
Abstract
Increased oxidative stress plays an important role in heavy ion radiation–induced cell death. The mechanism involved in the generation of elevated reactive oxygen species (ROS) is not fully illustrated. Here we show that NADPH oxidase activation is closely related to heavy ion radiation–induced cell death via excessive ROS generation. Cell death and cellular ROS can be greatly reduced in irradiated cancer cells with the preincubation of diphenyleneiodium, an inhibitor of NADPH oxidase. Most of the NADPH oxidase (NOX) family proteins (NOX1, NOX2, NOX3, NOX4, and NOX5) showed increased expression after heavy ion irradiation. Meanwhile, the cytoplasmic subunit p47phox was translocated to the cell membrane and localized with NOX2 to form reactive NADPH oxidase. Our data suggest for the first time that ROS generation, as mediated by NADPH oxidase activation, could be an important contributor to heavy ion irradiation–induced cell death.
Introduction
Reactive oxygen species (ROS) level is elevated when ionizing radiation interacts with water molecules, which produces additional ROS via radiolysis of water molecules. To prevent excessive generation of ROS during radiation exposure, addition of antioxidants has been shown to protect cells from the harmful effects of ionizing radiation both in vitro and in vivo. 1 –3 However, the sources of cellular endogenous ROS after radiation exposure remain unclear. Several enzymes contribute to the ROS generation in vivo, including NADPH oxidase, cyclooxygenases, lipoxygenases, myeloperoxidase, nitric oxide synthase, and xanthine oxidase. Among them, NADPH oxidase can be stimulated to generate ROS within a few minutes by various growth factors, such as cytokines and hormones. 4 Environmental stress factors such as nutrient stress, heavy metals, and ultraviolet radiation could activate NADPH oxidase to produce ROS. 5 –8 Several studies showed that low linear energy transfer (LET) radiation induced cellular damage via excessive ROS generation, which was produced by active NADPH oxidase. 9 –11 However, whether heavy ion irradiation could activate NADPH oxidase remains elusive.
NADPH oxidase was originally identified in phagocytes as the major source of ROS. NADPH oxidase complexes consist of 7 homologs, including NOX1 through NOX5, DUOX1, and DUOX2. The NADPH oxidase complexes transfer an electron from NADPH to molecular oxygen, producing superoxide anion, which is a precursor for other reactive oxygen and nitrogen species. 3 It is now clear that it is functionally expressed in many other cells such as fibroblasts, osteoclasts, smooth muscle cells, and neurocytes. 12,13
Here the role of NADPH oxidase in carbon ion irradiation–induced cancer cell death was investigated. Results showed that increased ROS generation, as produced by active NADPH oxidase, contributed to heavy ion irradiation–induced cell death. Carbon ion irradiation induced elevated expression of NOX1 through NOX5, and the translocation of p47phox subunit to the membrane was observed. This is the first report showing that NADPH oxidase–generated ROS is involved in heavy ion irradiation–induced cell death, which has potential importance for the combination of gene transfer and heavy ion irradiation in cancer therapy.
Methods
Cell Culture and Irradiation Treatment
The human liver cancer cell line HepG2, lung cancer cell line GLC-82, and cervix cancer cell line HeLa were purchased from the American Type Culture Collection (Manassas, USA). Cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum. Cells were cultured in 5% CO2 in humidified air at 37°C.
Carbon ion irradiations were performed at room temperature at the Heavy Ion Research Facility in Lanzhou of the Institute of Modern Physics, Chinese Academy of Sciences, with 300 MeV/n carbon ion; the LET value for carbon ion was 40 keV/μm. The dose rates were 1 Gy/min.
Reactive Oxygen Species Measurement
Reactive oxygen species measurement for whole cell and isolated mitochondria was carried out according to standard procedure. 14 Briefly, HeLa cells were harvested in an exponential growth phase and incubated at 37°C in 96-well plate in 300 µL DMEM medium for 6 hours. At indicated time points after treatments, each 96-well plate was incubated with 10 µM 2′,7′–dichlorofluorescin diacetate (DCFDA) for 30 minutes in the dark. Using excitation at 485 nm and emission at 525 nm, fluorescence was then recorded at each time points. For isolated mitochondria, each well of a 96-well plate was filled with phosphate-buffered saline (PBS) buffer, containing 10 μM of DCFDA and 0.5 mg/mL of submitochondrial particles, to a final volume of 200 μL for 30 minutes at room temperature. The recording procedure then was followed as described above.
Cell Survival Assay
HeLa cells were seeded in 96-well plates at a density of 5 × 104 cells in 200 µL of medium per well. On the next day, the medium was changed to fresh complete DMEM with or without 10 µM diphenyleneiodium (DPI) and the cells were incubated at 37°C for 10 minutes, followed by washing 5 times with PBS, then 200 µL of DMEM containing 10% fetal calf serum (FSC) was added to each well. At 48 hours after irradiation, the medium was removed and 0.5 mg/mL 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyl-2 H-tetrazolium bromide (MTT) was added followed by incubation at 37°C for 4 hours in a CO2 incubator. The MTT formazan product was released from cells by addition of 150 µL sodium dodecyl sulfate (SDS) with final concentration of 10% (vol/vol) per well. After incubation overnight at 37°C, the crystals were completely dissolved and absorbance at 620 nm (A620) was measured using a microplate spectrophotometer reader (Multiskan Ascent, Labsystems, Finland), and A620 was used to estimate the cell viability.
Western Blotting
All samples were mixed with loading buffer and subjected to 10% SDS-polyacrylamide gel electrophoresis. Each lane was loaded with 50 to 100 μg of equal amounts of protein. All samples were then transferred onto polyvinylidene fluoride (PVDF) membranes and blocked with 5% milk in Tris-buffered saline–Tween 20 (TBST) buffer for 1 hour at room temperature. The membranes were subsequently exposed to first antibody against NOX1, NOX2, NOX3, NOX4, NOX5, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Abcam Inc, Cambridge, MA) at the dilution of 1:1000 in 5% milk in TBST for overnight at 4°C and then membranes were washed and incubated with secondary antibody. Antibody binding was detected using enhanced chemiluminescence (BioRad, California, USA). The results were quantified by densitometry using Image J (version 1.43b) software.
Immunofluorescence Staining and Confocal Microscopy
Cancer cells were grown on coverslips in 12-well plates. After irradiation, cells were fixed with 1% formaldehyde in PBS at room temperature for 30 minutes and then permeabilized in 0.1% Triton X-100 in PBS for 10 minutes. Cells were incubated at 37°C for 2 hours with NOX2 (Abcam Inc, Cambridge, MA) and anti-p47phox (C-20; Santa Cruz Biotechnology, Texas, USA) goat polyclonal antibody, at a 1:500 dilution in PBS containing 10% bovine serum albumin (BSA) and 0.1% Tween 20. Cells were then stained with fluorescein isothiocyanate–conjugated rabbit antigoat immunoglobulin G (IgG) (81-1611; Zymed, Wien) or Cy-5-conjugated goat antirabbit IgG (81-6116; Zymed, Wien), at a ratio of 1:100 in PBS containing 10% BSA and 0.1% Tween 20 at 37°C for 1 hour. Following extensive washing in PBS, the cells were mounted on slides using a 4′,6-diamidino-2-phenylindole (DAPI) (H-1200; Vector Lab, California, USA) mounting medium. The stained cells were observed under a confocal laser scanning microscope (Zeiss LSM 510 Meta, Oberkochen, Germany).
Statistical Analysis
Statistical analysis was performed on the means of the data obtained from at least 3 independent experiments. Data are presented as means (standard deviation [SD]). Student t test program in Microsoft Excel was used to detect statistical significance. P < .05 was considered to be statistically significant.
Results
NADPH Oxidase Is Involved in Carbon Ion Irradiation–Induced Cell Death
MTT test was performed to estimate cell death after heavy ion irradiation. The viability of HepG2 cells was decreased to 57% compared with control. Pretreatment of 10 µL DPI (an inhibitor of NADPH oxidase) rescued cells from carbon ion radiation–induced cell death. Diphenyleneiodium treatment alone showed limited toxicity to the cells (Figure 1). Similar effects can be observed in both HeLa and GLC-82 cells.

Heavy ion irradiation–induced decrease in cell survival concerned with NADPH oxidase. Cancer cells were treated with or without 10 µL diphenyleneiodium (DPI), an inhibitor of NADPH oxidase, and then the cells were exposed to 4 Gy of heavy ion irradiation. At 48 hours after heavy ion irradiation, the 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyl-2 H-tetrazolium bromide (MTT) test was performed to estimate the cell viability. The values are expressed as means (standard error of the mean [SEM]) of 3 independent experiments. *P < .05, **P < .01 versus the control, n = 5.
NADPH Oxidase Mediates Heavy Ion Irradiation–Induced Elevation of Intracellular ROS
2′,7′-Dichlorofluorescin diacetate (DCFH-DA) staining was used to evaluate relative intracellular ROS level in cancer cells. The pattern of endogenous ROS generation was the same in all the 3 cancer cell lines tested. The level of ROS was increased significantly at 12 hours after 4 Gy irradiation. The ROS level in cells incubated with 10 µL DPI was markedly reduced in cells after 4 Gy heavy ion irradiation compared with heavy ion irradiation alone. The ROS level in cells treated with 10 µL DPI showed no significant difference compared with control (Figure 2).
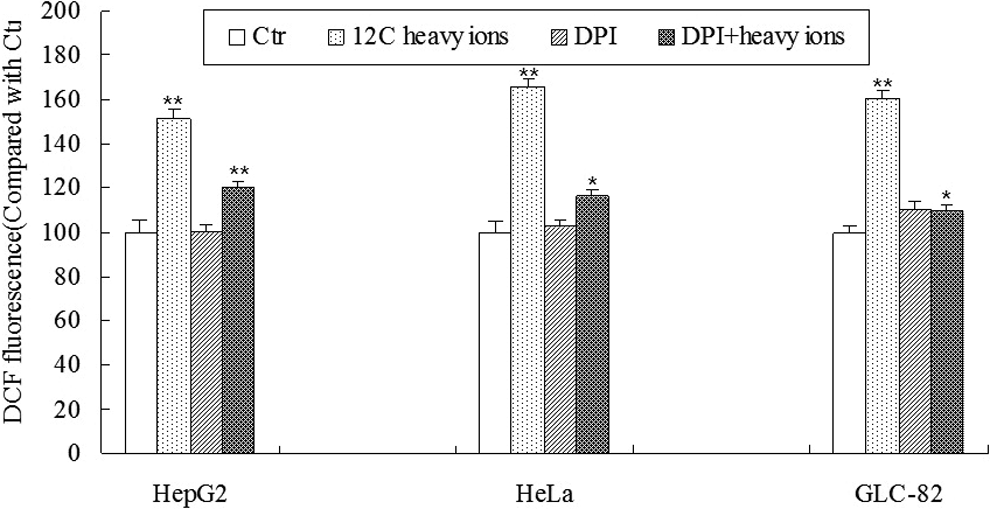
Diphenyleneiodium (DPI) hindered the increased output of reactive oxygen species (ROS) induced by heavy ion irradiation. Cancer cells were treated with or without 10 µL DPI and then the cells were exposed to 4 Gy heavy ion irradiation. The ROS level was measured 12 hours later. The values are expressed as means (standard error of the mean [SEM]) of 3 independent experiments. *P < .05, **P < .01 versus the control, n = 5.
Carbon Ion Irradiation Induces p47phox Subunit to Translocate to the Cell Membrane and Colocalized With NOX2
Immunofluorescence staining was used to determine whether carbon ion irradiation induced activation of NADPH oxidase. Translocation of cytosolic subunit p47phox subunit to the membrane is proposed to be one of the key steps in NADPH oxidase activation. 15 Anti-NOX2 (red fluorescence) combined with anti-p47phox (green fluorescence) were used to detect the localization of NOX2 and p47phox subunit, respectively. p47phox subunit was found to distribute ubiquitously within cytoplasm and nucleus in unirradiated cells. No significant accumulation was detected on the cell membrane. p47phox subunit translocated to the cell membrane and colocalized with NOX2 in all 3 cell lines after 4 Gy carbon ion irradiation, suggesting that NADPH oxidase was activated by carbon ion irradiation (Figure 3). These data suggested that carbon ion could stimulate NADPH oxidase activation via assembling p47phox subunit and NOX2 in cell membrane.

Colocalization of p47phox and NOX2 on the membrane after heavy ion irradiation. Cancer cells were irradiated with heavy ion irradiation (4 Gy) and 1 hour later stained with antibodies against p47phox (green) and NOX2 (red). Cytosolic subunit of p47phox is distributed equally in unirradiated cancer cells but are moved to the membrane and colocalized with NOX2 after 4 Gy heavy ion irradiation.
Carbon Ion Irradiation Upregulates NOX Expression
The expression level of NOX family (including NOX1, NOX2, NOX3, NOX 4, and NOX5) was quantified by Western blotting. As shown in Figure 4, most of the NOX family proteins were upregulated 1 hour after heavy ion irradiation. The induction of NOX expression followed a dose-dependent manner, in which significant higher expression was observed after 4 Gy heavy ion irradiation than that in 1 Gy irradiation. An exception was that in HepG2 cells, the NOX3 expression was lower after 4 Gy irradiation than that in 1 Gy irradiation.
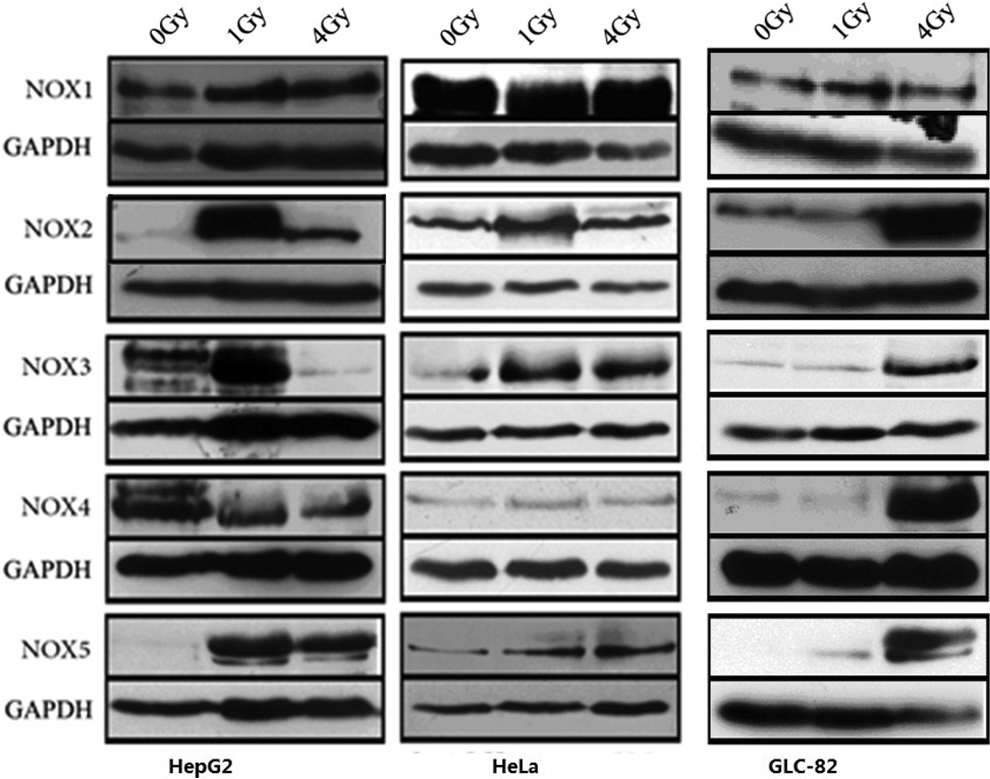
Heavy ion irradiation upregulates the expression of NOX family proteins. Cancer cells were irradiated with heavy ion irradiation (0, 1, and 4 Gy), and 1 hour later, proteins were extracted for Western blotting. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control.
Discussion
Radiation-induced oxidative stress has been suggested to be one of the important causes of cellular inactivation after ionizing radiation. 16,17 However, the main sources of endogenous ROS in irradiated cells remain controversial. Radiation-induced mitochondria dysfunction could be one of the causes of excessive ROS production, but until currently little evidence supports crucial roles of mitochondria-derived ROS in radiation-induced DNA damage. Alternative source of ROS, such as NADPH oxidase, has been shown to be responsible for the chronic oxidative stress in cells after exposure to radiation in vitro and in vivo. 18,19 It has been detected that a chronic oxidative stress response was ascribed to NOX activity in mouse hematopoietic cells after radiation. 18 NOX deregulation can lead to ROS overproduction, which in turn increased oxidative-induced DNA damage.
Heavy ion beams, as high LET radiation, have excellent lethal effect in cancer cells, resulting in multiple double-strand breaks (DSBs) and inefficient DSB repair. 20 Besides the DNA injury caused by direct effect of carbon ion, enhanced ROS levels were also seen in heavy ion beam–induced cell death. 21 However, whether NADPH oxidase contributes to endogenous ROS production by heavy ion beams is unclear. Here we report that NADPH oxidase can be activated by carbon ion irradiation at 2 dosages (1 and 4 Gy), resulting in ROS generation. The initial dose screening performed at 1, 4, and 8 Gy shows a dose-dependent pattern in cell death. Higher dosage radiation (8 Gy) induced a severe cell death and compromised ROS production, probably due to increased cell death or the inability of lethally irradiated cells to produce ROS. 22 Lower dosages (1 and 4 Gy) induced significant ROS response, which was also reported after 2 to 4 Gy carbon ion irradiations. 23 This dose-dependent ROS generation was in accordance with 56Fe ion irradiation at lower dose (<1 Gy). 22 Given previous data showing changed ROS status with dose and a significant biological effect (cell survival and ROS level) in 4 Gy carbon ion exposure, 23 –25 ROS in 3 cell lines were tested with DPI after 4 Gy carbon ion irradiations. The result indicated that increased ROS generation, as produced by active NADPH oxidase, contributed to heavy ion irradiation–induced cell death.
NOXs family expression is associated with carbon ion–induced ROS, which showed that lower cellular ROS levels by either inhibiting or silencing NOX4 or NOX5 protect against radiation-induced DNA damage and subsequent cell death. 19 In addition, IR-induced hematopoietic stem cell injury can be ameliorated by decreasing ROS production in association with the downregulation of NOX4 expression. 26 Therefore, elevated expression of NOX1 through NOX5 induced by carbon ion irradiation contributed to increased ROS generation. Interestingly, there were much higher level of NOX2, NOX3, and NOX5 expression at 1 Gy compared with that at 4 Gy in HepG2 cell line, which did not occurr in GLC-82 cell line. These changes with dose may help determine the likelihood of late effects of radiation exposure. 27 The translocation of p47phox subunit to the cell membrane and its colocalization with NOX2 initiate NADPH oxidase-mediated ROS generation. These findings elucidate a new mechanism that may contribute to heavy ion–induced cell death. We observed the colocalization of p47phox and NOX2 in the membrane in 3 cell lines tested at 2 dosages (1 and 4 Gy), which indicated the activation of NADPH oxidase in response to heavy ion irradiation.
Besides heavy ion radiation, X-ray radiation could also significantly activate NADPH oxidase and enhance the production of ROS, leading to X-radiation-induced growth inhibition of nasopharyngeal carcinoma cells. 28 It has been observed that Nox4 expression continues to increase at all time points from 1 day to 6 months post X-ray radiation in mice lung tissue. 29 Similarly, low-dose γ irradiation is capable of upregulating NADPH oxidase in central nervous system in mice. 30 Increased NADPH oxidase expression is possible sources of increased ROS production mirrored by an increase in DNA damage. Further investigation on the NOX’s activities in endogenous ROS production will help to clarify the additive effects of radiation-induced DNA damage. Our current data support the hypothesis that oxidative stress at least plays a causative role in radiation-induced cellular inactivation. 31 –33 Taken together, these data provide evidence that NADPH oxidase consisting of NOX contributes to the cellular inactivation of cancer cells via ROS generation.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Key Program of National Natural Science Foundation of China (U1432248), National Key projects of Research and Development (2016YFC0904600), the National Natural Science Foundation of China (11505245), and the Western Talents Program of the Chinese Academy of Sciences (Y460040XB0).