
Abstract
A central question in radiation protection research is whether low-dose and low-dose-rate (LDR) exposures to ionizing radiation play a role in progression of cardiovascular disease. The response of endothelial cells to different LDR exposures may help estimate risk of cardiovascular disease by providing the biological mechanism involved. We investigated the effect of chronic LDR radiation on functional and molecular responses of human aorta endothelial cells (HAoECs). Human aorta endothelial cells were continuously irradiated at LDR (6 mGy/h) for 15 days and analyzed at time points when the cumulative dose reached 0.05, 0.5, 1.0, and 2.0 Gy. The same doses were administered acutely at high-dose rate (HDR; 1 Gy/min). The threshold for the loss of angiogenic capacity for both LDR and HDR radiations was between 0.5 and 1.0 Gy. At 2.0 Gy, angiogenic capacity returned to normal only for HAoEC exposed to LDR radiation, associated with increased expression of antioxidant and anti-inflammatory genes. Pre-LDR, but not pre-HDR, radiation, followed by a single acute 2.0 Gy challenge dose sustained the expression of antioxidant and anti-inflammatory genes and stimulated angiogenesis. Our results suggest that dose rate is important in cellular response and that a radioadaptive response is involved for a 2.0 Gy dose at LDR.
Introduction
The impact of occupational or environmental exposure to low-dose ionizing radiation and the risk of developing noncancerous effects, including cardiovascular disease, have not been completely clarified. Experimental data indicate that several parameters could be involved in the cellular response to ionizing radiation, such as dose/dose rate, time of exposure, and type of tissue, suggesting that low-dose radiation could stimulate different reactions than high-dose radiation. 1 An excess risk of developing circulatory disease after high-dose exposure was found in Japanese atomic bomb survivors (acute single dose) and in some occupational groups . 2,3 A possible biological mechanism proposed was that an inflammatory response and the level of reactive oxygen species (ROS) contribute to the injury of endothelial cells although it is unclear whether this assumption can be extrapolated to low-dose and low-dose-rate (LDR) exposure. 4,5 Based on current understanding, direct correlations between low doses and the development of cardiovascular disease are difficult to establish, and this is a high priority for radiation protection research. 6 Further studies are needed to determine whether low dose and LDR have an effect on vascular disease and to elucidate the mechanisms underlying the cellular response.
Stewart et al showed that a single high-dose exposure (14 Gy) accelerated the development of atherosclerotic lesions in ApoE−/− mice (predisposed to atherosclerosis) and led to development of inflammatory plaque. 7 Other in vivo studies report that exposure to LDR (1 mGy/min) ionizing radiation slowed the progression of atherosclerosis in ApoE−/− mice, reducing plaque lesions and inflammatory cytokines during early or late stages of the disease, while high-dose rate (150 mGy/min) ionizing radiation induced both protective and detrimental effects. 8,9 Those results and others suggest that different responses are induced at high doses than at low doses, and it seems that the biological effects of low-dose ionizing radiation are not linear with the dose. 10
The endothelium plays an important role in the regulation of the arterial wall, which maintains its anatomical and functional integrity by producing many biologically active substances involved in the control of vascular tone, cell proliferation, inflammation, and homeostasis. The secretion of nitric oxide (NO) generated by the endothelial isoform of NO synthase (eNOS), which is generally considered to be atheroprotective, leads to a better endothelial barrier function, to an anti-inflammatory phenotype, and to a quiescence of smooth muscle cells. 11 Antioxidant enzymes also play an important role maintaining low and moderate amounts of reactive oxygen species (ROS), having beneficial effects on several physiological processes including wound healing and tissue repair. 12 The diminished production or availability of NO increases vascular oxidative stress and leads to inflammatory responses. Secretion of proinflammatory molecules, including interleukin 6 (IL-6) and tumor necrosis factor α (TNF-α), triggers a cascade of activation within the vessel wall. In this context, prolonged exposure to vascular stressors supresses the endogenous capacity of endothelial cells to repair the lesion, the endothelium loses its integrity, progresses to senescence, and becomes dysfunctional. 13
Radiation injury can upset the homeostatic balance of healthy macro-vessels by inducing endothelial cell senescence, inflammation, and loss of capillary formation following exposure to high-dose radiation. 14 It has been shown that in vitro exposure to a single high dose (>2 Gy) can stimulate cell adhesion molecules and the expression of gene mediators of endothelial cell proliferation, migration, and angiogenesis (differentiation into capillary-like vessels) in healthy macro-vessels. 15,16 However, the biological effect of chronic low-dose radiation on endothelial cells is not fully understood. In vitro studies showed that chronic LDR delivered at 4.1 mGy/h had harmful effects on human umbilical vein endothelial cells (HUVECs; premature senescence, loss of capacity to form vascular networks, and secretion of pro-inflammatory molecules) when the total dose was >2 Gy, while a lower dose rate (1.4 mGy/h) did not induce damage even at a dose of 2 Gy. 17,18 Moreover, Rombout et al showed that a low dose (0.05 Gy) induced subtle increases in DNA double-strand breaks in HUVECs compared to high doses. 19 Also, Rödel et al showed that acute radiation doses in the range of 0.3 to 0.7 Gy induced anti-inflammatory effects on human endothelial cells (EA.Hy926) by stimulating protein and mitochondrial RNA (mRNA) levels of transforming growth factor-β 1 (TGF-β1), leading to downregulation of leukocyte adhesion . 20 Moreover, acute exposure of endothelial cells to low doses delivered at LDR or HDR stimulated expression of genes 21 and proteins 22 involved in the regulation of oxidative stress and antioxidative defense, suggesting a radioadaptive response to low-dose radiation. A radioadaptive response is an important cellular phenomenon triggered in general by priming very low-dose irradiation, triggering a “protective” effect against subsequent high-dose irradiation. Evidence suggests that the radioadaptive response depends on the activation of DNA repair and cell cycle regulation or activation of antioxidant enzymes due to the oxidative stress caused by ionizing radiation. The increased activities of these antioxidant enzymes lead to rapid scavenging of ROS and consequently less cell damage in adapted cells (human lymphoblasts). 23 It is therefore possible that the biological responses of endothelial cells to ionizing radiation are guided by different functional and molecular mechanisms, depending on the dose and the dose rate.
The purpose of this study is to report whether and how low-dose radiation at different dose rates has an impact on cardiovascular disease, using human aortic endothelial cells (HAoECs). To gain more biological insights and to study the effects of dose and dose rate, we evaluated whether functional and molecular changes occurred in the endothelial cells. The phenomenon of radioadaptation is employed by cells to protect against a subsequent high-dose irradiation (challenge dose). To verify this hypothesis, HAoECs preexposed to a total dose of 2.0 Gy at LDR or HDR were given a challenge dose of 2 Gy delivered at HDR. We checked whether continuous LDR radiation impacts the functional parameters of HAoECs after postirradiation with a challenge dose. More specifically, we evaluated whether the effects of low (0.05 and 0.5 Gy) to moderated (1 and 2 Gy) doses delivered at LDR showed a distinctly different pattern of response from those caused by HDR. In this study, we compared LDR radiation exposure with high-dose rate exposure for both low (0.05 and 0.5 Gy) and moderate (1 and 2 Gy) accumulated doses. Finally, we investigated the capacity of LDR to induce adaptive mechanisms that could protect the vessels from endothelial dysfunction.
Material and Methods
Cell Culture
Human aortic endothelial cells (cat. #c-12271) from PromoCell (Heidelberg, Germany) were cultured in 75-cm2 flasks containing specific cell growth media. Endothelial cell growth medium MV (EGM-MV; cat. #c-22020) was supplemented with fetal bovine serum (FBS, 5%), epidermal growth factor (10 ng/mL), heparin (90 µg/mL), hydrocortisone (1 µg/mL), penicillin (10 µg/mL), and (0.1 mg/mL) streptomycin. Cells were passaged every 3 to 5 days upon reaching ∼80% confluence and used from passages 5. Culture media were changed every 2 days.
Schematic of the experimental protocol. Irradiations of the cells (Irradiated acutely or chronically) starts at the same time point (at D0).

Radiation Exposure
Chronic ionizing radiation exposure
Human aorta endothelial cells were cultured with EGM-MV, in 25- and 75-cm2 flasks. Cells were placed in a cell culture incubator equipped with 60Co and exposed to chronic LDR radiation (6 mGy/h; 16 days [394 hours]). Cell samples were harvested for analysis at 4 time points when the cumulative dose reached 0.05 Gy (very low dose), 0.5 Gy (low dose), 1.0 Gy (moderate dose), and 2.0 Gy (moderate dose) after 1/2, 4, 8, and 16 days, respectively. Irradiation was carried out continuously except during the replacement of culture media and the subculturing of cells (2 hours every 4 days). Control cells were grown in an identical incubator without exposure to ionizing radiation and used at the same passage.
Acute ionizing radiation exposure
Human aorta endothelial cells cultivated with EGM-MV in 25- and 75-cm2 flasks were exposed to single doses of 0.05, 0.5, 1, and 2 Gy of γ ionizing radiation at an HDR (1 Gy/min). Cells were grown in a regular incubator, and samples were harvested for analyses at the same passage and at the same time points as those used for the chronic LDR radiation group (1, 4, 8, and 16 days).
Challenge dose exposure
For the adaptive response protocol, HAoECs previously exposed to 2 Gy at LDR or HDR were exposed to a supplementary single dose of 2 Gy delivered at HDR (1 Gy/min). Two hours after irradiation, HAoECs were harvested for analysis. Nonirradiated cells were used at the same passage.
Schematic of the experimental protocol
Irradiations of the cells (irradiated acutely or chronically) starts at the same time point (at D0).
In vitro tube formation assay
Basement membrane gels (Matrigel, 8 mg/mL) from Dominique Dutscher (Brumath, France) were allowed to polymerize for 30 minutes at 37°C. Irradiated HAoECs (105 cells/cm2) were detached, diluted in EGM-MV, and irradiated cells were counted with trypan blue and only viable cells were seeded at 105 cells/cm2 on the gel matrix on gel to form pseudotubes for 6 hours at 37°C with 5% CO2. Then, cells were fixed with 1.1% glutaraldehyde for 15 minutes, and the total length of the tube structure was quantified with Videomet software (Microvision Instruments, France). Thirty random macroscopic fields were counted (10× objective). The results are reported as the average of 3 different experiments in each condition. Nonirradiated cells were set as 100%.
Cell Proliferation
Human aorta endothelial cells were grown to 70% confluence in 25-cm2 flasks containing EGM-MV or media containing 2% FBS. After irradiation, cells were fixed with 4% paraformaldehyde for 15 minutes at 37°C and permeabilized with phosphate-buffered saline (PBS) 0.5% Triton for 10 minutes. After saturation with bovine serum albumin ( 3%), cells were incubated overnight at 4°C with mouse monoclonal anti-Ki-67 (1:50) from DAKO (Glostrup, Denmark). After washing in PBS 0.1% Tween, cells were incubated with a goat antimouse immunoglobulin G tagged with Alexa 488 (1:500) for 30 minutes at room temperature. After washing, 30 µL of mounting medium with 4',6-diamidino-2-phénylindole (DAPI) was added to the flasks, and images of immunofluorescence staining were photographed using an inverted microscope. The percentage of Ki-67-positive cells was calculated as a ratio of DAPI-positive cells. The results are reported as the average of 3 different experiments in each condition. Nonirradiated cells were set as 100%.
Real Time-Polymerase Chain Reaction
Total RNA was extracted from T-75cm2 flasks using Tri Reagent solution (Sigma-Aldrich LLC, St Louis, Missouri). RNA quality (260/280 nm) was determined using a Nanodrop ND 1000 spectrophotometer. One microgram of total RNA was synthesized to 20 µL complementary DNA (cDNA) using the high-capacity cDNA Reverse Transcription Kit from Applied Biosystems (Life Technologies, Saint Aubin, France) according to the manufacturer’s protocol. Quantitative polymerase chain reaction analysis was performed with a QuantStudio 12K Flex Real-Time PCR System (Life Technologies) using TaqMan 6 carboxyfluorescein-labeled probes and a standard thermal cycler protocol (50°C for 2 minutes before the first cycle, 95°C for 15 seconds, and 60°C for 1 minute repeated 45 times). Human TaqMan gene primers for eNOS, endothelin-1, vascular endothelial growth factor-B (VEGF-B), VEGF-receptor2 (VEGF-R2), TGF-β, copper/zinc-superoxide dismutase (Cu/Zn-SOD), manganese-superoxide dismutase (Mn-SOD), interleukin 6 (IL-6), and fibroblast growth factor 2 were from Applied Biosystems. Relative expression levels were calculated with glyceraldehyde-3-phosphate dehydrogenase and ribosomal protein, large, P0 (RPLP0) as internal control genes, and Ct mean was used to analyze the results. Nonirradiated cells were set as 1. Each experiment was performed in triplicate.
Statistical Analysis
Significant differences between mean values were identified by analysis of variance with the Tukey posttest for multiple comparisons and t test with unpaired posttest. The results are expressed as the mean ± standard error of the mean of 3 different experiments for the chronic LDR group and 2 different experiments for the acute HDR and challenge dose groups. P values <.05 were considered significant.
Results
Low-dose-rate radiation stimulates endothelial cell proliferation in a dose rate–dependent manner and modulates capillary-like tube formationWe first evaluated cell proliferation using Ki-67 staining, a marker of cell cycle. As shown in Figure 1, the proliferation of HAoECs was not stimulated with single doses of 0.05, 0.5, and 1.0 Gy delivered at HDR, but a significant inhibition of cell proliferation was observed with a dose of 2.0 Gy (day 16) compared to nonirradiated cells. On the other hand, a significant augmentation of Ki67-positive HAoECs exposed to continuous LDR radiation was detected at all accumulated doses compared to nonirradiated cells and to the HDR group (Figure 1). A peak of proliferation was observed at day 4, when the cumulative dose reached 0.5 Gy and a less but still significant increase was observed at days 8 (1.0 Gy) and 16 (2.0 Gy). These results show that continuous LDR ionizing radiation stimulates cell proliferation compared to HDR exposure, suggesting a dose rate–dependent process.We then evaluated the capacity of HAoECs to differentiate into capillary-like tube structures using a model of in vitro Matrigel plug. In Figure 2, we observe that capillary-like structures were not stimulated when HAoECs received a single HDR irradiation of 0.05 (day 1) or 0.5 Gy (day 4). Inversely, with single HDR radiation exposure of 1.0 Gy (day 8) and 2.0 Gy (day 16), we observed significant inhibition of tube formation in comparison to nonirradiated cells. In contrast, we observed a significant increase in tube formation in the Matrigel with cumulative LDR radiation of 0.05 and 0.5 Gy (evaluated after days 1 and 4 postexposure, respectively) compared to nonirradiated cells and the single HDR group. However, when the cumulative dose reached 1.0 Gy (day 8), capillary-like network formation was significantly reduced but returned to baseline levels with LDR radiation of 2.0 Gy (day 16) compared to nonirradiated cells. These results suggest that continuous LDR ionizing radiation induces a nonlinear proangiogenic response with a transitory effect and a threshold observed between 0.5 and 1.0 Gy for the impairment of angiogenic potential.
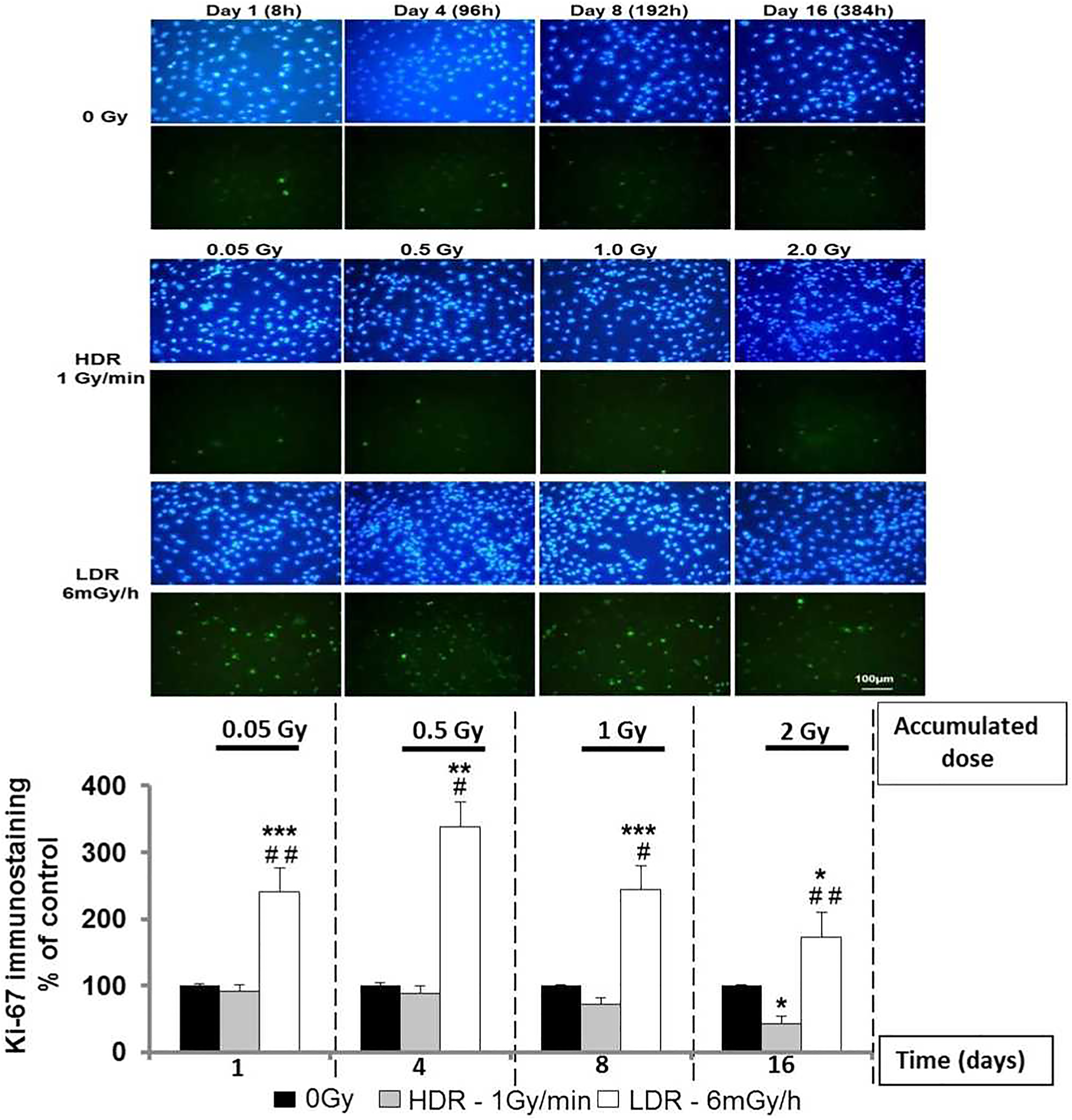
Chronic LDR radiation exposure stimulates cell proliferation in a dose rate–dependent manner. HAoECs (2 × 105) were seeded in 25-cm2 flasks, grown to 70% confluence, and irradiated at HDR (1 Gy/min) or LDR (6 mGy/h). Then, cells were immunofluorescent stained for Ki-67 (green) and counterstained with DAPI (blue). The number of Ki-67 positive cells/total number of cells (DAPI staining) was measured using 30 random fields/condition. Cell proliferation was quantified at days 1, 4, 8, and 16 of culture. Data are represented as a percentage of control (nonirradiated cells). Values are a mean ± SEM of 3 determinations. *P < .05; **P < .01; ***P < .001 versus no irradiated HAoECs at every time point. # P < .05; ## P < .01 versus the HDR group of cells. HAoEC indicates human aorta endothelial cells; HDR, high-dose rate; LDR, low-dose rate; SEM, standard error of the mean.
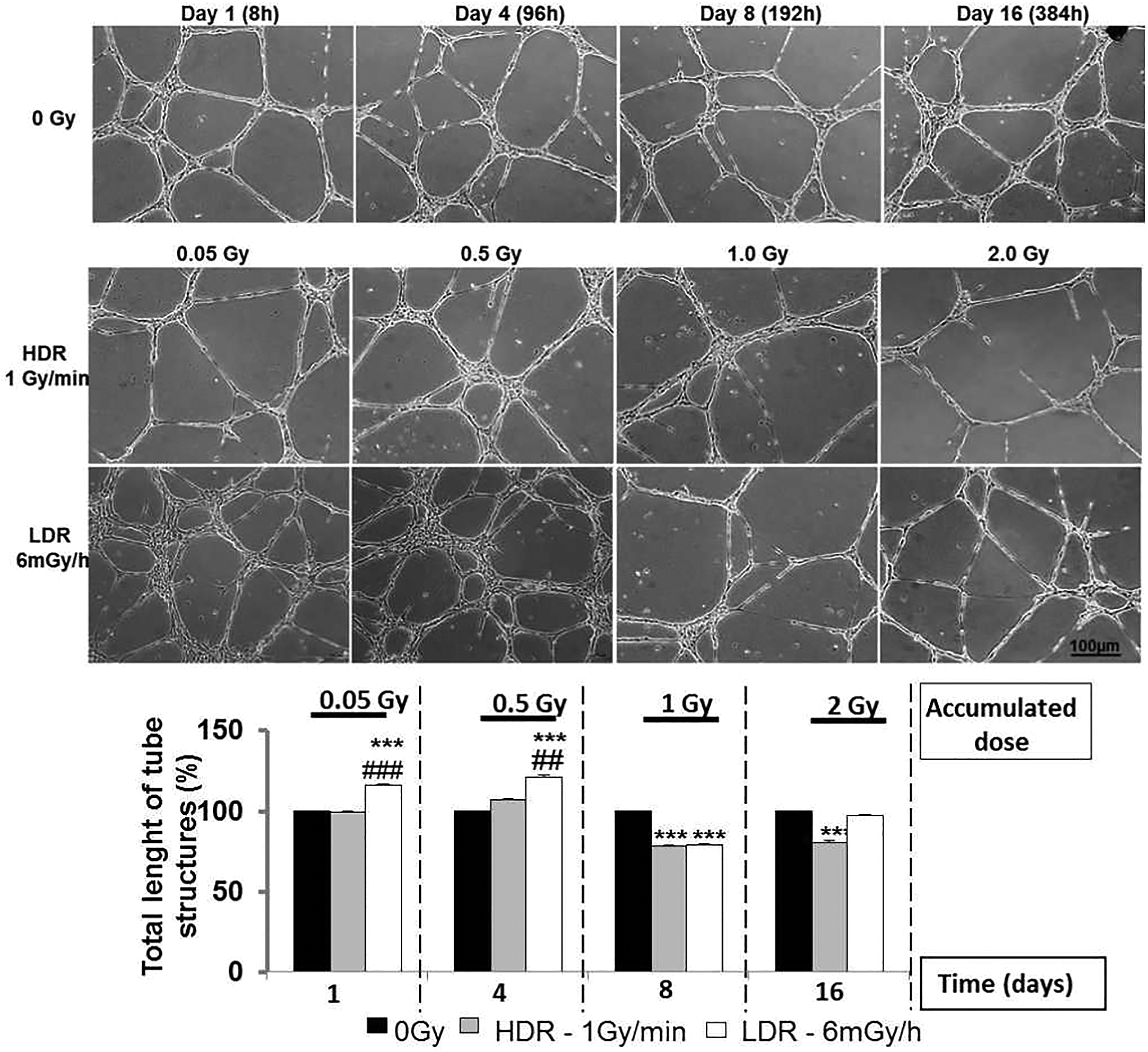
In vitro tube formation in Matrigel. HAoECs (105) irradiated at HDR (1 Gy/min) and LDR (6 mGy/h) were seeded on 24-well plates and quantified after 6 hours of differentiation. A, Morphological appearance of the Matrigel assay after 6 hours, for unirradiated (0 mGy) and irradiated HAoECs at HDR (1 Gy/min) or LDR (6 mGy/h). B, Quantification of tubular structures in Matrigel. 30 random fields/condition were measured. Tube formation was quantified at days 1, 4, 8, and 16 of culture. Data are represented as a percentage of control (unirradiated cells). Values are a mean ± SEM of 3 determinations. ***P < .001 versus nonirradiated HAoECs at every time point. ### P < .001 versus the HDR group of cells. HAoECs indicates human aorta endothelial cells; HDR, high-dose rate; LDR, low-dose rate; SEM, standard error of the mean.
Low-Dose-Rate Radiation Stimulates Antioxidant Genes in a Threshold-Dependent Manner
Using real time-polymerase chain reaction, we studied the genes involved in antioxidative stress. Figure 3A and B shows that exposure of HAoECs to single HDR radiation did not alter the gene expression of Cu/ZnSOD (SOD-1) and MnSOD (SOD-2) at any dose evaluated. Gene expression of catalase (CAT) was reduced after exposure to single HDR ionizing radiation of 0.5 (day 4), 1.0 (day 8), and 2.0 Gy (day 16) compared to nonirradiated cells (Figure 3C). Interestingly, continuous LDR radiation significantly upregulated all 3 genes after 8 days of exposure when cumulative dose reached 1.0 Gy. After 16 days of continuous LDR radiation, when the cumulative dose reached 2.0 Gy, the expression of SOD-1 mRNA levels was maintained upregulated, while gene expression of SOD-2 and CAT returned to basal levels.

Chronic LDR radiation induces gene expression of antioxidant enzymes in a dose-dependent manner. RT-PCR for SOD-1 (A), SOD-2 (B), and CAT (C) was performed in HAoECs irradiated at HDR (1 Gy/min) and LDR (6 mGy/h). RNA was extracted at days 1, 4, 8, and 16 of culture. Values are a mean ± SEM of 3 determinations. *P < .05 **P < .01; ***P < .001 versus nonirradiated HAoEC at every time point. # # P < .01 versus the HDR group of cells. HAoECs indicates human aorta endothelial cells; HDR, high-dose rate; LDR, low-dose rate; RT-PCR, real time-polymerase chain reaction; SEM, standard error of the mean.
Low-Dose-Rate Radiation Modulates Expression of Genes Involved in Angiogenic Processes in a Threshold-Dependent Manner
We evaluated the expression of pro-angiogenic genes. In Figure 4A and B, we observe that exposure of HAoECs to single HDR radiation stimulated neither VEGF nor VEGFR2 gene expression. Gene expression of VEGF-R2 was downregulated after a single HDR of 1.0 (day 8) and 2.0 Gy (day 16), compared to nonirradiated cells. Gene expression of VEGF was significantly increased when cumulative LDR reached 1.0 Gy (day 8), compared to nonirradiated HAoEC. When the cumulative dose reached 2.0 Gy (day 16), VEGF mRNA returned to normal levels. Gene expression of VEGF-R2 was significantly increased at a cumulative dose of 1.0 Gy (day 8) but sustained this pattern when LDR radiation continued for 16 days (2.0 Gy) compared to controls and to the HDR group.

Chronic LDR radiation induces the expression of genes involved in angiogenesis. RT-PCR for VEGF (A), VEGF-R2 (B), and eNOS (C) was performed in HAoECs irradiated at HDR (1 Gy/min) and LDR (6 mGy/h). RNA was extracted at days 1, 4, 8, and 16 of culture. Values are a mean ± SEM of 3 determinations. *P < .05; ***P < .001 versus nonirradiated HAoEC at every time point. # P < .05 versus the HDR group of cells. eNOS indicates endothelial isoform of NO synthase; CAT, catalase; HAoEC, human aorta endothelial cells; HDR, high-dose rate; LDR, low-dose rate; SEM, standard error of the mean; VEGF, vascular endothelial growth factor; VEGF-R2, vascular endothelial growth factor-receptor 2.
We next evaluated the gene expression of endothelial nitric oxide synthase (eNOS) in HAoECs. In Figure 4C, we observe that a single HDR radiation exposure significantly reduced eNOS mRNA expression at time points of 0.05 and 1 Gy (day 1 and 8) compared to nonirradiated cells, then sustained this profile with 2.0 Gy (day 16). After continuous LDR exposure of 0.05 and 0.5 Gy, we observed a significant upregulation of eNOS mRNA levels evaluated at days 1 and 4, respectively, compared to nonirradiated cells. Gene expression of eNOS returned to basal levels after 8 days (1.0 Gy) and sustained this profile after 16 days (2.0 Gy) of continuous LDR exposure. Together, these results suggest that continuous LDR ionizing radiation at low doses (0.05 and 0.5 Gy) does not have harmful effects on HaoEC but conversely seems to increase eNOS expression, which could contribute to vasoprotection at very LDR.
Low-Dose-Rate Radiation Counterbalances Genes Involved in Inflammation
We evaluated the regulation of inflammatory genes. In Figure 5A, we observe that a single HDR radiation exposure did not regulate gene expression of IL-6 in HAoECs, whereas a peak value, not significant, was observed with a single dose of 1.0 Gy (day 8). Interestingly, the expression of IL-6 was significantly downregulated when continuous LDR exposure reached 0.05 (day 1) and 0.5 Gy (day 4) compared to nonirradiated cells. When the cumulative dose reached 1.0 Gy (day 8), IL-6 mRNA was significantly upregulated, and then IL-6 was downregulated when LDR radiation continued for 16 days (2.0 Gy). In Figure 5B, we observe that the gene expression of TGF-β1 was regulated with a single HDR of 1.0 Gy (day 8), but a significant increase was only observed at 2.0 Gy (day 16) compared to nonirradiated cells. A significant upregulation of TGF-β1 mRNA was observed when continuous LDR radiation reached 0.5 (day 4), 1.0 (day 8), and 2.0 Gy (day 16), compared to nonirradiated cells. Together, these data suggest that continuous LDR exposure counterbalances pro- and anti-inflammatory genes between 0.5 and 1.0 Gy.

Chronic LDR radiation counterbalances pro- and anti-inflammatory factors. RT-PCR for IL-6 (A) and TGF-β1 (B) was performed in HAoEC irradiated at HDR (1 Gy/min) and LDR (6 mGy/h). RNA was extracted at days 1, 4, 8, and 16 of culture. Values are a mean ± SEM of 3 determinations. *P < .05; ***P < .001 versus nonirradiated HAoEC at every time point. # P < .05 versus the HDR group of cells. HAoEC indicates human aorta endothelial cells; HDR, high-dose rate; LDR, low-dose rate; RT-PCR, real time-polymerase chain reaction.
Low-Dose-Rate Radiation, But Not Acute HDR, Induces an Adaptive Response in Endothelial Cells: Challenge Dose Experiments
Based on the results showing that continuous LDR ionizing radiation, but not HDR exposure, stimulated endogenous protective genes (SOD-1, VEGF, and TGFb1) when the total dose reached 2.0 Gy, we wondered whether HAoECs could adapt to chronic ionizing radiation exposure. In Figure 6A, we observe that challenge dose exposure stabilized cell proliferation of HAoECs that were preirradiated with a cumulative dose of 2.0 Gy at LDR. However, when HAoECs were preirradiated with single HDR dose of 2.0 Gy, cell proliferation was inhibited. We then evaluated the effects of a challenge dose on capillary-like tube formation. In Figure 6B, we observed a significant increase in capillary-like tube formation when HAoECs first received a cumulative radiation dose of 2.0 Gy at LDR and were postirradiated with a challenge dose. In contrast, tube formation was not stimulated when HAoECs were irradiated early with a single 2.0 Gy dose at HDR and were postirradiated with a challenge dose. Thus, it seems that continuous LDR radiation stimulated an inherent angiogenic response of endothelial cells, while single HDR exposure induced harmful effects.
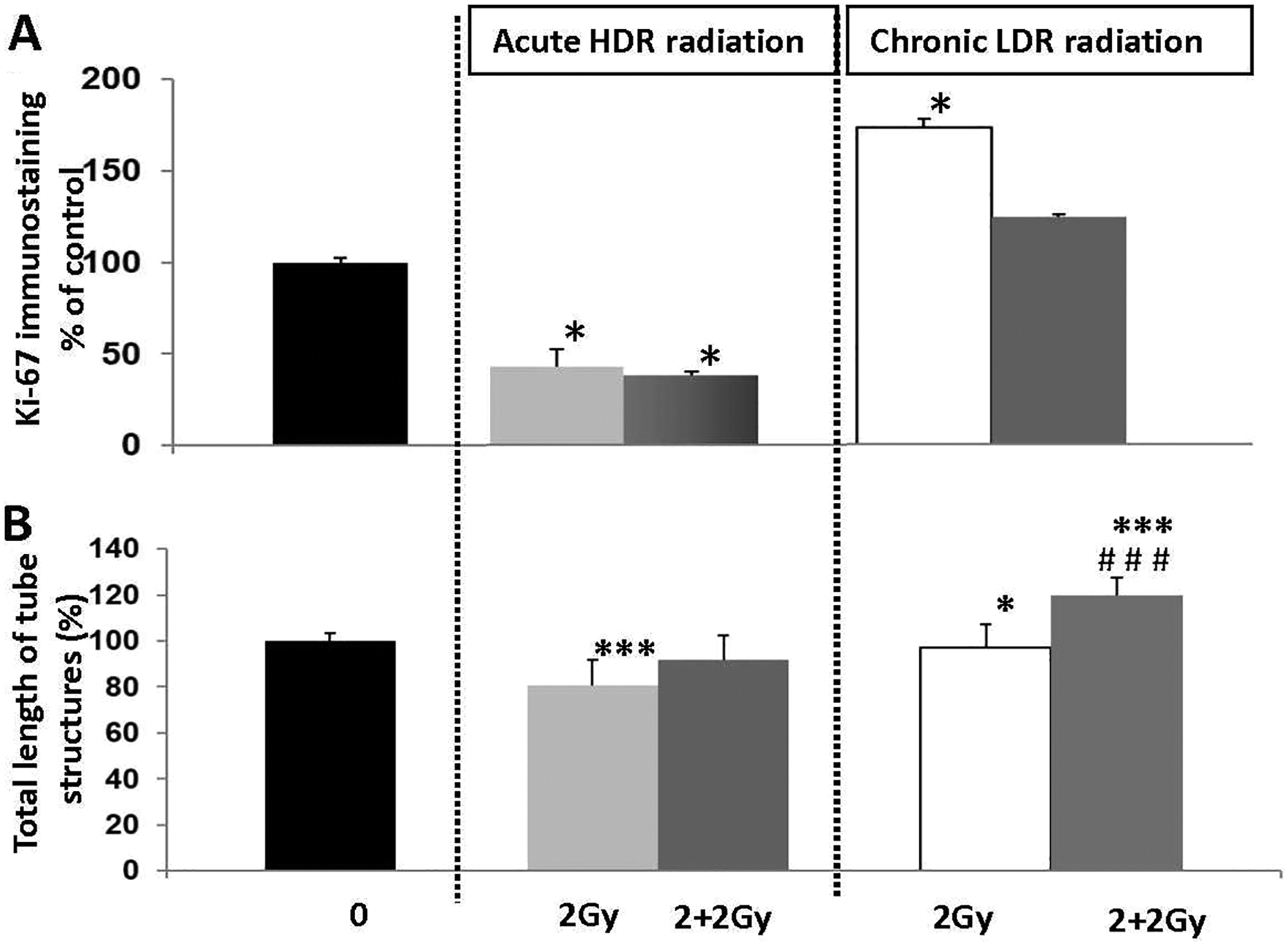
LDR, but not HDR radiation, stimulates functional parameters in HAoECs after a challenge dose. A, Cell proliferation. HAoEC (2x105) were seeded in 25-cm2 flasks, grown to 70% confluence, pre-irradiated at HDR (1 Gy/min) or LDR (6 mGy/h), and exposed for a second time to a challenge dose (2.0 Gy delivered at an HDR of 1 Gy/min). Human aorta endothelial cells were immunofluorescently stained for Ki-67 (green) and counterstained with DAPI (blue). The number of Ki-67positive cells/total number of cells (DAPI staining) was measured using 30 random fields/condition. Cell proliferation was quantified at day 16 of culture. B, In vitro tube formation in Matrigel. HAoECs (105) preirradiated at HDR (1 Gy/min) and LDR (6 mGy/h) were exposed for a second time to a challenge dose (2.0 Gy delivered at an HDR of 1 Gy/min), seeded on 24-well plates and quantified after 6 hours of differentiation. Thirty random fields/condition were measured. Cell proliferation and tube formation were quantified at day 16 of culture. Data are represented as a percentage of control (unirradiated cells). Values are a mean ± SEM of 3 determinations. *P < .05; ***P < .001 versus nonirradiated HAoEC at every time point. ### P < .001 versus LDR group that received a challenge dose of 2 Gy. HAoECs indicates human aorta endothelial cells; HDR, high-dose rate; IL-6, interleukin 6; LDR, low-dose rate; SEM, standard error of the mean.
Second, we verified whether a challenge dose would be able to regulate genes involved in vessel protection (SOD-1, SOD-2, and eNOS), endothelial cell repair (VEGF-2 and VEGF-R2), and inflammation (TGF-β and IL-6) after continuous LDR radiation. Figure 7A and B shows that a challenge dose of 2.0 Gy induced a weak but significant increase in gene expression of both SOD-1 and SOD-2 in comparison to HAoECs only preirradiated continuously at LDR at the time point of 2.0 Gy. In contrast, challenge dose exposure inhibited both SOD expressions when HAoECs were preexposed to a single dose of 2.0 Gy at HDR. Likewise, gene expression of VEGF-2 and VEGF-R2 underwent a small but significant increase following a challenge dose of 2.0 Gy after continuous LDR irradiation of 2.0 Gy. Both VEGF-2 and VEGF-R2 mRNA levels were downregulated when HAoECs were first exposed to acute HDR ionizing radiation of 2.0 Gy and then exposed to a challenge dose (Figure 7C and D). Notably, the challenge dose strongly upregulated TGF-β1 mRNA levels (4-fold) when HAoECs were continuously preirradiated at LDR, compared to the time point of 2.0 Gy, when cells were only irradiated with a continuous dose of 2.0 Gy at LDR. Inversely, a challenge dose significantly downregulated TGF-β1 mRNA levels when HAoECs were preirradiated with a single HDR dose, compared to the time point of 2.0 Gy (Figure 7E). Finally, the expression of eNOS and IL-6 mRNA levels was not modified after exposure to a challenge dose in HAoECs primed with continuous LDR or with acute HDR ionizing radiation (Figure 7F and G). Altogether, these findings suggest that chronic LDR ionizing radiation but not acute HDR exposure stimulates functional and molecular responses in HAoECs after a challenge dose.

Low-dose rate, but not HDR radiation, stimulates genes involved in vascular protection after a challenge dose. HAoECs preirradiated at HDR (1 Gy/min) or LDR (6 mGy/h) were exposed for a second time to a challenge dose (2.0 Gy delivered at an HDR of 1 Gy/min). RT-PCR for SOD-1 (A), SOD-2 (B), VEGF (C), VEGF-R2 (D), eNOS (E), IL-6 (F), and TGF-β1 (G) was performed in HAoECs irradiated at HDR (1 Gy/min) and LDR (6 mGy/h). RNA was extracted at day 16 of culture. Values are a mean ± SEM of 3 determinations. *P < .05; **P < .01***P < .001 versus nonirradiated HAoECs at every time point. ### P < .001 versus the HDR group of cells irradiated with 2 Gy. £ P < .05; ££ P < .01; £££ P < .001 versus the LDR group irradiated with 2 Gy. HAoECs indicates human aorta endothelial cells; HDR, high-dose rate; IL-6, interleukin 6; LDR, low-dose rate; TGF-β1, transforming growth factor-β1; SOD, superoxide dismutase; VEGF, vascular endothelial growth factor; VEGF-R2, vascular endothelial growth factor receptor 2.
Discussion
Although epidemiological studies have yielded insight into the relation of radiation exposure to cardiovascular risk, there are still many uncertainties. These include whether chronic exposure to ionizing radiation has an impact on cardiovascular disease incidence or progression; whether there is a threshold dose; and whether low dose/LDR has a potential protective effect against a subsequent high-dose irradiation, which characterizes the phenomenon of the radioadaptive response. There is therefore a need to improve the evidence available on which to base low-dose/LDR radiation risk assessment through cellular and molecular studies, so as to understand the mechanisms involved in vascular tissue. 24
We demonstrate here that dose rate plays an important role when estimating the relation between exposure to low-dose ionizing radiation and the risk of cardiovascular disease. The most relevant results of our in vitro study are that: (1) a threshold (between 0.5 and 1.0 Gy) for the anti-angiogenic effect on endothelial cells is dose dependent for both LDR and HDR exposure; (2) exposure to LDR ionizing radiation, but not HDR exposure, induces an upregulation of genes involved in vascular integrity and restores the ability of HAoECs to differentiate into vascular networks (return to normal levels) when the cumulated dose reaches 2.0 Gy (day 16), and (3) postexposure to a challenge dose upregulates genes involved in antioxidative and anti-inflammatory responses only in HAoECs preexposed to LDR, while preexposure to HDR radiation followed by a challenge dose induces the downregulation of all genes evaluated.
Also, only LDR exposure stimulated cell proliferation at every time point, suggesting a dose rate dependency. Moreover, a single acute 2.0 Gy challenge dose sustained the expression of antioxidant and anti-inflammatory genes and induced a better functional proangiogenic response of HAoECs after pre-LDR chronic radiation exposure but not after pre-HDR acute exposure. These results suggest that chronic LDR exposure induces a radioadaptive response when the chronic dose reached 2.0 Gy. Together, our results suggest that dose rate is an important parameter in estimating the cellular response after exposure to chronic and acute ionizing radiation. Furthermore, for the first time our study shows an adaptive response in endothelial cells for a chronic 2.0 Gy dose.
It has been shown that, depending on dose and dose rate, ionizing radiation induces stress and changes the redox environment by eliciting transient responses at the molecular, cellular, and tissue levels to counteract the toxic effects of radiation. 22 Notably, ROS produced by low-dose γ radiation have biological effects similar to those generated by normal oxidative mechanisms. 25 Reactive oxygen species act on endothelial cells through tightly regulated mechanisms that are dependent on the amount and the site of production of ROS as well as on the balance of pro-oxidant and antioxidant enzyme activity. 26 Reactive oxygen species act as a double-edged sword in the vasculature because excess amounts (oxidative stress) contribute to various diseases, including atherosclerosis and diabetes, whereas transient or low levels of ROS promote physiological angiogenesis and homeostatic maintenance of healthy vasculature in response to ischemia and wound healing. 27 Superoxide dismutases are the major ROS detoxing enzymes protecting the cells from potential damage caused by excessive amounts of ROS. 1,28 In vitro and in vivo studies demonstrate that exposure to chronic LDR ionizing radiation upregulates gene expression of antioxidant defenses (eg, glutathione and SOD). By contrast, HDR propagates oxidative stress associated with decreases in activity of MnSOD, CuZnSOD, catalase, and glutathione peroxidase. 29,30 Also, Large et al showed that low doses between 0.3 and 3 Gy stimulate nonlinear expression of antioxidative systems in irradiated transformed human umbilical vein endothelial cells human umbilical vein endothelial cells, suggesting a contribution to anti-inflammatory effects in stimulated endothelial cells. 31 We observed upregulation of MnSOD (SOD-2), CuZnSOD (SOD-1), and catalase after exposure to continuous LDR but not after HDR ionizing radiation. It is interesting to note that endothelial cells could regulate these antioxidant genes between 0.5 and 1 Gy as when the antiangiogenic process was observed. In this way, it could be possible that the antiangiogenic response observed when chronic LDR ionizing radiation reached 0.5 Gy in endothelial cells occurred in a response to an excess of ROS, possibly balanced by upregulation of antioxidants when the dose at LDR reached 1.0 Gy, allowing the restoration of function and the maintenance of low levels of ROS. Inversely, acute HDR ionizing radiation did not induce redox balance in endothelial cells. More studies are needed to understand the role of the ROS/SODs balance in response to low-dose radiation and LDR radiation.
The roles of VEGF and VEGFR2 as endogenous modulators of key steps of angiogenesis, including cell proliferation and cell differentiation into new vessels, 32 are well known. It has been postulated that ionizing radiation increases the activity of VEGF and VEGFR2 in endothelium, resulting in protection from cell death. 33 Kermani et al demonstrated that even after exposure to a high dose of 10 Gy, the antiangiogenic effect of ionizing radiation on human coronary artery endothelial cells was correlated with upregulation of VEGFR2. This inhibitory response was reversed after the treatment of endothelial cells with VEGF165 (10 ng/mL). Kermani et al 34 suggest that the radiation-induced upregulation of VEGFR2 expression in endothelial cells, correlated with the rescued functional response after the treatment with VEGF165, seems to be important in the survival of radiation-damaged endothelium and regulation of reendothelization in blood vessels in vivo, playing a radioprotective role. In agreement with these findings, our results show that gene expression of both VEGF and VEGFR2 was upregulated after exposure to a dose of 1.0 Gy delivered at LDR and that VEGFR2 gene expression was also increased after exposure to 2.0 Gy, suggesting that a compensatory response between doses of 0.5 and 1.0 Gy may have been stimulated in endothelial cells to impair the harmful effects of chronic LDR ionizing radiation. However, in our experiments, it is important to take into account the fact that since cells were subcultured during chronic irradiation exposures, particularly for the higher accumulated doses with longer culturing periods, some of the effects might have been additive and transgenerational in nature and the cumulative dose should be lower than 1 and 2 Gy because of cell turn over. However, with these type of in vitro mechanistic studies of chronic exposure, it is hard to exclude cell turn over and the fact that cells received lower doses. Therefore, we could consider that the newly divided cells are generated from irradiated ones. Furthermore, even with in vivo chronic low-dose irradiation exposure experiments one cannot exclude cell turn over in the body.
It is well known that in vascular disease, inflammation is the driving force, 35 and strong evidence supports the central role of pro-inflammatory cytokines such as IL-6 and TNF-α. Endothelial cells play a crucial role in the regulation of the local inflammatory process both by their ability to recruit leukocytes from peripheral blood and to express a variety of cytokines/chemokines and growth factors. 36 As a consequence, the effect of low-dose irradiation on the adhesion process in in vitro assays using human EA.Hy926 (endothelial hybrid cells the immortalize derivative of human umbilical vein endothelial cells) had biphasic kinetics and involved elevated expression of the anti-inflammatory cytokine TGF-β1, both as mRNA and protein, following exposure to 0.3 to 0.5Gy. 37 This indicates that TGF-β1 is a key player in the modulation of adhesion following low-dose exposure. We observed an increase in both TGF-β1 and IL-6 gene expression when the cumulated dose reached 1.0 Gy after LDR irradiation. However, after continuous LDR exposure, the level of IL-6 mRNA was reduced, while the gene expression of TGF-β1 was maintained upregulated when LDR exposure reached 2.0 Gy. The fact that LDR ionizing radiation stimulates anti-inflammatory proteins and inhibits pro-inflammatory molecules in endothelial cells could be understood as a mechanism to protect from injury, but more studies are needed to clarify the role of inflammation in low-dose exposure and LDR exposure. The implications of the anti-inflammatory effect of low-dose irradiation are well documented. Several experimental findings were reviewed recently and showed that low-dose radiation induces anti-inflammatory properties and may thereby protect against inflammatory disease. 38,39 Chronic LDR radiation of different wild-type mouse strains may have stimulating effects that activate the immunological network of the whole body, including cell populations and their surface molecules, together with antibody-producing activity. 40 Low doses administered at LDR are anti-inflammatory and decrease atherosclerosis in most situations. 8,41,42 Recent studies demonstrate the importance of dose rate in the inflammatory response. Indeed, HDR is associated with an upregulation of inflammation, whereas LDR induces an anti-inflammatory response. 18,43 In the latter study, chronic low-dose 137Cs exposure for 6 months reduced the inflammatory mediators CRP, TNFα, MCP-1, IFN-γ, and plaque macrophage content, suggesting a potential increase in stability.
Little is known about the proteins and genes involved in the adaptive response in endothelial cells after exposure to low-dose radiation. The genes involved in antioxidant defense, such as MnSOD, CAT, Cu/ZnSOD, and GPXI, are upregulated after low-dose ionizing radiation and induce a radioadaptive response, with enhanced cellular homeostasis and repair capacity against subsequent similar high-dose radiation. 21,44 We have shown that when cells were pretreated at LDR, even when the dose reached 2 Gy, we observed a better functional and molecular cellular response after a challenge dose compared to cells pretreated at HDR. Indeed, the pro-angiogenic effect was upregulated after the challenge dose when cells were pretreated at LDR compared to cells that did not receive a challenge dose. On the contrary, when the cells were pretreated at HDR we did not observe an increase in pro-angiogenic activity. Radioadaptive responses have been observed in vitro and in vivo using various indicators of cellular damage, such as cell lethality, chromosomal aberrations, mutation induction, radiosensitivity, and DNA repair. 45 –49 Indeed, adaptation has been shown in response to both low linear energy transfer (LET) (X-rays, γ-rays, β-particles) 50 –52 and high LET (neutrons, α-particles) radiation. 48,53 A combination of a low LET (X-rays) priming dose and a high LET (α-particles of radon gas) challenge dose leads to adaptation in human lymphocytes. 54,55 In cellular studies, values of priming doses and dose rates resulting in adaptive responses seem to be located within certain ranges. 46,51,56,57,58 Consequently, no adaptive response is seen with priming doses or dose rates outside this window. In general, in experimental set ups resulting in adaptive behavior, the values range from 0.01 to 0.5 Gy and from 0.01 to 1.0 Gy/min for priming dose and priming dose rate, respectively. Moreover, adaptive responses can be induced in cells not receiving a preconditioning dose, the evolution of nontargeted effect radiobiology meant that at low doses the previous assumptions needed to be reconsidered on the light of the existence of non-DNA mechanisms. 59,60
We have shown that even at the medium dose of 2 Gy, there was an adaptive response only when 2 Gy was administered at LDR. In experiments in numerous patients, adaptation was observed in 50% to 78% of cases, and its magnitude ranged from 11% to 32% reduction in the measured effects. 61,62 Variability has been shown to depend on factors such as dose rate and time between doses. It was also shown in animal experiments that the same dose given at LDR can be beneficial, whereas at HDR, it is detrimental in the progression of atherosclerotic plaques in ApoE−/− mice. 8
At the cellular level, SOD2 plays a key role in providing the radioadaptive effect by reducing the amount of toxic superoxide formed after radiation. It catalyzes the conversion of 2 molecules of superoxide anion into water and hydrogen peroxide, and the latter can be further oxidized to water. SOD2 loss or deficiency sensitizes cells to ionizing radiation, whereas reinstating SOD2 in SOD-2-deficient cells restored the radioadaptive phenotype, indicating that SOD2 is important in protecting cells against radiation via the reduction in mitochondrial damage caused by radiation-produced free radicals, which may lead to cellular apoptosis (Grdina Dj, 2013; Iyer R, 2002 (Rad res); Iyer R, 2002 (Mut Res); Murley JS, 2011). 63,64,65,66 The radioadaptive response that we observed may be mediated by the SOD2 signaling pathway.
In this study, we have shown that LDR radiation induces an adaptive molecular and functional response in HAoECs. Even at a dose of 2 Gy administered at LDR, we observed an adaptive response expressed by better proangiogenic function and molecular antioxidative response. Further studies are required, for example, with a coculture of endothelial and smooth muscle cells to identify the dialogue between these cells during low-dose chronic or acute exposure and to identify signal transduction pathways involved after LDR-induced responses.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Electricité de France (EDF).