
Abstract
DNA damage repair and G2/M arrest are the key factors regulating the survival of cancer cells exposed to radiation. Recent studies have shown that long noncoding RNAs (lncRNAs) play important roles in a variety of biological processes, including DNA repair, cell cycle regulation, differentiation, and epigenetic regulation. However, the knowledge about the genome scale of lncRNAs and their potential biological functions in tumor cells exposed to radiation are still unclear. In this study, we used LncRNA + mRNA Human Gene Expression Microarray V4.0 to profile lncRNA and messenger RNA (mRNA) from HeLa, MCF-7, and A549 cells after irradiation with 4 Gy of γ-radiation. We identified 230, 227, and 274 differentially expressed lncRNAs and 150, 214, and 274 differentially expressed mRNAs in HeLa, MCF-7, and A549 cells, respectively, among which there are 14 common differentially expressed lncRNAs and 22 common differentially expressed mRNAs in all of the 3 cell lines. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes pathway analysis indicated that these differentially expressed mRNAs were mainly associated with cell cycle. Further, we also predicted the target genes and functions of these differentially expressed lncRNAs. Our study on lncRNAs has greatly expanded the field of gene research in the relationship of radiation, cell cycle, and DNA damage.
Introduction
Radiotherapy is one of the most frequent methods used to treat carcinomas. 1 DNA damage repair and G2/M arrest are the key factors which regulate the survival of cancer cells exposed to radiation. 2 -5 The current models of the mechanism of DNA double-strand break (DSB) repair and G2/M arrest are based on studies of proteins. 6,7
It is estimated that 98% of the human transcriptome is noncoding RNA, which is limited with protein-coding capacity and was long thought to have no obvious molecular function (MF). 8,9 Long noncoding RNAs (lncRNAs) are a group of noncoding RNAs with length ranging from 200 nt to 100 kb. 10 The regulatory modes of lncRNAs can involve cis interactions to affect the expression of nearby genes or protein complexes. 11 Furthermore, lncRNAs can interact with DNA, RNA, and proteins to either activate or suppress gene expression, and they can bind to transcription factors or chromosome-regulating complexes. 12 -14 Long noncoding RNAs have lower levels of expression compared to the protein-coding transcripts but exhibit more tissue-specific expression and evolutionary conservation. 15 Recent studies have shown that lncRNAs play important roles in a variety of biological processes (BPs), including DNA repair, cell cycle regulation, differentiation, and epigenetic regulation. 16 -19 Especially, a number of studies have shown that lncRNA expression can be deregulated in human cancers. 20 However, the knowledge about the genome scale of lncRNAs and their potential biological functions in tumor cells exposed to radiation are still far from clear.
In this study, we used LncRNA + mRNA Human Gene Expression Microarray V4.0 to profile lncRNA and messenger RNA (mRNA) from HeLa, MCF-7, and A549 cells after irradiation with 4 Gy of γ-radiation. We identified lncRNA and mRNA expression signatures after irradiation in comparison to sham. Our results indicate that lncRNAs may exert a partial or key role in the regulation of cell cycle regulation induced by radiation.
Materials and Methods
Cell Culture and Irradiation
HeLa, A549, and MCF-7 cells were used throughout the present study. HeLa cells were cultured in RPM1640 medium supplemented with 10% fetal calf serum and A549 and MCF-7 cells were cultured in Dulbecco’s Modified Eagle’s Medium (Gibco, Scotland, United Kingdom) supplemented with 10% fetal calf serum. All cells were supplemented with appropriate penicillin and streptomycin and maintained at 37°C in a humidified incubator supplied with 5% CO2.
Cells were irradiated with γ-rays to a dose of 4 Gy using a cobalt-60 source (Beijing Institute of Radiation Medicine, Beijing, China). After irradiation, the cells were maintained at 37°C for 2 hours and then subjected to extract total RNA or prepared for other analysis.
Cell Cycle Analysis
Two hours after irradiation, cells were fixed in 70% ethanol at −20°C overnight and were washed with PBS and treated with 0.25% Triton X-100 for 15 minutes at room temperature. Cell pellets were incubated with the p-H3 antibody (1:1000, Bethyl, Montgomery, Texas) and goat anti-rabbit IgG (H+L) antibody labeled with fluorescein isothiocyanate (FITC) (1:200, KPL, Milford, Massachusetts), which was followed by incubation with RNase A and propidium iodide (PI). Stained cells were analyzed by BD FACS Calibur flow cytometer (BD Biosciences, Sparks, Maryland). Data were analyzed using CellQuest software (version 6.1).
Western Blotting
Two hours after irradiation, cells were lysed in lysis buffer (50 mmol/L Tris-HCl pH8.0, 150 mmol/L NaCl, 1% TritonX-100, 100 μg/mL PMSF). Then, the lysates were electrophoresed on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (12%) and transferred onto nitrocellulose membranes (Bio-Rad, Hercules, California). Anti-γ-H2AX (Millipore, Burlington, Massachusetts), anti-β-actin antibody (Proteintech, Rosemont, Illinois), and goat anti-mouse IgG (H+L) secondary antibody (Sungene Biotech, Tianjin, China) were used for Western blot.
Cell Colony Formation Assay
Cells were irradiated by 4 Gy and reseeded with 500 cells per well. After 12 days, cells were fixed with methanol for 30 minutes and stained with Giemsa for 30 minutes. The number of colonies with at least 50 cells was counted.
Array Data Production
Long noncoding RNA + mRNA Human Gene Expression Microarray V4.0 (cat No. 360069; CapitalBio, Beijing, China) containing 41 000 human lncRNAs and 34 000 human mRNAs was used in this study. 21 Sample preparation and microarray hybridization were performed by CapitalBio Corporation according to the manufacturer’s protocol. Briefly, total RNA was extracted using the TRIzol reagent (Invitrogen, Shanghai, China) and purified with mirVanami RNA Isolation Kit (Ambion, Austin, Texas). The quantification and quality of RNA were assessed using NanoDrop ND-1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, Massachusetts). RNA integrity and genomic DNA (gDNA) contamination were assessed by 1% formaldehyde denaturing gel electrophoresis. The complementary RNA (cRNA) amplification and labeling were performed using a CapitalBio cRNA Amplification and Labeling Kit (CapitalBio, Beijing, China) according to the manufacturer’s protocol. Controls and irradiated groups were labeled with Cy5-dCTP and Cy3-dCTP, respectively. Then, each sample was hybridized to the LncRNA + mRNA Human Gene Expression Microarray V4.0. Finally, the arrays were scanned with an Agilent DNA Microarray Scanner G2505C (Agilent Technologies, Beijing, China).
Microarray Imaging and Data Analysis
Array images were acquired by Agilent Feature Extraction (v10.7; Agilent Technologies, Beijing, China), and array data summarization, normalization, and quality control were analyzed using the GeneSpring software V13.0 (Agilent Technologies, Beijing, China). To select differentially expressed genes, we normalized the raw data with the Lowess and used threshold values of ≥1.5 and ≤−1.5-fold changes (FCs). The probes that have flags in “Detected” were chosen for further data analysis. Hierarchical clustering was performed using Adjust Data function of CLUSTER 3.0 software then further analyzed with average linkage.
The functional enrichment of the differently expressed mRNAs was analyzed by Gene Ontology (GO) enrichment analysis in the Database for Annotation, Visualization and Integrated Discovery (DAVID, http://david.abcc.ncifcrf.gov/). Gene Ontology analysis was divided into BP, cellular component (CC), and MF. The biological pathway was analyzed using Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis on DAVID.
Quantitative Real-Time PCR
Total RNA was reverse transcribed into cDNA using a PrimeScript RT reagent Kit with gDNA Eraser (Takara, Dalian, China) according to the manufacturer’s protocol. Quantitative PCR (qPCR) was performed using a TaqMan probe Ex Taq (TaKara, Dalian, China) on MyiQTM2 (Bio-Rad, Hercules, California). Relative gene expression was analyzed using the 2−ΔΔ CT method and normalized to β-actin. Specific primers and probes of genes were shown in Supplementary Table S1.
Results
Radiation Induces G2/M Arrest and DNA Damage
It is well known that the G2/M checkpoint responds to radiation-induced DNA damage. 22,23 In order to detect G2/M arrest after radiation, we performed a flow cytometric analysis of HeLa, MCF-7, and A549 cells treated with radiation. Two hours after radiation at a dose of 4 Gy, cells were marked with the p-H3 antibody and PI and analyzed with flow cytometer. Figure 1A shows that the cells treated with radiation significantly decreased in M phase. These results suggested that treatment with radiation resulted in a cell cycle arrest in G2/M phase.
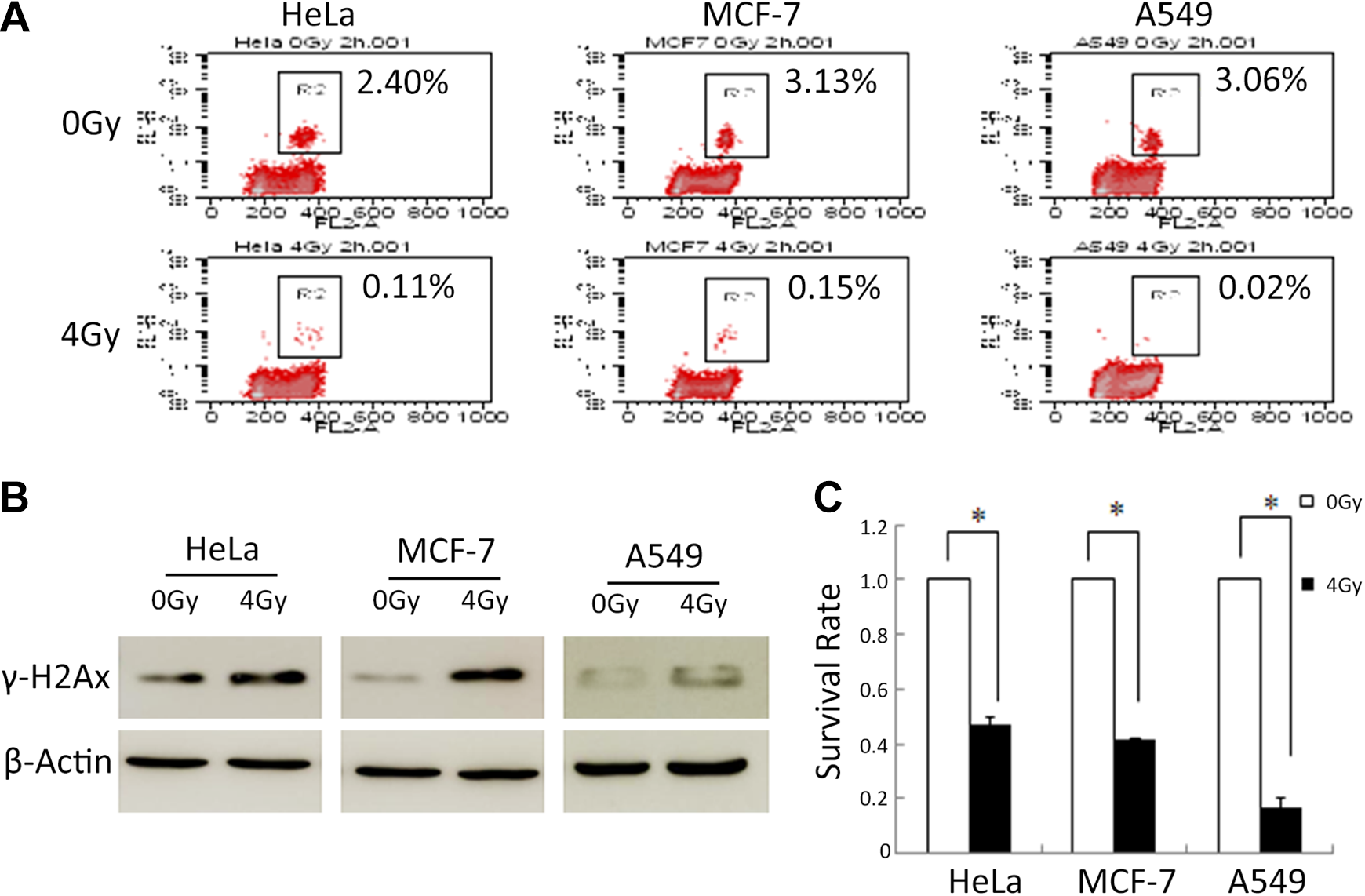
Radiation induces G2/M arrest and DNA damage. Two hours after irradiation by γ-ray, the percentages of Hela, MCF-7, and A549 cells in M phase were analyzed by flow cytometer (A), and the γ-H2AX expression was analyzed by Western blot (B). C, Two hours after irradiation, cells were reseeded. After 12 days, the number of colonies was counted and the survival rate was analysis. Representative results are shown, and mean ± SD are presented. N = 3, *P < .05 versus control group. SD indiates standard deviation.
Phosphorylated H2AX (γ-H2AX) is a marker of DSBs. 24 To investigate the DNA damage, we analyzed γ-H2AX expression in cells 2 hours after exposure to IR by Western blotting. As shown in Figure 1B, the expression level of γ-H2AX was markedly increased in cells treated with radiation compared with control cells.
The radiosensitivity of cells was examined by clonogenic assay after exposure to radiation at dose of 4 Gy. After 12 days treated by radiation, colonies were counted to determine the cell viability. As shown in Figure 1C, compared with the control group, the survival cells treated with radiation were significantly reduced. This suggested that HeLa, MCF-7, and A549 cells were relatively sensitive to radiation.
Differentially Expressed LncRNAs and mRNA in Tumor Cell Lines After Irradiation Exposure
To identify potential lncRNAs and mRNAs that may be involved in DNA damage and G2/M checkpoint induced by radiation, Agilent Feature Extraction (version10.7, Agilent Technologies) was utilized to analyze acquired array images to get raw data. GeneSpring (version 13.0, Agilent Technologies) was employed to finish the data summarization, normalization, and quality control with the raw data. To begin with, the raw data were normalized with the Lowess. The probes that have flags in “Detected” were chosen for further data analysis. Differentially expressed lncRNAs or mRNAs were then identified through FC. The threshold set for upregulated or downregulated RNAs was FC ≥1.5. We scanned 14 784 lncRNA probes and 20 875 mRNA probes in 3 cell lines; 230, 227, and 274 differentially expressed lncRNAs and 150, 214, and 274 differentially expressed mRNA were identified in HeLa, MCF-7, and A549 cells, respectively (Supplementary Tables S2 and S3). The 10 most differentially expressed lncRNAs and mRNAs are shown in Tables 1 and 2.
The 10 Most Differentially Expressed LncRNAs (5 Upregulated and 5 Downregulated lncRNAs) in Different Cells.

Abbreviations: FC, fold change; lncRNA, long noncoding RNA.
The 10 Most Differentially Expressed mRNAs (5 Upregulated and 5 Downregulated mRNAs) in Different Cells.

Abbreviations: FC, fold change; mRNA, messenger RNA.
Validation of Differential LncRNA or mRNA Expression by Quantitative Real-Time PCR
To verify the results of microarray, we detected the expression of 9 lncRNAs and 5 mRNAs selecting randomly from the differentially expressed transcripts using qPCR. β-actin was used as a normalization control. As shown in Figure 2, the expression dates of 67% were consistent with the lncRNA and mRNA array analysis, which indicates the reliability of the microarray data.
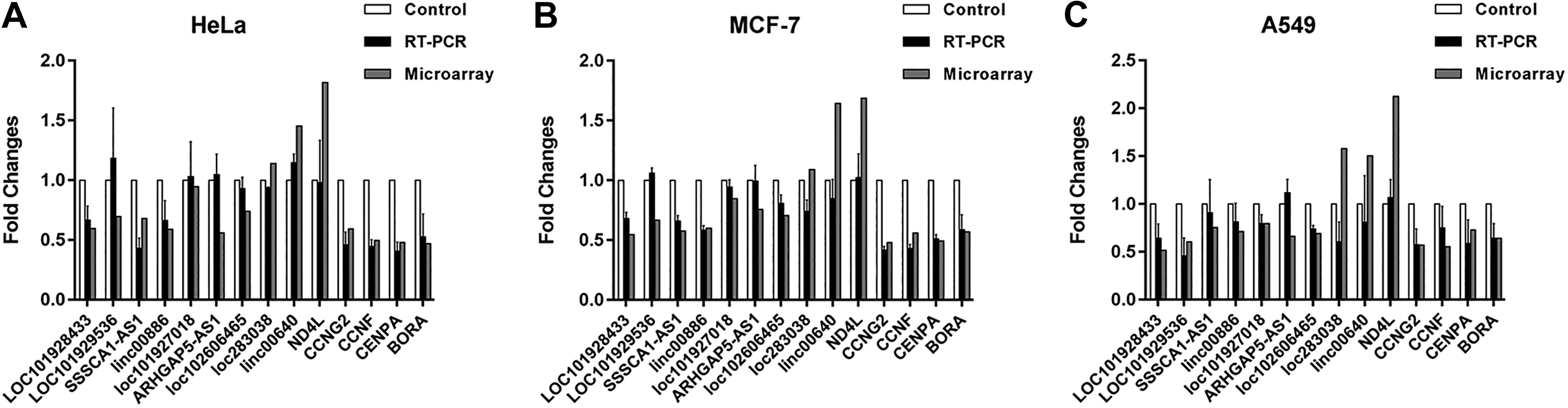
Validation of differential lncRNAs and mRNAs expression by qRT-PCR. Nine differentially expressed lncRNAs and 5 differentially expressed mRNAs were randomly selected and their expression in HeLa (A), MCF-7 (B), and A549 (C) was analyzed by qRT-PCR. The qRT-PCR data are presented as the mean ± SD. LncRNA indicates long noncoding RNA; mRNA, messenger RNA; qRT-PCR, quantitative real-time polymerase chain reaction; SD, standard deviation.
Potential Function Identification of Differentially Expressed mRNAs
The differentially expressed mRNAs in the 3 cell lines were selected for functional enrichment analysis. As shown in Figure 3 and Supplementary Table S4, we found that the differentially expressed mRNAs were mostly enriched in microtubule binding and protein binding in its MF, cell division, and mitotic metaphase plate congression in its BP and midbody, microtubule cytoskeleton, and microtubule in its CC. Kyoto Encyclopedia of Genes and Genomes pathway analysis indicated that the mRNAs were mainly enriched in cell cycle pathway.

Gene Ontology enrichment and KEGG pathway analysis of differentially expressed mRNAs. The top 10 most enriched GO categories including molecular function, biological process, cellular component, and pathways of differentially expressed mRNAs were calculated and plotted. A, Hela, (B) MCF-7, and (C) A549. GO indicates Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; mRNA, messenger RNA.
Long Noncoding RNA Target Analysis of Differentially Expressed LncRNAs
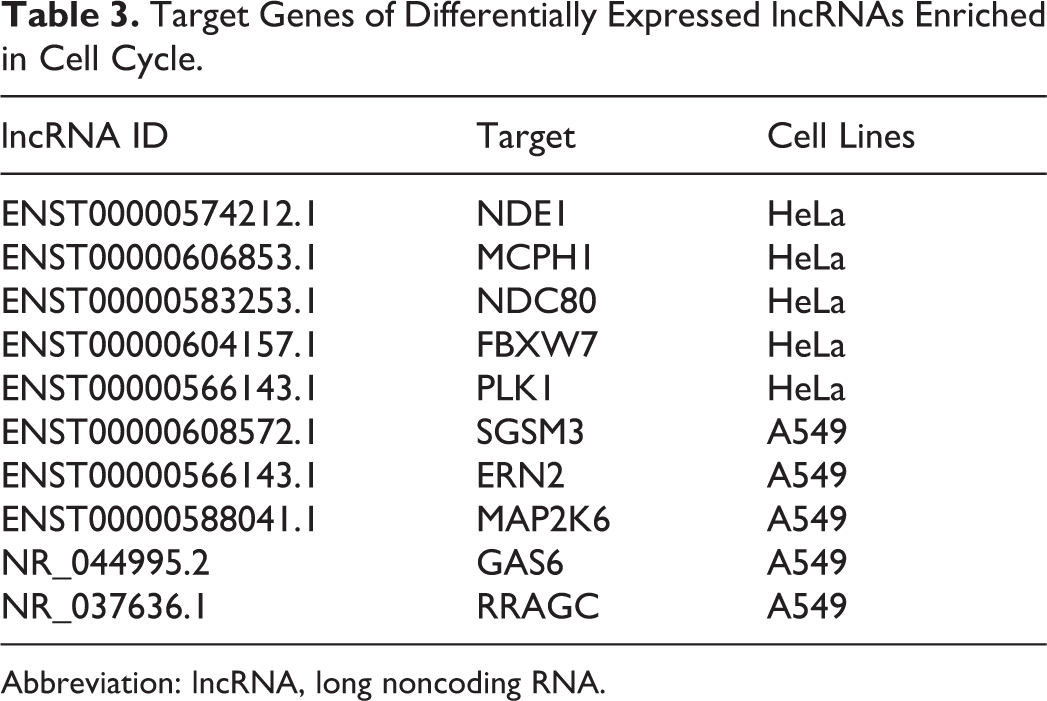
Long noncoding RNAs can regulate target genes in cis. The rationale for identifying cis target genes was that the lncRNAs should be in relatively close proximity to the protein-coding genes. Therefore, all genes in the proximity of the lncRNA loci (10 kb upstream or downstream) were selected as target genes, and the enrichment of specific MFs among the target genes was analyzed to predict the functions of lncRNAs. In order to further identify the functions of each individual lncRNA, we analyzed differentially expressed lncRNAs in each cells. There were 150 genes in HeLa, 196 genes in A549, and 135 genes in MCF-7 predicted to be the targets of the lncRNAs (Supplementary Table S5). We also conducted the GO enrichment and KEGG pathway analysis for these lncRNA-targeted genes (Supplementary Table S6). Ten genes enriched in cell cycle are listed in Table 3.
Target Genes of Differentially Expressed lncRNAs Enriched in Cell Cycle.
Abbreviation: lncRNA, long noncoding RNA.
Differentially Expressed LncRNAs and mRNAs in All of the 3 Cell Lines
Venn diagrams of the numbers of differentially expressed lncRNAs and mRNAs in HeLa, MCF-7, and A549 cell are shown in Figure 4. The results showed that there were 14 common differentially expressed lncRNAs and 22 common differentially expressed mRNAs in the 3 cell lines (Figure 4A and B and Supplementary Table S7). Hierarchical clustering of these common differentially expressed lncRNAs and mRNAs was performed to display the same expression patterns of each lncRNA or mRNA in different cell lines (Figure 4C and D). We also selected the 22 differentially expressed mRNAs for GO analysis. The results showed that these mRNAs were enriched in cell cycle (Figure 4E and Supplementary Table S8).

Differentially expressed lncRNAs and mRNAs in all of in HeLa, MCF-7, and A549. Venn diagrams of the numbers of differentially expressed lncRNAs (A) and mRNAs (B) in HeLa, MCF-7, and A549. Hierarchical clustering of common differentially expressed lncRNAs (C) and mRNAs (D) in HeLa, A549, and MCF-7. E, GO enrichment analysis of common differentially expressed mRNAs. GO indicates Gene Ontology; LncRNA, long noncoding RNA; mRNA, messenger RNA.
Discussion
Radiotherapy is one of the mainstream approaches for the treatment of carcinomas. The purpose of radiotherapy is to kill tumor cells as efficiently as possible and reduce recurrence. Radiation results in a series of changes in tumor, but which molecules change and how these changes regulate tumor cells after radiation exposure is still unclear. Basic research on these effects is urgently needed. DNA damage repair and G2/M arrest are the important factors which regulate cell survival after radiation. The current models of the mechanism of DNA damage repair and G2/M arrest are based on studies of proteins. In order to screen the lncRNAs which involved in the regulation DNA damage repair and G2/M arrest after radiation, we further carried out a high-throughput lncRNA + mRNA microarray from HeLa, MCF-7, and A549 cells 2 hours after 4 Gy radiation and discovered a series of differentially expressed mRNAs and lncRNAs after radiation.
One hundred fifty mRNAs in HeLa, 214 mRNAs in MCF-7, and 274 mRNAs in A549 were identified as differentially expressed transcripts between the irradiated group and the control group. Among these differentially expressed mRNA, some mRNAs have been reported to be associated with radiation. Naito et al found that Cyclin G2 (CycG2) is localized at DNA repair foci following DNA damage induced by ionizing radiation and that CycG2 regulates the dephosphorylation of several factors necessary for DNA repair. 25 Wang et al proposed that the level of IL-8 may predict radiation-induced lung toxicity in non-small cell lung cancer. 26 MDM2-p53 pathway is well known as a key factor in the protection against cancer and confers radiosensitivty. 27 Series of other mRNAs such as SERHL2, PSRC1, BTG2, ATF3, and ARHGAP29 are also considered to be related to radiation. 22,23,28 -30 Meanwhile, many mRNAs such as KIAA1751, ERN1, and PIF1 are identified to be related to radiation for the first time in our profile. These findings provide new insights into relationship between radiation and cell survival.
Gene Ontology term enrichment analysis showed that many mRNAs were involved in cell division and cell cycle. It is well known that Aurora-A (AURKA) is a key regulator in the G2/M transition and knocking down the expression of Aurora-A gene induces G2/M phase arrest. 31,32 Brooks et al showed that the localization of G2E3 plays a role in cell cycle regulation and the cellular response to DNA damage. 33 Other studies reported that BORA, AURKE, and PLK1 play important roles in the regulation of mitosis. 34 -37 These analyses indicated that radiation may regulate tumor process by altering the expression of cell cycle–related molecules.
Studies about the function of lncRNAs are difficult to carry out, for most of the lncRNAs are not determined, and there is no existing database that could be used to find their functional annotations. Long noncoding RNAs are well known to regulate target genes in the proximity of its upstream or downstream by 10 kb. To predict the potential functions of differently expressed lncRNAs after radiation, we identified their targets and analyzed the function of these coding genes. The GO enrichment analysis revealed that these targets were associated with different biological progress in different cancer cells. Targets in HeLa were enriched in transcription, targets in MCF-7 were enriched in regulation of cellular glucuronidation, and targets in MCF-7 were enriched in regulation of gene expression. Ten targets were found to be associated with cell cycle. Their matched lncRNAs include ENST00000574212.1, ENST00000606853.1, ENST00000583253.1, ENST00000604157.1, ENST00000566143.1, ENST00000608572.1, ENST00000566143.1, ENST00000588041.1, NR_044995.2, and NR_037636.1. It needs further study for the effect of these lncRNAs on cell cycle after radiation.
Furthermore, there were 14 common differentially expressed lncRNAs and 22 common differentially expressed mRNAs in HeLa, MCF-7, and A549 cells. The expression patterns of these differentially expressed mRNAs or lncRNAs are similar. Gene Ontology analysis indicated that these common differential mRNAs were enriched in cell cycle, consistent with the analysis in each cells. From these results, we considered that the function of these common mRNAs and lncRNAs are independent of cancer type.
In summary, we discovered a profile of mRNAs and lncRNAs differentially expressed 2 hours after irradiation with 4 Gy. .Although the mechanisms of the discovered lncRNAs in radiation damage regulation remain to be elucidated, our study on lncRNAs has greatly expanded the field of gene research in the relationship of radiation, cell cycle, and DNA damage.
Supplemental Material
Supplementary_Table_S1 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S1 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S2 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S2 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S3 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S3 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S4 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S4 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S5 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S5 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S6 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S6 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S7 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S7 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Supplemental Material
Supplementary_Table_S8 - Screening of Long Noncoding RNAs Induced by Radiation Using Microarray
Supplementary_Table_S8 for Screening of Long Noncoding RNAs Induced by Radiation Using Microarray by Yilong Wang, Qi Wang, Shuangjing Chen, Yingchun Hu, Chang Yu, Ruixue Liu and Zhidong Wang in Dose-Response
Footnotes
Authors’ Note
Y.W. and Q.W. contributed equally as cofirst authors.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Natural Science Foundation of China (81573083 and31770913to Z.W.).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.