP-2 expression in the host mice also affects tumor metastasis. For example, the adenovirus-mediated overexpression of TIMP-2 prior to or after tumor challenge results in a significant reduction of tumor metastasis. 84 The TIMP-2 alone also inhibited cell proliferation or cell migration. 85 -87 Thus, we examined changes of expression and activity of TIMP-2 in lungs of the metastasis model mice at 14 days after the first γ irradiation. We found that TIMP was increased at the protein, mRNA, and activity levels. Overall, elevation of the expression and activity of TIMP-2 in lung tissues appears to inhibit tumor metastasis. Moreover, the alteration of the MMP-2/TIMP-2 balance induced by low-dose whole-body γ irradiation might result in an increase of free TIMP-2, which could act directly on tumor cells in the lung. Besides induction of cell-mediated immunity by low-dose γ rays, we thus identified a suppressive and/or an enhancing effect of radiation on MMP-2 and/or TIMP-2, respectively. 88 It is also clear that γ irradiation at high doses (more than 8 Gy) activates MMPs and promotes tumor metastasis. 89 Thus, low-dose whole-body γ irradiation appears to inhibit solid tumor growth and tumor metastasis via its effects on cell-mediated immunity and MMPs (Figure 4).
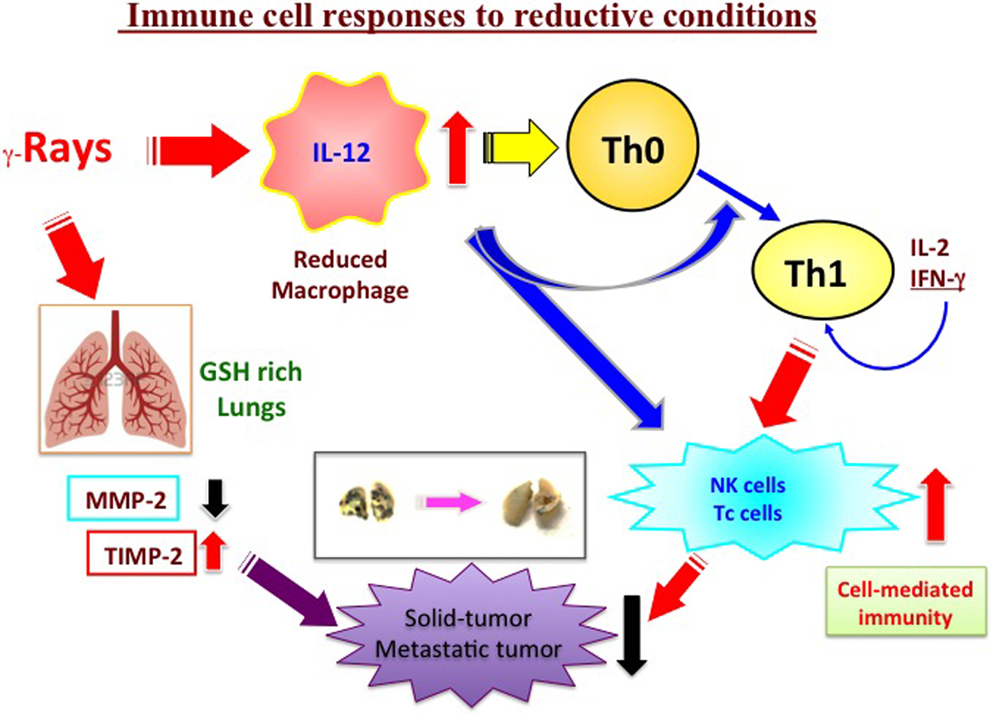
Activation of cell-mediated antitumor immunity and suppression of matrix metalloproteases (MMPs) under reductive conditions. Enhancement of intracellular reduced condition induced by low-dose γ irradiation could regulate Th1/Th2 and MMP-2/TIMP-2 balance, resulting in suppression of tumor growth and metastasis. Th indicates T helper; TIMP-2, tissue inhibitors of metalloprotease 2.
Role of ATP Signaling in Radioresistance
A small preceding/priming/adapting/conditioning dose of ionizing radiation decreases the biological effectiveness of a subsequent large challenging dose in the living body. This phenomenon of radioadaptive response is called induction of radioresistance. 90 -94 It is generally thought that various factors, including high contents of antioxidants, DNA damage-repairing activities, cell cycle control gene expression, signal transmission, reduction of apoptotic death, and so on are involved in the response. 95 -100 Living organisms have an elaborate antioxidant defense system, including SOD, CAT, GPX, and reduced GSH, to protect them against ROS. There is little doubt that these antioxidants also participate in the radioresistance induced by a small dose of ionizing radiation, as described above.
However, ATP signaling also plays a role in radioresistance by promoting DNA damage repair. In the case of double-strand breaks, ataxia telangiectasia mutated (ATM) is activated at sites of damage, and phosphorylated histone variant H2AX (γH2AX) is phosphorylated. Then, repair proteins such as tumor suppressor p53-binding protein 1 (53BP1) are accumulated at the same sites. 101 However, the mechanisms are not yet fully understood. As regard the role of ATP, focus formation of ATM-γH2AX, which is considered an index of DNA repair processes, is observed within 3 hours in human lung cancer A154 cells irradiated with γ rays at a dose of 2 Gy. Formation of these foci is significantly inhibited by pretreatment with the ectonucleotidase apyrase, whereas conversely posttreatment with ATP or UTP promotes their formation, suggesting an important role of these extracellular nucleotides in focus induction, and thus DNA damage repair. 101 The subtypes of P2 receptors participating in repair signaling were examined using various kinds of P2 receptor antagonists, and it was found that antagonists of P2Y6 and P2Y12 markedly blocked formation of the foci. Involvement of these P2Y receptors in the repair mechanisms was further confirmed using cells in which each receptor had been knocked down. Finally, it was concluded that 3 P2 receptors, P2X7, P2Y6, and P2Y12, participate in DNA repair signaling in response to γ irradiation. 102,103 That is, γ irradiation causes release of ATP into extracellular space via P2X7, and the released ATP activates P2Y6 and/or P2Y12, resulting in formation of foci consisting of γH2AX, 53BP1, and ATM, which mediate repair of DNA damage (Figure 5).
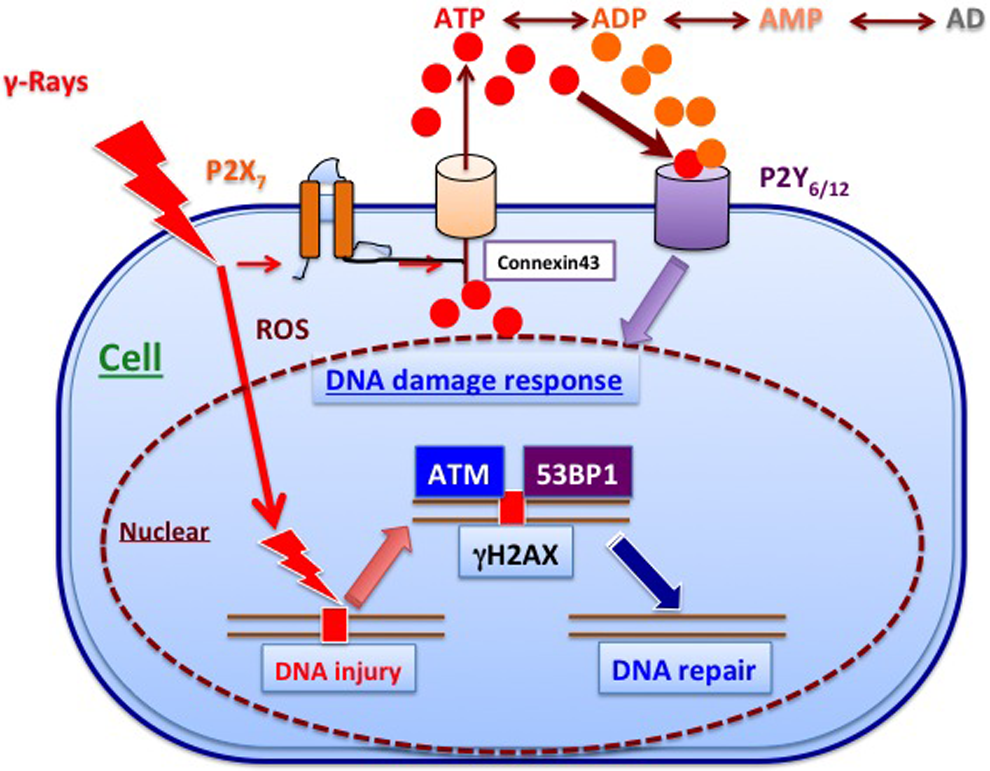
Involvement of ATP signaling in DNA damage repair. Irradiation with γ rays releases ATP via P2X7 receptor, and its metabolites activate P2Y6 and P2Y12 receptors. The activation induces the activation of ATM at sites of DNA damage, followed by induction of γH2AX and activation of 53BP1 in the irradiated cells. ATM indicates ataxia telangiectasia mutated; ATP, adenosine triphosphate; P2, purine 2.
Involvement of Adenosine A2B Receptor in Differentiation of Regulatory T Cells
We have reported that repeated 0.5 Gy γ irradiation upregulates the percentage of regulatory T (Treg) cells in various autoimmune disease models. 104 -107 In this section, we focus on the role of ATP in the induction of Treg in irradiated mice. Upon interaction with a self- or a cross-reactive antigen, naive CD4+ T helper cells activate, expand, and differentiate into various T-cell subsets with different properties, depending on cytokines. T helper cells include type 1 Th (Th1) cells and type 2 Th (Th2) cells, 108 Th17, 109,110 Th9 cells, 111,112 and regulatory subsets such as induced Treg cells and type 1 regulatory T cells. 113 -115
Cytoplasmic ATP is released to the extracellular space in response to various stresses, as described above. The released ATP is immediately degraded to adenosine by extracellular ectonucleotidases, including CD39 and CD73, and activates P1 receptors. 116 -118 Extracellular adenosine binds to multiple adenosine receptors, that is adenosine A1, A2A, A2B, and A3 receptors, which belongs to a family of G protein-coupled receptors. Stimulation of A2A receptor on T cells by adenosine inhibits T-cell receptor (TCR)-mediated T-cell proliferation and upregulation of IL-2 receptor, as well as most of the effector functions of cytotoxic T cells. 119,120 Moreover, recent studies indicate that extracellular adenosine is involved in immunosuppressive functions of Treg; immunosuppressive adenosine is produced via increased expression of CD39 and CD73, which degrade ATP to adenosine, on the Treg surface. 121,122 With this background, we hypothesized that ATP would be released from splenocytes in response to low-dose γ ray irradiation, and adenosine derived from this ATP would be involved in differentiation of naive CD4+ T cells into Treg. To test this hypothesis, we examined the involvement of P1 receptors in upregulation of Treg by low-dose γ irradiation. Among the G protein-coupled cell surface receptors A1, A2A, A2B, and A3, the A1 and A2A receptors have greater affinity for adenosine than A2B and A3. 117,118 However, the A2B receptor promotes inflammatory responses of mast cells, epithelial cells, smooth muscle cells, and fibroblasts, contributing to the pathophysiology of asthma and colitis. 123 Moreover, some pro-inflammatory cytokines, such as IL-6, are induced via activation of A2B receptor in dendritic cells. 124,125 On the other hand, it has been reported that A2B receptor protects against inflammation and excessive vascular adhesion; the levels of pro-inflammatory cytokines, including TNF-α and IL-6, were increased in A2B receptor-KO mice, whereas anti-inflammatory IL-10 was decreased. These findings imply that ATP and its metabolite, adenosine, are involved in differentiation of Treg and Th17. 126 Thus, we focused on the involvement of adenosine and P1 receptors in the differentiation of Treg. Intriguingly, we found that inhibition of adenosine A2B receptor suppressed the differentiation of Treg without suppressing T-cell activation. 127 These results suggested that stimulation of A2B receptor might promote Treg differentiation. We also showed that low-dose γ irradiation attenuates autoimmune manifestations in Murfy and Roths Laboratories (MRL)-lpr/lpr mice and collagen-induced arthritis (CIA) mice via suppression of cytokines and autoantibodies, and upregulation of Treg, but not by directly damaging lymphocytes. Moreover, γ irradiation-induced ATP release would contribute to the differentiation of Treg from naive CD4+ T cells via stimulation of adenosine A2B receptor by the ATP metabolite, adenosine. The putative role of extracellular adenosine, generated from ATP released by γ irradiation, in promoting the differentiation of Treg from naive CD4+ T cells via adenosine A2B receptor is illustrated schematically in Figure 6.

Regulatory T (Treg) cell differentiation via ATP signaling. ATP would be released in response to low-dose γ ray irradiation from splenocytes and degraded adenosine from ATP might be involved in differentiation into Treg from naïve CD4+ T cells via stimulation of adenosine A2B receptor. ATP indicates adenosine triphosphate.
Conclusions
In this review, we have introduced and reviewed the results of extensive studies by our group and others, demonstrating that ATP signaling plays an important role in the biological effects of radiation. More recently, detailed studies have also shown that transient receptor potential melastatin, a nonselective cation channel, is activated dependently on P2X7 receptor, 128 resulting in release of nucleotides such as ATP through connexin43 hemichannel. 35 Subsequently, P2Y6/P2Y12 receptor is activated, leading to a range of low-dose irradiation-induced molecular events, such as activation of epidermal growth factor-extracelluar signal-regulated kinase (EGFR-ERK)1/2, 129,130 repair of damaged DNA, 103 ROS production, induction of endogenous antioxidants, 39 and so on (Figure 7). Such a detailed understanding of the role of ATP signaling in adaptive responses, such as radiation resistance and DNA damage repair, is important, both because of the apparent beneficial effects of low-dose radiation in the living body and also because of the role of resistance in radio/chemotherapy of cancer. 131

ATP signaling and biological effects of low-dose ionizing radiation. Irradiation with low-dose γ rays releases nucleotides such as ATP through connexin 43 hemichannel, 35,36 followed by activation of P2Y6/P2Y12 receptor, leading to a range of low-dose irradiation-induced molecular events. Such an event includes activation of EGFR-ERK1/2, 129,130 DNA damage repair, 103 production of ROS, induction of antioxidants, 39 and so on. ATP indicates adenosine triphosphate; P2, purine 2; ROS, reactive oxygen species.
It has also been suggested that ATP signaling is involved in the so-called bystander effect, which is currently attracting much interest in the field of radiobiology. Both ATP and connexin43 were found to participate in the bystander effect in in vivo mouse model experiments. 10,132 In this connection, we have reported ripple-like release of ATP from a single cell irradiated with an X-ray microbeam. Clearly, the ramifications of ATP signaling remain to be fully explored.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.