
Abstract
Guangsi Yulin Decoction (GSYLD) is a traditional Chinese medicine (TCM) formula with therapeutic efficacy against spermatogenesis dysfunction. However, the underlying mechanism of the alleviation remains unclear. To investigate the effects of GSYLD on Tripterygium wilfordii polyglycolide (GTW)-induced spermatogenesis dysfunction in mice and to explore associated mechanisms, particularly its role in oxidative stress and inflammation. A total of 60 (7–8-week-old) BALB/c mice were grouped into control (NC), model (GTW), low-dose GSYLD (GSYLD-L, 0.27 g/kg/d), high-dose GSYLD (GSYLD-H, 1.08 g/kg/d), and PTEN inhibitor (1.08 g/kg/d GSYLD-H + 0.3 mg/kg PTEN inhibitor). After eight weeks, the model was validated, and semen quality was evaluated by assessing sperm vitality and concentration. Testicular tissues were analyzed by H&E staining, while Western blotting and RT-qPCR (Reverse Transcription Quantitative PCR) were used to assess the therapeutic efficacy of the GSYLD. In comparison to the GTW group, the GSYLD-H group showed a significant improvement in sperm vitality and concentration (p < .01). GSYLD upregulated PI3K, AKT, and FoxO1, while downregulating PTEN mRNA expression (p < .001). A parallel recovery was observed at the protein level. Specifically, the GSYLD-H group exhibited a significant amelioration of oxidative stress and inflammation, marked by reduced MDA, TNF-α, and IL-6 levels (p < .01) and enhanced SOD activity (p < .001) relative to the GTW group. GSYLD alleviated GTW-induced spermatogenesis dysfunction, potentially through PTEN/PI3K/AKT/FoxO1-mediated regulation of oxidative stress and inflammation.
Introduction
Male sterility refers to the inability to achieve pregnancy after more than 1 year of regular unprotected intercourse, primarily due to male-associated factors (Agarwal et al., 2021). Globally, sterility affects 8% to 12% of couples, with male-associated factors contributing to approximately 50% of sterility cases. Epidemiological investigations have shown a significant decline in semen quality among Chinese males between 1981 and 2019, accompanied by a continuous increase in the incidence of male sterility (Lv et al., 2021). The etiology of male sterility is highly complex and involves multiple determinants, including testicular dysfunction, congenital anatomical abnormalities, environmental exposures, lifestyle factors such as tobacco use and obesity, gonadotoxic agents, and aging. Spermatogenesis dysfunction, characterized by impaired sperm production, maturation, and function, represents a major contributor to male infertility (Khosravi et al., 2021; Sciorio et al., 2025). Clinically, this condition is presented as decreased sperm concentration, reduced motility, abnormal morphology, and impaired fertilizing capacity, all of which hinder successful fertilization. Current pharmacological interventions for spermatogenesis dysfunction, such as coenzyme Q10, carnitine, and vitamin E, remain largely empirical, and robust evidence-based therapies are lacking (Eisenberg et al., 2023). The absence of effective drugs and therapies highlights a significant need in this field. Although assisted reproductive technologies can bypass many barriers to conception, these approaches remain subject to ethical, genetic, financial, and safety concerns (Berntsen et al., 2025; Blake et al., 2019). Thus, novel therapeutic strategies that restore spermatogenesis represent an urgent clinical need.
Traditional Chinese Medicine (TCM) is widely regarded as a safe therapeutic option and an important component of alternative and complementary medicine, with reported capacity to improve seminal parameters (Wang et al., 2024; G. Yan et al., 2021; Y. Zhu, Zhang et al., 2024). In traditional Chinese medicine theory, “kidney essence” refers to the innate vital substance stored in the kidney, which governs growth, development, and reproductive functions. The association between kidney essence and sperm production arises from clinical observations that tonifying kidney essence improves spermatogenesis. Modern studies provide preliminary scientific basis: for example (Li et al., 2021) demonstrates that the kidney essence-tonifying formula BZBS ameliorates sperm quality in aging mice by increasing sperm density and motility. GSYLD is a classical TCM prescription for treating spermatogenesis dysfunction. Experimental evidence has suggested that SC-FL, a key constituent, restores blood–testis barrier integrity through regulation of the MAPK signaling cascade and inhibits spermatogonial apoptosis via the SCF/c-kit/PI3K/Bcl-2 pathway, thereby improving spermatogenic function in murine models (Guan et al., 2020). However, the GSYLD protective mechanisms in spermatogenesis dysfunction remain incompletely understood.
Tripterygium wilfordii polyglycolide (GTW), an active compound extracted from Tripterygium wilfordii, is known to impair reproductive function following prolonged exposure (Qin et al., 2023). Experimental studies have validated that GTW administration effectively induces spermatogenesis dysfunction in murine models (Ding et al., 2023). This research aims to clarify the mechanisms through which GSYLD exerts its protective influence against GTW-induced spermatogenesis dysfunction. The findings may yield significant insights and deepen comprehension, potentially laying the groundwork for progress in innovative treatment strategies targeting spermatogenic dysfunction.
Materials and Methods
Animal Model
Male BALB/c mice (7–8 weeks old, 22 ± 2 g) were procured from Charles River Experimental Animal Technology Co. (Beijing, China). They were kept in highly specific pathogen-free (SPF) and an environmentally controlled conditions (a 12-hour light/dark cycle, 22°C, 60% humidity). Also, food and water were provided ad libitum. The current research was carried out in full compliance with the National Institutes of Health (NIH) guidelines for laboratory animal welfare, under a protocol validated by the Medical and Laboratory Animal Ethics Committee of Beijing University of Chinese Medicine (Approval No. DZMYY24-28).
Experimental Procedure and Treatment Protocol
Following one week of acclimatization, all mice were weighed and randomly assigned using a random number table to the following groups: normal control (NC), model (GTW), low-dose intervention (GSYLD-L, 0.27 g/kg/d), high-dose intervention (GSYLD-H, 1.08 g/kg/d), and PTEN inhibitor (GSYLD-H, 1.08 g/kg/d combined with PTEN inhibitor 0.3 mg/kg). The NC group served as the untreated control and received a daily dose of distilled water (10 mL/kg). To establish the model of spermatogenesis dysfunction, the remaining experimental groups were administered GTW (Zhejiang Deende Pharmaceutical Co., Ltd., batch no. 2310122B) via daily intragastric gavage at a dose of 10 mL/kg/day. This volume delivered a daily GTW concentration of 40 mg/kg, based on a formulation of 4 mg GTW per 1 mL.
GSYLD, a decoction comprising 10 traditional Chinese medicines, Chinese wolfberry (Gouqizi), dodder seeds (Tusizi), schisandra fruit (Wuweizi), raspberry (Fupenzi), plantain seeds (Cheqianzi), prepared rehmannia (Shudihuang), astragalus (Huangqi), Chinese yam (Shanyao), Chinese angelica (Danggui), and raw oyster shell (Shengmuli), was purchased from Dongzhimen Hospital (Beijing University) and prepared as a decoction. GSYLD was administered orally once daily for 4 weeks at either 0.27 g/kg/d or 1.08 g/kg/d. From weeks 1–8 of the experiment, the PTEN inhibitor group received BPV (HY-136065, MCE, USA) at 0.3 mg/kg/d.
Mice were weighed and euthanized by isoflurane inhalation at the end of treatment. Testes and epididymides were harvested. Fixation of the testicular tissues was carried out with paraformaldehyde (PFA, 4%) for histological and ultrastructural analyses, while the remaining specimens were kept at –80°C for analyzing mRNA and protein expression. The semen was collected from the cauda epididymis of the mice and analyzed using Computer-Aided Sperm Analysis (CASA). The remaining tissues were preserved in liquid nitrogen for further analysis.
Evaluation of Testicular and Epididymal Organ Indicators
The testicular organ coefficient was calculated by determining testis mass with a precision balance and applying the formula: (testicular weight / body weight) × 100%.
Sperm Concentration and Viability Analysis
Epididymal tissue was minced in phosphate-buffered saline (PBS) and subjected to a 15-minute incubation in a water bath maintained at 33°C. A sample of the resulting sperm suspension was placed on a slide and analyzed using an automated semen analysis system (Hamilton Thorne, MA, USA).
Testicular Tissue Staining With Hematoxylin and Eosin (H&E)
To evaluate the structural integrity of spermatogenesis and supporting cells, as well as the seminiferous tubules, testicular specimens were fixed in 4% paraformaldehyde (PAFA) over a 48-hour period. Tissue dehydration was carried out while employing graded ethanol, followed by clearing with xylene, and preparing paraffin-embedded blocks. For histological analysis, 6 µm-thick sections were prepared and subjected to H&E staining. Sections were observed under a light microscope to evaluate histological features, and images were acquired for further assessment. For each slide, five distinct microscopic fields were examined, and the procedure was repeated three times for each slide. To minimize bias, evaluations were conducted in a blinded manner. Testicular spermatogenic function was assessed according to the Johnsen scoring criteria, using a 10-point scale to quantify the health status, with higher scores indicating better spermatogenic function (Ito et al., 2021).
Serological Analyses of TNF-α and IL-6 Using ELISA
Blood samples were obtained from all mice via the abdominal aorta. Following coagulation, blood samples were centrifuged at 4,000 r/min for 10 min to obtain serum. Serum IL-6 (MM-0163M2) and TNF-α (MM-0132M2) concentrations were measured while employing ELISA kits (Jiangsu Meimian Immunoassay Industry Co., Ltd.).
Measurement of Oxidative Stress Markers
Testicular levels of superoxide dismutase (SOD) and malondialdehyde (MDA) were determined using commercially available assay kits (Nanjing Jiancheng Bioengineering Institute, China; A001-3-2 and A003-1-2, respectively).
Western Blot
Testicular tissues preserved in liquid nitrogen were thawed slowly on ice. RIPA buffer (Cell Signaling, USA) was added at 10 times the tissue volume, and samples were homogenized for 15 min. Following centrifugation of the lysates (15 min, 12,000 × g, 4°C), the supernatant was carefully gathered and quantified for protein content with a commercial BCA kit (SolarBio).
Samples were diluted accordingly to ensure concentrations aligned with the linear range of the standard curve. Each protein sample was mixed with 5× reduced protein loading buffer (4:1) and denatured in a water bath by boiling for 15 min. Protein separation was carried out using SDS-PAGE (10%), followed by transferring them onto a methanol-activated PVDF membrane. To facilitate specific antigen-antibody binding, membranes were first blocked with 5% skim milk. Target proteins were then identified through an overnight incubation with primary antibodies at 4°C, a step critical for binding specificity. The following day, a series of three 5-minute TBST washes ensured the removal of nonspecifically bound antibodies prior to a 1-hour room temperature incubation with HRP-labeled secondary antibodies, which enabled subsequent chemiluminescent detection. Signal detection was performed using ECL, and band intensities were quantified with ImageJ. All protein expression data were normalized against the β-actin signal as an internal control. All experiments were performed in three independent biological replicates.
RT-qPCR Analysis
Total RNA was extracted from testicular samples with the TRIpure reagent (G3013, Wuhan Xavier Biotechnology Co., Ltd.) as recommended. Extracted RNA concentrations were measured using a Nanodrop 2000, and A260/A280 ratios were verified to be 1.8-2.0, ensuring purity. The obtained RNA was subsequently reverse-transcribed into cDNA using 5X SweScrlpt All-in-One SuperMix (G3337, Wuhan Xavier Biotechnology Co., Ltd.), which was then employed as the template for quantitative PCR. The amplification protocol was executed on a Bio-Rad CFX CT 96-well system using a two-stage thermal profile: an initial denaturation for 30 seconds at 95°C, followed by 40 cycles of denaturation for 15 seconds at 95°C and annealing/extension for 30 seconds at 60°C.
Following amplification, a melting curve analysis was performed to confirm the amplification of a single, specific PCR product. In this experiment, β-actin served as the endogenous control gene. The 2−ΔΔCt approach was applied to determine relative expression levels of the genes of interest. Final results were presented as fold changes compared with the control group, which was normalized to 100%. The corresponding primer sequences are listed in Table 1.
The List of Primer Sequences Applied for Gene Expression Profiling.

Statistical Analysis
The obtained values are expressed as means ± standard deviation (means ± SD). Data analysis was performed using SPSS (v26.0; IBM Corp., NY, USA). Intergroup comparisons were conducted using an unpaired Student’s t-test or one-way analysis of variance (one-ANOVA), where appropriate. Where analysis of variance indicated significant differences, the Student-Newman-Keuls (SNK) method was applied for multiple comparisons. A p-value of less than 0.05 was considered the criterion for statistical significance throughout the study.
Results
Measurement of Body Weight and Analyzing Organ Coefficients
No significant difference was detected in body weight between the groups after the experimental period (Figure 1A). In comparison to the NC group, the testicular organ coefficient significantly (p < .05) diminished in the GTW group (Figure 1B). When compared with the GTW group, the testicular organ coefficients in the GSYLD-L, GSYLD-H, and PTEN inhibitor groups exhibited a recovery trend, although intergroup differences did not reach statistical significance.
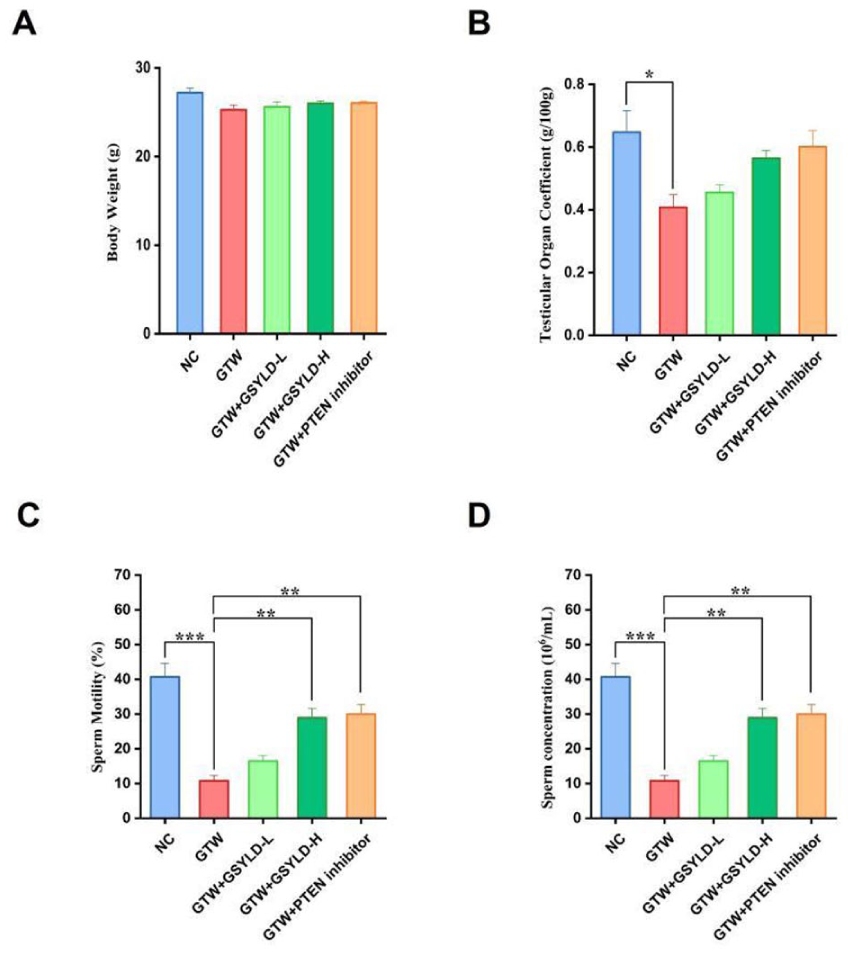
The Impact of Guangsi Yulin Decoction on Body Weight, Testicular Coefficient, and Semen Quality in Mice. (A) Post-Experimental Body Weigh in Mice; (B) Testicular Organ Coefficient; (C) Sperm Motility; (D) Sperm Concentrations.
GSYLD Improve Semen Quality
Statistical analysis showed a significant reduction in sperm motility and concentration in the GTW group compared with the NC group (Figure 1C and D; p < .001). Post hoc comparisons showed that such a deficit was significantly attenuated by high-dose GSYLD and the PTEN inhibitor (p < .01). Although the low-dose GSYLD group exhibited an improving trend, the difference from the GTW group was not statistically significant.
GSYLD Attenuates Pathological Damage of Testicular Tissues for GTW-Induced Spermatogenesis Dysfunction
Histological analysis of testicular tissues (Figure 2) revealed that seminiferous tubules in the NC group maintained normal morphology, with clearly defined lumens and intact basement membranes separating interstitial compartments. However, testes from the GTW group displayed structural abnormalities, including a high frequency of malformed seminiferous tubules, expanded intertubular spaces, disrupted and fragmented luminal organization, and cytoplasmic vacuolation in Sertoli cells. The Johnsen score of the GSYLD-L group was higher than that of the GTW group (p < 0.05; Table 2), although it remained lower than the scores in the GSYLD-H and PTEN inhibitor groups.
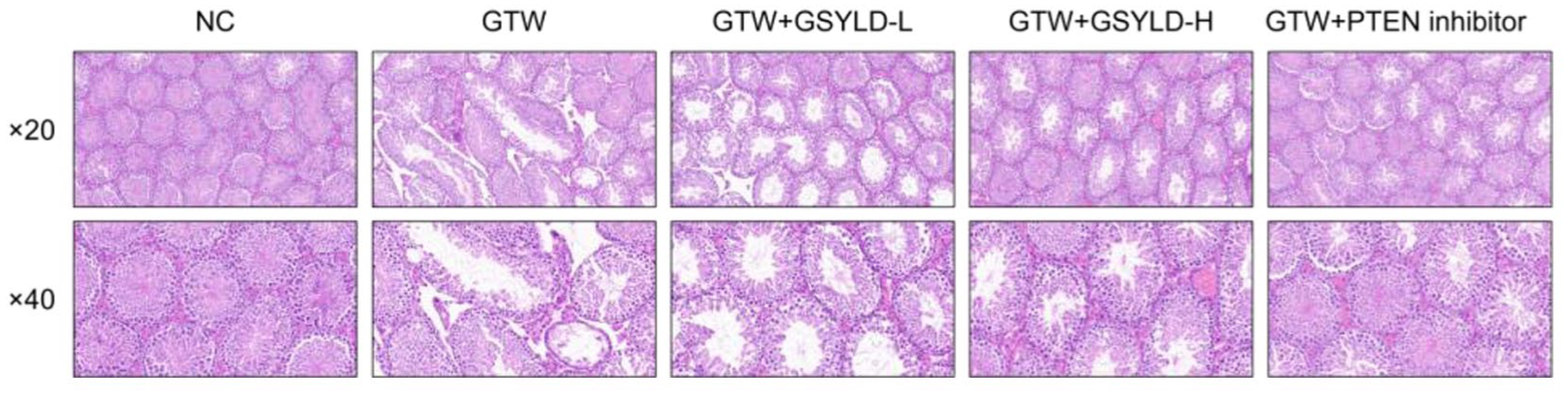
Testicular Morphology.
Comparison of Johnsen Scores for Spermatogenic Function in Mouse Testicular Tissues.
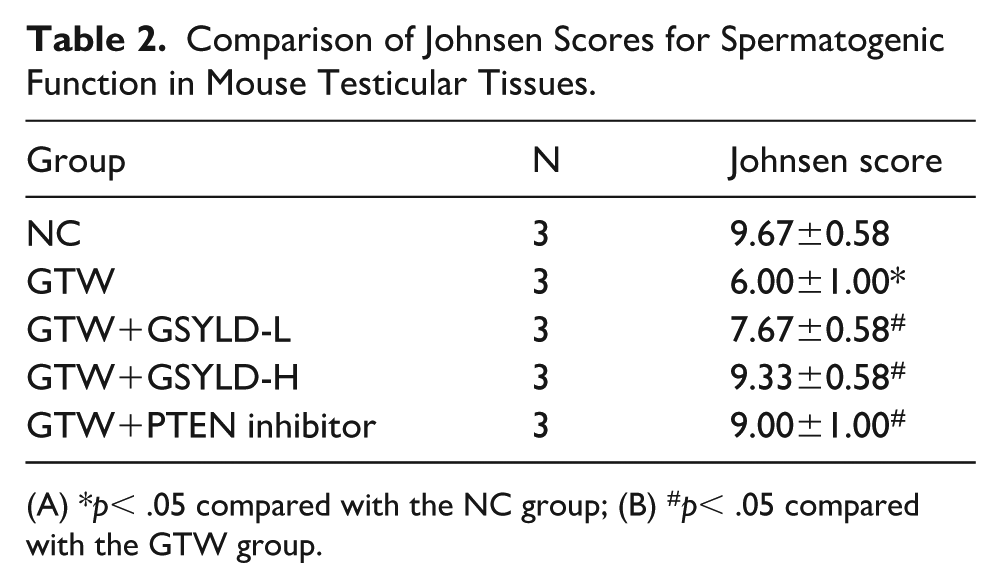
(A) *p< .05 compared with the NC group; (B) #p< .05 compared with the GTW group.
GSYLD-Mediated Amelioration of GTW-Induced Spermatogenesis Dysfunction via Testicular Inflammatory Cytokine Regulation
Cytokine profiling via ELISA identified a pronounced inflammatory response in GTW-treated mice, characterized by significantly increased circulating levels of IL-6 and TNF-α relative to the NC group (Figure 3A and B; p < .001). A significant reversal of GTW-induced inflammation was achieved in the GSYLD-H and PTEN inhibitor groups, both of which demonstrated significantly lower concentrations of TNF-α and IL-6 relative to the GTW group (Figure 3A, p < .01; Figure 3B, p < .001). Although the GSYLD-L group showed a downward trend compared with the GTW group, these differences were not statistically significant.
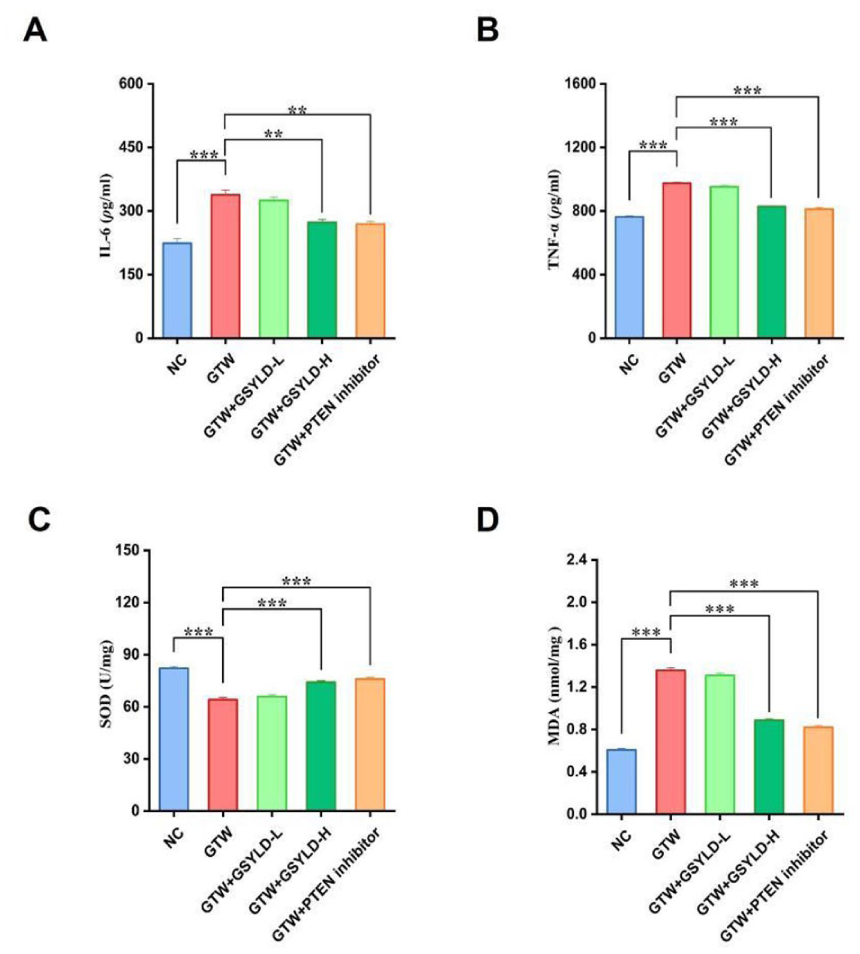
The Serum Concentrations of (A) IL-6, (B) TNF-α, (C) SOD, and (D) MDA in Mice (n = 3).
GSYLD Improves SOD and MDA Levels in the Mouse Model for GTW-Induced Spermatogenesis Dysfunction
Spectrophotometric analysis revealed that, compared with the NC group, the GTW group showed significantly lower SOD levels (p < .001; Figure 3C) and markedly higher MDA levels (p < .001; Figure 3D). Both the GSYLD-H and PTEN inhibitor groups showed higher SOD levels and lower MDA levels than the GTW group (Figure 3C and D; p < .001). While the GSYLD-L group exhibited a favorable trend relative to the GTW group, the observed differences were not statistically significant.
Quantitative Analysis of Proteins Involved in the PTEN/PI3K/AKT/FoxO1 Pathway
Western blot analysis revealed a significant disruption of the PTEN/PI3K/AKT pathway in the GTW group relative to the NC group, characterized by downregulated phosphorylation of both PI3K and FoxO1 (p < .05; Figure 4B and E) and a more pronounced reduction in AKT phosphorylation (p < .01; Figure 4C). However, PTEN protein expression was increased (Figure 4F; p < .01). In comparison to the GTW group, the GSYLD-H and PTEN inhibitor groups exhibited recovery trends in p-PI3K, p-AKT, p-FoxO1, and PTEN expression. Notably, PTEN expression was significantly decreased and p-AKT levels were significantly increased in the PTEN inhibitor group compared with the GTW group (Figure 4C and 4F; p < .05).
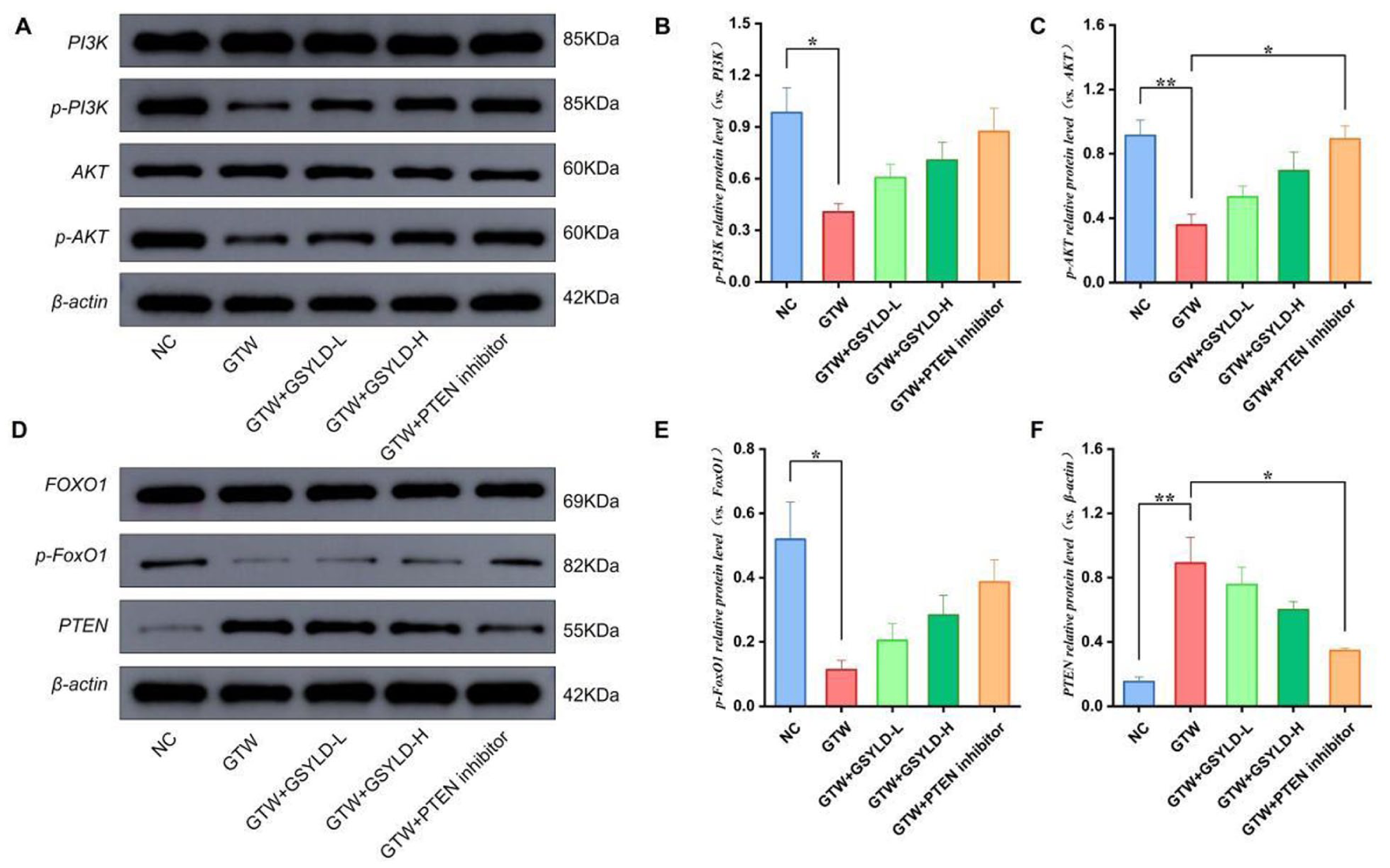
The Impact of Guangsi Yulin Decoction on Protein Expression Related to the PTEN/PI3K/AKT/FoxO1 Pathway. (A–C) Immunoblot Bands and Quantitative Results for AKT, PI3K, p-AKT, and p-PI3K (n = 3). (D–F) Immunoblot Bands and Quantitative Results for FoxO1, p-FoxO1, and PTEN (n = 3).
Expression Analysis of Genes Involved in the PTEN/PI3K/AKT/FoxO1 Axis
The GTW group exhibited significantly lower gene expression levels of PI3K, AKT, and FoxO1 relative to the NC group, denoting a broad downregulation of this signaling axis (Figure 5A–C; p < .001). However, the gene expression level of PTEN was significantly elevated (Figure 5D; p < .001). Relative to the GTW group, the GSYLD-H and PTEN inhibitor groups showed significantly increased PI3K, AKT, and FoxO1 expression (Figure 5A–C; p < .001) and significantly decreased PTEN expression (Figure 5D; p < .001). Although the GSYLD-L group showed a recovery trend, differences compared with the GTW group did not reach statistical significance.
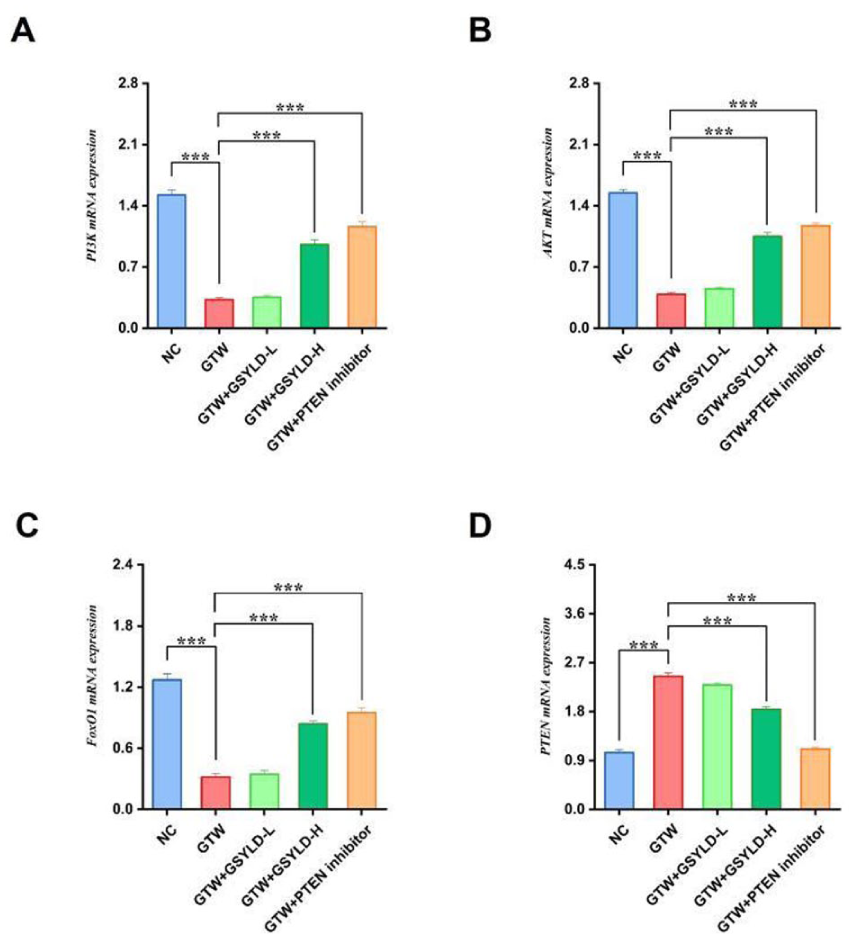
Effects of Guangsi Yulin Decoction on mRNA Expression Related to the PTEN/PI3K/AKT/FoxO1 Pathway. (A) PI3K. (B) AKT. (C) FoxO1. (D) PTEN.
Discussion
Globally, the intensifying challenges of population aging and declining fertility rates have raised increasing concern regarding male fertility (Minhas et al., 2025). At present, no effective interventions exist to reverse this trend. Clinical therapeutic strategies for spermatogenesis dysfunction remain limited in application (Bouhadana et al., 2025). Therefore, identifying novel therapeutic agents to improve semen quality carries significant societal importance. TCM represents an effective clinical option for managing spermatogenesis dysfunction. It is regarded as both a vital component of complementary and alternative medicine and a treatment with favorable safety (F. Yan et al., 2025; R. Zhu, Gao et al., 2024). Also, in a mouse model of GTW-induced spermatogenesis dysfunction, GSYLD (i) significantly increases sperm concentration and motility; (ii) alleviates pathological injury in testicular tissues; (iii) reduces oxidative stress in spermatogonia; (iv) activates the PI3K/AKT pathway while inhibiting PTEN expression, thereby promoting spermatogonial proliferation; and (v) upregulates FoxO1 expression, subsequently inhibiting the release of IL-6, TNF-α.
Testicular tissues consist of seminiferous tubules and interstitial compartments. The walls of seminiferous tubules are formed by the seminiferous epithelium, which comprises supporting cells and spermatogenic cells (Sosa et al., 2025). The development of sperm from spermatogonia is known as spermatogenesis, which proceeds through three distinct stages: spermatogonial proliferation, spermatocyte meiosis, and spermiogenesis. Thus, damage to testicular architecture severely compromises sperm production. In this study, GTW caused severe damage to murine testicular tissues, including seminiferous tubule disruption, interstitial edema, and vacuolization of Sertoli cell cytoplasm. However, the GSYLD-H group and the PTEN inhibitor group demonstrated improved spermatogonia morphology, a more organized seminiferous tubule structure, increased numbers of supporting cells, and reduced histological lesions. These histological improvements were accompanied by increased sperm vitality and concentration in the GTW-induced spermatogenesis dysfunction model.
The proliferation and differentiation of spermatogonia in the testes are regulated by the PI3K/AKT pathway (Lim et al., 2019). PI3K, an intracellular kinase stimulated by receptor tyrosine kinases (RTKs), is composed of two main subunits: p110, which functions as the catalytic component, and p85, which acts as the regulatory component (Huang et al., 2024). AKT is the major downstream effector of PI3K. Upon PI3K activation, phosphatidylinositol 3,4,5-trisphosphate (PIP₃) translocates to the plasma membrane, leading to AKT phosphorylation at Thr308 and Ser473. Activated AKT subsequently enters the cytoplasm and phosphorylates downstream substrates to control diverse cellular processes (Su et al., 2011).
FoxO1, a key transcription factor downstream of AKT, is regulated by PI3K/AKT-mediated phosphorylation and dephosphorylation (Chen et al., 2025). FoxO1 effectively contributes to the self-renewal and differentiation of spermatogonial stem cells (Sinha et al., 2018; Goertz et al., 2011). Following activation, AKT, as a key serine/threonine kinase, translocates from the cytoplasm into the nucleus, where it phosphorylates FoxO1 at specific amino acid residues, thereby causing its export to the cytoplasm. Although FoxO1 retains the ability to bind DNA, its transcriptional activity is lost when it cannot access target gene promoters. Importantly, nuclear FoxO1 functions as a transcriptional activator for pro-inflammatory genes. In the nucleus, FoxO1 binds to the promoter of the TLR4 gene, acting as a “key” to “turn on the switch,” and initiating its transcription and subsequent expression. Elevated TLR4 levels subsequently activate downstream inflammatory cascades (e.g., NF-κB signaling pathway), resulting in the increased secretion of TNF-α, IL-6, and IL-1β (Guo et al., 2019).
PTEN, a key negative regulator of the PI3K/AKT pathway, suppresses signal transduction by dephosphorylating PIP3 to PIP2, thereby inhibiting AKT activation (Liu et al., 2017). Thus, PTEN reduces the activity of the PI3K/AKT pathway. Loss or mutation of PTEN leads to abnormal accumulation of PIP₃, causing excessive PI3K/AKT pathway activation (Han et al., 2024). Overactivated AKT elevates ROS production through NADPH oxidase, while PTEN deficiency decreases the expression of SOD, thereby reducing the ROS clearance capacity (Kang et al., 2009).
Our results demonstrate that administration of high-dose GSYLD significantly alleviated GTW-induced spermatogenic dysfunction, an improvement comparable to PTEN inhibition. This was evidenced by the recovery of sperm parameters, reduced testicular pathological injury, alongside decreased levels of IL-6, TNF-α, and MDA, and restored antioxidant capacity (SOD). These findings suggest that the therapeutic effects may be attributed to the effective regulation of the PTEN/PI3K/AKT/FoxO1 pathway and the reversal of abnormal mRNA expression of key genes.
This study observed that high-dose GSYLD intervention significantly reversed the abnormal mRNA expression of genes in the GTW-induced PTEN/PI3K/AKT/FoxO1 pathway, whereas phosphorylation levels of the corresponding proteins (p-PI3K, p-AKT, p-FoxO1) showed only an improving trend without reaching statistical significance. This inconsistency between mRNA and protein levels suggests complex post-transcriptional regulatory mechanisms within the pathway. Possible explanations include (a) although mRNA levels were restored after GSYLD intervention, translation may have been inhibited by microRNAs or RNA-binding proteins; (b) following GSYLD intervention, the protein synthesis may have been increased, but simultaneous acceleration of degradation through the ubiquitin–proteasome system or autophagy–lysosome pathway may have offset this, leading to stable net protein levels; (b) the time point assessed may have captured only transcriptional responses, while proteomic responses had not yet occurred, indicating a lag effect. Further in-depth studies are warranted to explore the underlying mechanisms.
This study has several limitations. First, the investigation of the PTEN/PI3K/AKT/FoxO1 pathway relied on pharmacological inhibition; genetic gain- or loss-of-function models would provide more definitive causal evidence. Second, and more pertinent to confounding, the complex nature of the GSYLD formula itself presents an inherent challenge. Its multitude of bioactive components means that biological effects observed in this study could be confounded by interactions with pathways beyond the one we focused on. Finally, despite rigorous randomization and standardized housing conditions, unmeasured confounding variables, such as subtle differences in gut microbiota composition affecting drug metabolism or minor fluctuations in environmental stressors, could potentially influence individual animal responses and introduce bias into the outcomes.
Notwithstanding these limitations, the dose-dependent effects and the synergistic results with the PTEN inhibitor strengthen the association between GSYLD and the observed improvement in spermatogenesis. Future studies employing compound-knockout approaches or isolating active constituents will be crucial to minimize these confounding influences and confirm the precise mechanistic targets.
Conclusion
In a mouse model of GTW-induced spermatogenesis dysfunction, GSYLD demonstrated potential to improve spermatogenesis. The mechanism may involve activation of the PTEN/PI3K/AKT/FoxO1 pathway, reduction of oxidative stress, and suppression of inflammatory cytokine release, thereby alleviating spermatogenesis dysfunction. These findings expand current understanding of how GSYLD promotes male fertility and provide experimental support for its potential application in managing male infertility. Future studies may focus on isolating and characterizing the specific bioactive constituents of GSYLD to identify essential molecules that preserve spermatogenic capacity, facilitating the development of novel pharmacological interventions for spermatogenesis-related disorders.
Footnotes
Consent for Publication
Not applicable.
Author Contributions
All authors contributed to the final version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Beijing Natural Science Foundation (Grant No. 7244488); National Natural Science Foundation of China (82174385); Major Difficult Diseases Clinical Cooperation Project between Chinese and Western Medicine by the National Administration of Traditional Chinese Medicine of China (ZDYN-2024-A-046); “Unveiling the List and Appointing the Commander” Project by Beijing University of Chinese Medicine (2025-JYB-JBGS-013).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.