
Abstract
The increasing use of ionizing radiation (IR) in medical diagnosis and treatment has caused considerable concern regarding the effects of occupational exposure on human health. Despite this concern, little information is available regarding possible effects and the mechanism behind chronic low-dose irradiation. The present study assessed potential genomic damage in workers occupationally exposed to low-dose X-rays. A variety of analyses were conducted, including assessing the level of DNA damage and chromosomal aberrations (CA) as well as cytokinesis-block micronucleus (CBMN) assay, gene expression profiling, and antioxidant level determination. Here, we report that the level of DNA damage, CA, and CBMN were all significantly increased. Moreover, the gene expression and antioxidant activities were changed in the peripheral blood of men exposed to low-dose X-rays. Collectively, our findings indicated a strong correlation between genomic instability and duration of low-dose IR exposure. Our data also revealed the DNA damage repair and antioxidative mechanisms which could result in the observed genomic instability in health-care workers exposed to chronic low-dose IR.
Introduction
Occupational radiation exposure is very common, and a variety of man-made radiation types are extensively used by the health-care, research, and nuclear power industries. 1,2 In just health care alone, more than two million workers are exposed to radiation on a daily basis. 3 The increasing use of ionizing radiation (IR) in medical diagnosis and treatment has raised concerns about its potential long-term effects on human health, as prolonged exposure to low-dose radiation has been found to cause adverse health effects including genomic instability and cancer. 4,5
Given this, recent years has seen increased interest in understanding the biological effects of chronic low-dose radiation exposure and its relationship to genomic instability. 6 This work has shown that occupational radiation exposure leads to increased genomic instability; however, little information is available regarding the effects and possible mechanisms of low-dose radiation over a protracted period of time. Ionizing radiation induces a variety of DNA damage, including DNA double-strand breaks (DSBs) and single strand breaks (SSBs). 7 There is a strong correlation between the incidence of chromosomal abnormalities and both low- and high-dose IR. 8 Moreover, previous studies have reported an increase in chromosomal aberrations (CA) in the lymphocytes of workers occupationally exposed to IR when compared with unexposed workers. 9
Radiation-induced genomic instability is a delayed and persistent problem, as the effect of IR exposure manifests in the unirradiated progeny of irradiated cells. 10,11 Ionizing radiation ionizes water molecules and/or directly ionizes target molecules, triggering the formation of reactive oxygen species (ROS). 12 Reactive oxygen species play central roles in cell fate determination, as they function as second messengers and modify various signaling molecules, ion channels, transporters, and protein kinase components. 13,14 Critically, long-lasting ROS will continue to damage the cellular genome and other important biomolecules over the long term. 15 Ionizing radiation also induces DNA damage responses, such as the activation of checkpoint pathways and DNA damage repair mechanisms. 16 Therefore, we sought to study the relationship between antioxidant enzyme activity and long-term exposure to low-dose IR.
Regular low-dose occupational exposure to IR may induce deleterious health effects, which may be of particular interest to medical radiation workers who work in interventional radiology diagnostics. 17 The aim of this study was to expand on previous investigations by conducting a comprehensive analysis of the potential health risks to health-care workers, who are exposed to low-dose IR. Human peripheral blood lymphocytes are able to retain the signature of radiation-induced DNA damage; 18 therefore, we investigated the level of DNA damage, CAs, cytokinesis-block micronucleus (CBMN), gene expression profile, and antioxidant status in hospital workers occupationally exposed to low doses of X-ray radiation.
Materials and Methods
Population and Sample Collection
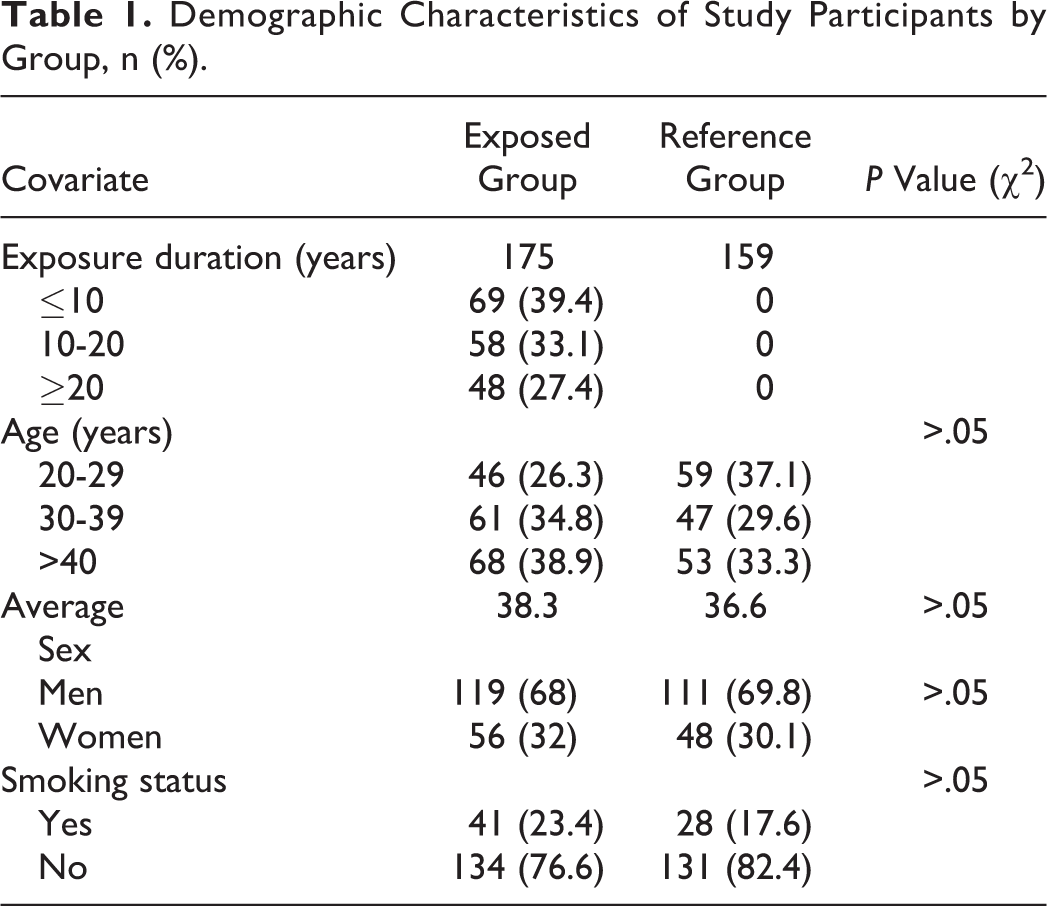
The study population consisted of 175 healthy individuals who had been occupationally exposed to low levels of X-ray radiation, and the type of exposure was whole-body irradiation. The occupational exposure to IR is monitored by personal thermoluminescence dosimeter every 3 months. The cumulative effective dose in exposed group ranged from 2.81 to 416.43 mSv (38.41 ± 27.36 mSv) based upon their duration of X-ray exposure, which ranged from 1 to 31 years. The cumulative effective dose of exposed group with the duration of ≤10 years ranged from 2.81 to 178.73 (18.28± 16.29) mSv, the cumulative effective dose of the exposed group with the duration of 10 to 20 years ranged from 25.43 to 369.45 (92.11± 85.62) mSv, whereas those with >20 years ranged from 48.64 to 416.43 (185.15± 143.51) mSv. No limitations regarding age, sex, or smoking status were imposed during the sample selection process. A control group was established, which included 159 nonexposed workers who were selected from the same hospital; controls were matched for age, sex, and smoking habits. The sample demographic characteristics were determined via questionnaire, which was administered to all participants in both groups (Table 1). The study protocol was approved by the ethics committee of the hospital and all participants gave their written informed consent.
Demographic Characteristics of Study Participants by Group, n (%).
Peripheral blood samples (10 mL) were collected from exposed workers and controls, and used immediately in CA, CBMN, and DNA damage assays as well as in gene expression profiling. Oxidative stress biomarker levels were analyzed in peripheral blood serum, selected biomarkers including malondialdehyde (MDA), glutathione (GSH), and superoxide dismutase (SOD).
Assessment of DNA Damage Using Single-Cell Gel Electrophoresis (Comet Assay)
An alkaline comet assay was used to detect DNA single- and double-strand breaks (SSBs and DSBs, respectively). Peripheral blood mononuclear cells were isolated using BD Vacutainer™ CPT™ Cell Preparation Tubes with sodium heparin, according to the manufacturer’s instructions (Becton Dickinson, New Jersey). Briefly, lymphocytes were suspended in phosphate-buffered saline at a concentration of 1 × 105/mL and the harvested cells were then mixed with 0.75% low-melting agarose at 37°C. Cells were then spread on a fully frosted slide that had been precoated with 0.75% normal agarose. The slide was then immersed in lysis buffer for 2.5 hours at 4°C. Each slide was transferred to a horizontal electrophoretic tank with Tris/borate/EDTA (TBE) buffer and incubated for 20 minutes. Electrophoresis was performed at 30 V for 20 minutes in TBE buffer. After electrophoresis, the slide was immersed in a neutralizing buffer for 20 minutes at 4°C and stained with ethidium bromide. The slides were the imaged using fluorescence microscopy (ETLPSE 90i; Nikon, Tokyo, Japan). The percentage of total fluorescence in the tail (TDNA%), tail moment (TM), and the olive tail moment (OTM) of at least 200 cells per slide was recorded and analyzed using CASP software (Wroclaw, Poland).
Chromosomal Aberrations Analysis
Chromosomal aberrations were quantified as previously described. 19 Briefly, peripheral blood was incubated in 4.5 mL RPMI 1640 medium containing 20% fetal bovine serum and phytohemagglutinin under 5% CO2 at 37°C for 48 hours. Colchicine were added to the culture medium for a final concentration of 0.06 μg/mL, and the cells were cultured for an additional 6 hours. Lymphocytes were harvested for hypotonic treatment, fixed and conventionally prepared for Giemsa staining. At least 100 metaphases cells with well-spread chromosomes were analyzed for each individual of the 2 groups (experimental and control). Chromosomal aberrations type—including dicentric chromosomes (dis), ring chromosomes (r), and acentric fragments (ace)—were recorded and reported as total aberrations.
Analysis of Cytokinesis Block Micronucleus
For each donor, 3 parallel blood cultures were established within 48 hours after sample collection. Whole blood (1 mL) was added to 10 mL of RPMI 1640 culture medium, and cultures were incubated at 37°C for 72 hours. Cytochalasin B was added for a final concentration of 6 mg/mL; cultures were then incubated for 44 hours after culture initiation to block cytokinesis. Cells were harvested at 72 hours, fixed with methanol and acetic acid (3:1) and dropped onto dry clean slides and stained with 5% Giemsa. For each slide, micronuclei were scored in at least 1000 binucleated cells for each experimental point according to the criteria proposed by ZEISS MetaSystem (Jena, Germany).
Gene Expression Profiling
Data regarding gene expression profiling was obtained from 12 donors. The 12 donors were stratified into 4 groups based upon their duration of X-ray exposure (0, ≤ 10 years, 10-20, and ≥ 20 years group). Peripheral blood cell messenger RNA (mRNA) was isolated using an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Gene expression profiling was analyzed using a Human DNA Damage Signaling pathway PCR Array, which included 82 key genes (Wcgene Biotech, Shanghai, China). Analysis was conducted according to the manufacturer’s protocol. Each array experiment was normalized to the housekeeping genes β-actin (ACTB) and glyceraldehyde 3-phosphate dehydrogenase as an endogenous controls. The relative gene expression levels of target genes were calculated using the 2−ΔΔCt method. 20 Genes that had significantly different expression were analyzed using IPA 5.0 (Ingenuity Systems Inc, California). Data were normalized to the reference gene ACTB using the cycle threshold (Ct) values. The log2 (fold-change) was calculated based on the 2−ΔΔCt method and a log-ratio of ±1 was considered statistically significant. A heatmap based on the microarray data was generated using Heatmap Illustrator (Wuhan, China), version 1.0. Pearson correlation coefficient was used to assess the dose–dependence relationship of mRNA expression for selected genes. Two main clusters showed genes that were up- or downregulated.
Analyses of Oxidative Stress Biomarkers
Levels of the oxidative stress biomarkers SOD, GSH, and MDA were quantified in peripheral blood serum using a commercially available ELISA kit in accordance with the manufacturer’s instructions (MLBio, shanghai, China). Briefly, the kit assayed human MDA, SOD, and GSH levels in the collected samples using purified human MDA, SOD, and GSH primary antibodies to coat microtiter plate wells, resulting in a solid-phase antibody. Malondialdehyde, SOD, or GSH was then added to the respective wells to combine with the primary, horseradish peroxidase (HRP)-labeled antibody, resulting in an antibody-antigen-enzyme-antibody complex. After washing completely, TMB substrate solution was added, which turns blue in an HRP enzyme-catalyzed reaction. The reaction was terminated by adding a sulfuric acid solution. The final color change was measured spectrophotometrically at a wavelength of 450 nm. Sample concentrations of MDA, SOD, and GSH were determined by comparing sample’s optical density to a standard curve.
Statistical Analysis
Statistical analyses were conducted using Graph Pad Prism 5.0 and SPSS software package 20.0 (SPSS, Chicago, Illinois). Results are expressed as mean ± standard error of the mean. A crosstabs χ2 test was used to analyze the demographic characteristics and CA of the exposure and control groups. A Student t test was performed to determine the statistical significance of TDNA%, TM, OTM, oxidative stress biomarkers, and CBMN results between the exposure and reference groups. A P value of less than .05 was considered statistically significant.
Results
General Characteristics of the Study Populations
The demographic characteristics of both exposed and control groups are shown in Table 1. In total, 334 participants (175 exposure and 159 controls) were involved in the study. Of the 175 workers exposed to interventional radiological and diagnostic X-rays, 119 were males and 56 were females with an average age of 38.3 years. Of the 159 healthy control participants, 111 were males and 48 were females with an average age of 36.6 years. Both groups were similar in distribution of gender, age, and smoking habits. A crosstabs analysis revealed no significant differences in the demographic characteristics between the 2 groups, as indicated by the matched demographic characteristics for both the exposure and reference groups as shown in Table 1.
DNA Damage in Lymphocytes
To analyze the relationship between the level of DNA damage and duration of low-dose IR exposure, the exposure group was stratified into 3 subgroups based upon exposure duration exposed to X-rays for interventional radiological and diagnostic X-ray exposure (≤ 10, 10-20, and ≥ 20 years). The level of DNA damage was represented by the percentage of DNA in the comet tail (TDNA%), the TM, and the OTM, as determined by single-cell gel electrophoresis (comet assay). Representative comet images of lymphocytes from the reference and exposure groups were shown in Figure 1A and B, respectively. We found that lymphocytes obtained from X-ray-exposed workers displayed remarkably obvious tails (Figure 1B). As shown in Figure 1C, D, and E, TDNA%, TM, and OTM of lymphocytes were all significantly greater for X-ray-exposed participants compared to healthy controls. This finding indicated that there was a greater number of broken DNA in X-ray-exposed individuals. Critically, the level of DNA damage increased with increasing duration of IR exposure. Taken together, these results revealed a statistically significant relationship between DNA damage in lymphocytes and duration of low-level IR exposure.

DNA damage of lymphocytes in X-ray-exposed (exposure group) and nonexposed (reference group) workers as detected by a comet assay. (A) Representative comet image of lymphocytes from the reference group. (B) Representative comet image of lymphocytes from the exposure group. Tail DNA% (C), tail moment (D), and olive tail moment (E) values in lymphocytes of workers in the exposure group were all significantly greater than those in the reference group. The level of DNA damage was represented as the mean of 3 independent experiments and at least 200 cells were counted (*P < .05, **P < .01).
Cytokinesis-Block Micronucleus Analysis in Lymphocytes
We next examined the CBMN in peripheral blood lymphocytes, which is a marker for evaluating the genomic instability caused by radiation exposure. 21 This analysis was conducted in both exposure and reference groups. The results of the CBMN assay are reported as total number of micronucleus (MN) per 1000 binucleated cells and were shown in Figure 2A. Figure 2C showed representative MN images in a binucleated lymphocyte from X-ray-exposed workers. As shown in Figure 2A, results showed a relationship between CBMN frequency and duration of radiation exposure in the peripheral blood lymphocytes of X-ray-exposed workers. Analysis revealed significantly greater CBMN frequency in peripheral blood lymphocytes obtained from X-ray-exposed workers of all 3 subgroups (≤ 10, 10-20, and ≥ 20 years) when compared with controls. Consistent with the comet assay results, these findings indicated that the CBMN frequency increased in workers who had been exposed to chronic low-levels of IR.
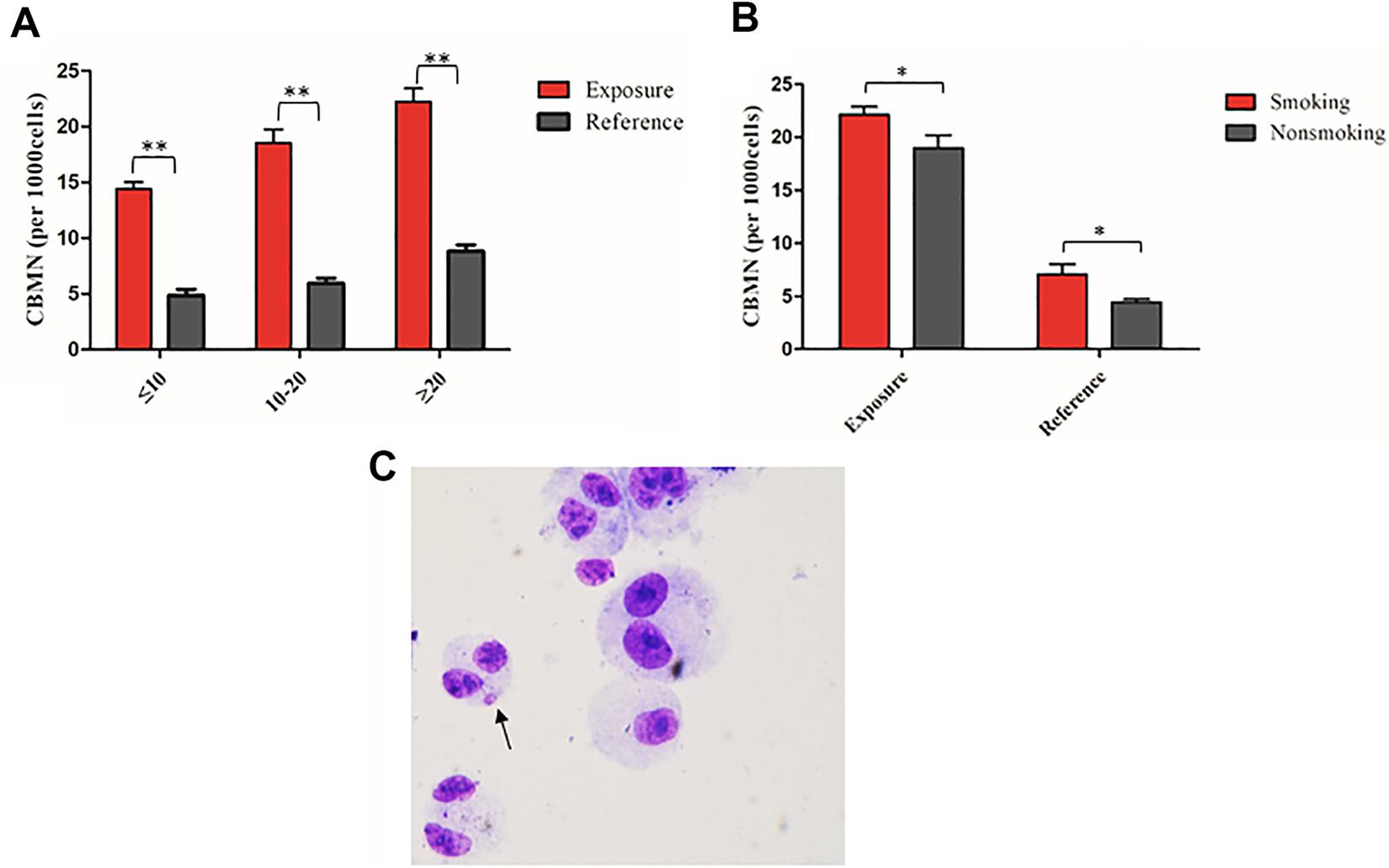
Cytokinesis-block micronucleus in the lymphocytes of the exposure and reference groups. (A) The rate of CBMN in lymphocytes of the exposure group was greater than that of the reference group. (B) Cytokinesis-block micronucleus frequency in lymphocytes of smoking workers was greater than that of nonsmoking workers in both the exposure and control groups. (C) Representative micronucleus image in a binucleated lymphocyte of workers exposed to low-dose IR. Experiments were performed independently in triplicate; more than 1000 cells were counted in each experiment (*P < .05, **P < .01). CBMN indicates cytokinesis-block micronucleus; IR, ionizing radiation.
Aware that there could have been confounding factors like smoking status—which may have affected micronucleus rate in the lymphocytes—we next stratified the exposure and reference groups into 2 subgroups (smoking and nonsmoking). For each subgroup, there were significant differences in the CBMN frequency in lymphocytes between X-ray-exposed workers and controls. Moreover, CBMN frequency was significantly higher in X-ray-exposed smokers when compared with those X-ray-exposed nonsmokers (Figure 2B). Taken together, our results were in agreement with previous work showing that smoking status affected micronucleus rate in the lymphocytes. 22
Chromosome Aberration Analysis in Lymphocytes
Chromosomal aberrations is known to be a sensitive marker for evaluating the genetic damage caused by radiation exposure. 23 To confirm CA changes after hospital workers were occupationally exposed to low levels of IR, we next examined the number of dicentric (dis), ring (r), and acentric fragment (ace) chromosomes in peripheral lymphocytes from exposed workers and controls. Since smoking status may have also affected DNA damage, the exposure and reference groups were both stratified by smoking status into smoking and nonsmoking groups. As shown in Figure 3A, there was a significantly higher CA rate in smoking workers exposed to low levels of X-rays. Comparatively, there was no significant difference in CA rate in nonsmoking X-ray-exposed workers (P > .05). Since age might affect the condition of the DNA in the lymphocytes, the exposure and reference group were both stratified by age into 3 subgroups (20-29, 30-39, and ≥ 40 years old). For each subgroup based on age, there was significantly higher CA in the workers exposed to low-dose X-ray radiation compared to the reference group, and no significant difference was found among subgroups in either the exposure or reference group (Figure 3B). Figure 3C, D, and E show representative normal chromosomal images obtained from healthy controls as well as abnormal images obtained from X-ray-exposed workers. As shown in Table 2, the incidence rate of total CA in X-ray-exposed workers was significantly different (P < .05) when compared with workers who had never been exposed to IR. Collectively, these results indicated that CA frequency increased in health-care individuals occupationally exposed to IR.

Chromosomal aberrations rate in the lymphocytes of exposure and reference workers. (A) Chromosomal aberrations rate in low-dose X-ray-exposed workers was greater than that of the reference group. Chromosomal aberrations rate of smoking participants was higher than that of nonsmoking participants in the exposure group. (B) Chromosomal aberrations rate in lymphocytes of workers exposed to low-dose X-ray radiation was higher than that in reference group for different age subgroups. (C) Representative normal chromosomal image obtained from the reference group. (D) and (E), Representative, abnormal chromosomal images of the low-dose, X-ray-exposed workers. A 2-way ANOVA was used to test the interactions between smoking and CA rate in lymphocytes (*P < .05, **P < .01). ANOVA indicates analysis of variance; dis, dicentric chromosomes; r, ring chromosomes.
Effect of Low-Dose Ionizing Radiation on the Frequencies of CA-Total, dis, r, and ace in the Exposure and Reference Groups.

Abbreviations: CA, chromosome aberration; dis, dicentric chromosomes; r, ring chromosomes; ace, acentric fragments.
Gene Expression Profiling in Peripheral Blood Lymphocytes
Our results showed the increased genomic instability after X-ray exposure, as determined by significant increases in DNA damage level, CA, and CBMN. These results provided direct evidence for the adverse effects of low-dose IR exposure on genetic material. Given this, we next conducted DNA damage-related gene expression profiling to explore expression level changes following low-dose IR exposure. Total RNA was isolated from the peripheral blood of the 2 groups and the expression levels of DNA damage-related genes were evaluated. The identity of the 82 selected genes (Supplemental Table 1) and a heatmap for the data from each sample were shown in Figure 4. As shown, the gene expression in workers who had been exposed to low-dose IR for less than 10 years was similar to the expression profile of the reference group. Comparatively, expression of DNA damage-related genes increased significantly in workers who had been exposed to low-dose X-ray radiation for more than 20 years. In particular, men in the IR-exposed group had 18 significantly upregulated genes when compared with the reference group: BIRC5, GTSE1, CCND1, CDC25C, E2F1, MKI67, CCNA2, CDK1, ATM, CDK5R1, BRCA2, CCNE1, BRCA1, TFDP2, CDC25A, CDKN2B, ATR, and RB1. When compared with the reference group, 7 genes (RAD51, CDC6, MCM4, TP53, MCM2, MCM3 and KPNA2) were significantly downregulated in this same X-ray-exposed group. These differentially expressed genes between the 2 groups were predominantly involved in biological processes and pathways, including DNA repair, cell cycle, apoptosis, and genome stability. Finally, there was a significant relationship between upregulated gene expression and duration of IR exposure.
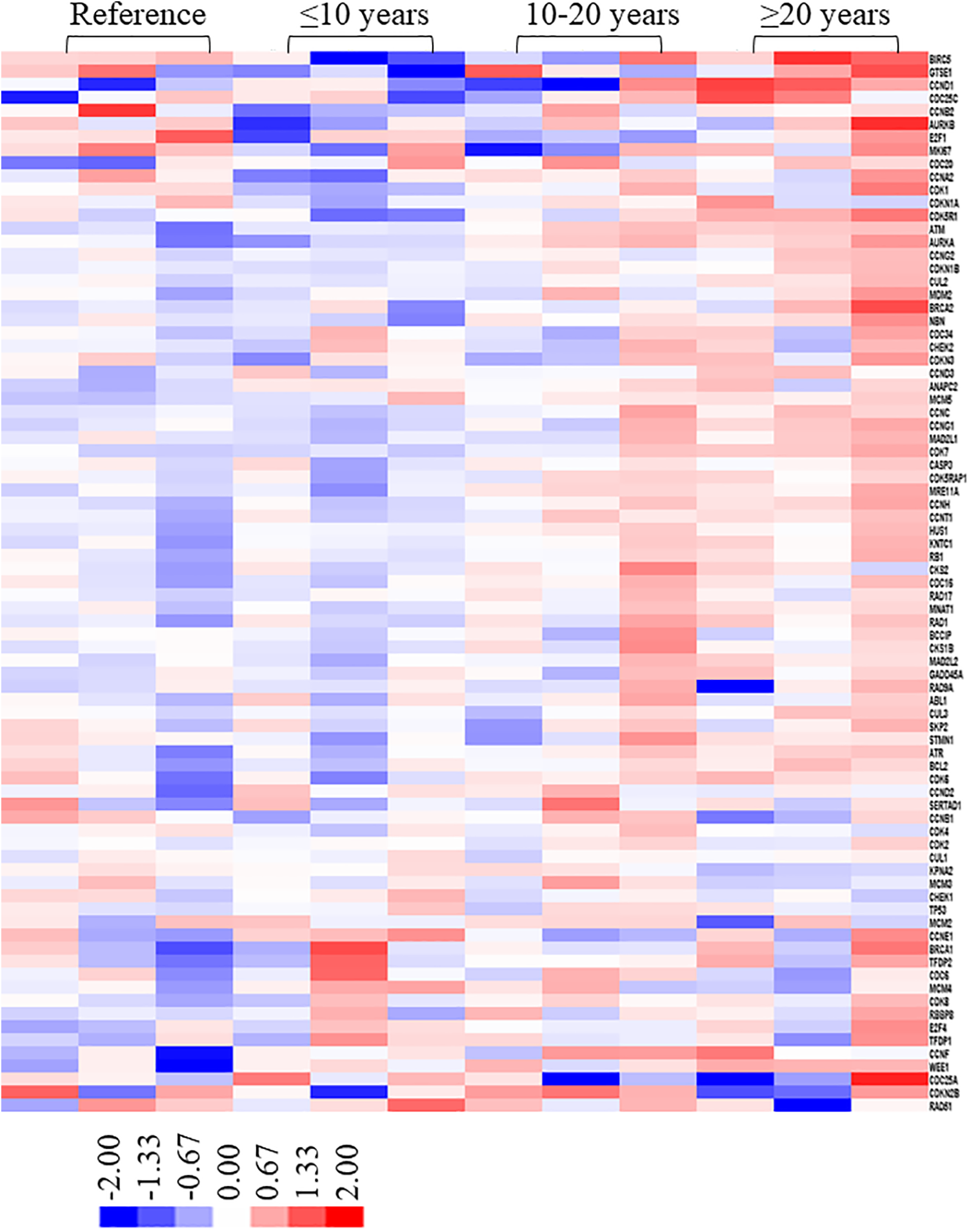
Identification of genes responsive to low-dose X-ray exposure using gene profiling. Heat map showing the log (fold-changes) in DNA damage-related genes in exposure and control groups as determined by PCR array analysis. Fold changes were calculated versus the control group average. Color scale ranges from red to blue, which denotes up- or downregulated genes, respectively. Red and blue color indicate genes with a > 2-fold upregulation and genes with a > 2-fold downregulation, respectively. (To better interpret the color references in the figure legend, please refer to the online version of this article.)
Biomarkers of Oxidative Stress
Sustained oxidative stress owing to chronic IR exposure results in DNA damage and a reduction in antioxidant enzyme activities. 24,25 In order to analyze the relationship between antioxidant enzyme activities and chronic low-dose IR, we analyzed the concentrations of MDA, GSH, and SOD in the serum of exposed and control groups. As shown in Figure 5A and B, MDA and SOD serum concentrations were not significantly different between individuals exposed to low-dose IR for less than 20 years and those who had no IR exposure. However, the MDA and SOD levels in workers who had been exposed to low-dose IR for more than 20 years were significantly lower than those of controls. There was no statistically significant relationship between GSH activity and the duration of low-dose IR (Figure 5C). Taken together, these results indicate that there is a relationship between serum MDA and SOD activities and the duration of low-dose IR exposure.

Malondialdehyde, SOD, and GSH levels in blood serum of the exposure and reference groups. (A) Malondialdehyde levels were significantly lower in the X-ray-exposed workers with more than 20 years of exposure. (B) Superoxide dismutase levels were significantly lower in X-ray-exposed workers with more than 20 years of exposure. (C) The relationship between the level of GSH and the X-ray exposure duration was not significant (*P < .05, **P < .01). GSH indicates glutathione; SOD, superoxide dismutase.
Discussion
The results of this study demonstrated the potential effects of chronic low-dose occupational IR exposure in medical workers from the Shandong region of China and subsequent genomic instability. Ionizing radiation is a potent genotoxic agent and human carcinogen, which induces cellular damage and gives rise to a variety of long-term effects, including genomic instability and carcinogenesis. 26,27 Ionizing radiation causes various types of DNA damage, of these DSBs are the most dangerous since they contribute to the formation of CA and compromise cell viability. 28 Ionizing radiation also induces changes in gene expression that are critical for a cell deciding between resuming normal functioning or undergoing apoptosis. 29 The increasing use of IR in medical diagnosis and treatment has raised concerns about its potential long-term effects on human health. 30 The results of our study sought to assess the association between IR and genomic stability in the peripheral blood of adults exposed to chronic low-dose IR.
In this study, we demonstrated that the level of DNA damage, CA, and CBMN frequency in individuals exposed to low-dose IR were all significantly higher when compared to unexposed controls. Our findings were in agreement with other studies examining Korean and Romanian workers exposed to occupational IR, where a higher MN frequency was reported after exposure to chronic low-doses of IR. 31 Additionally, we also conducted DNA damage-related gene expression profiling to reveal changes in gene expression levels following low-dose IR exposure. We concluded that the differently expressed genes were related to duration of IR exposure. All of the changed genes play a key role in maintaining genomic stability during the cell cycle as well as in apoptosis, cellular stress response and DNA repair. These genes have also exhibited impaired abilities to repair damaged DNA, which might be due to oxidative damage to nucleobases and changes to DNA methylation. 32 When compared with the reference group, our results indicated that men in the IR group had 18 up- and 7 downregulated genes. And the changed genes can be used as a potential biomarkers to evaluate the damage induced by IR. The changes in gene expression biomarkers (BIRC5, GTSE1, CCND1, CDC25C, E2F1, MKI67, CCNA2, CDK1, ATM, CDK5R1, BRCA2, CCNE1, BRCA1, TFDP2, CDC25A, CDKN2B, ATR and RB1) may be employed as more sensitive markers than cytogenetic markers (CA and micronucleus) to estimate the risks of low-dose radiation exposures. We found different genes involved in DNA repair, including BRCA1 and BRCA2. Moreover, ATM, CHEK1, and CDK1 are involved in both cell cycle regulation and DNA repair. 33,34 All the results suggest a complex signaling pathway activated by low-dose medical radiation exposure. On exposure to low doses of radiation (in workers who had been exposed to low-dose X-ray radiation for 10-20 years), the TP53 response pathway was triggered upregulation. Also, the DNA damage response pathways (ATM/CHK2) trigger upregulation in the group that had been exposed to low-dose X-ray radiation for more than 20 years, if there were very few DNA DSBs and they were repaired rapidly. And the complex change in regulators of gene expression may also contribute to differences in radiation sensitivity and response. Low-dose IR might disrupt the expression of key genes that take part in DNA repair pathways. The findings presented here may be helpful in understanding the mechanisms behind low-dose IR-mediated genome instability.
The levels of DNA damage, gene expression, CA, and CBMN frequency in individuals exposed to low-dose IR were higher in lymphocytes, indicating that peripheral blood components are sensitive to IR. This may be due to enzymatic antioxidants such as SOD, MDA, and GSH in blood serum that play an important role in scavenging ROS. 35 In the presence of molecular oxygen, IR converts hydroxyl, superoxide, and organic radicals into hydrogen peroxide and organic peroxides, which are longer-lived ROS that continue to damage the cellular genome and other important biomolecules. 36 Our results indicated that workers exposed to occupational IR for more than 20 years had decreased MDA and SOD activity; however, GSH level was not statistically different when compared with controls. These results indicate that the constant oxidative stress due to protracted IR exposure may have produced a greater amount of DNA damage in workers occupationally exposed to IR. Collectively, the direct oxidative damage to DNA and gene expression changes may lead to genomic instability in humans exposed to chronic low-dose IR.
Medical radiology workers exposed to low-levels of IR may be at a greater health disadvantage owing to the deleterious and long-term adverse health effects triggered by IR. Our study showed the level of DNA damage, CA, and CBMN frequency were all significantly increased. Moreover, the gene expression and antioxidant activities were changed in the peripheral blood of men exposed to low-dose X-rays. These results indicated a strong correlation between genomic instability and duration of exposure to low-dose IR. We also found that DNA damage repair and antioxidative changes were likely involved in the mechanisms of low-dose IR exposure. Given these results, further studies are required to elucidate the exact molecular mechanisms of genomic instability in health-care workers exposed to chronic low-dose IR, and it will be necessary to implement additional measures to minimize the occupational IR exposure of workers in medical diagnosis and treatment.
Supplemental Material
Supplementary_Materials - Assessment of Genomic Instability in Medical Workers Exposed to Chronic Low-Dose X-Rays in Northern China
Supplementary_Materials for Assessment of Genomic Instability in Medical Workers Exposed to Chronic Low-Dose X-Rays in Northern China by Lianying Fang, Jieqing Li, Weiguo Li, Xuesong Mao, Ya Ma, Dianjun Hou, Wei Zhu, Ximing Jia and Jianwei Qiao in Dose-Response
Footnotes
Authors’ Note
Lianying Fang and Jieqing Li conceived and designed the experiments; Lianying Fang, Weiguo Li, Ya Ma and Ximing Jia performed the experiments; Lianying Fang and Jieqing Li analyzed the data; Ya Ma, Ximing Jia, Wei Zhu, Dianjun Hou and Jianwei Qiao contributed reagents and materials; Lianying Fang wrote the paper.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Natural Science Foundation of Shandong (ZR2017YL007) and the Technical Development of Medical and Health of Shandong (2016WS0520).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.