
Abstract
Background
The long-term impact of traumatic brain injury (TBI) on ageing remains poorly understood without controlled studies.
Objective
Cross-sectionally examine how chronic moderate–severe TBI and advancing age interact to affect brain structure and related cognition in adults aged 40 to 90 years who received compensated inpatient rehabilitation in Australia.
Methods
Cross-sectional magnetic resonance imaging (MRI) and neuropsychological data were collected from 123 individuals with chronic moderate–severe TBI (6-33 years post-injury; Mage = 59.64) and age- and gender-matched to 74 non-TBI controls (Mage = 62.10). Analyses examined cortical thickness, subcortical volume, and white matter microstructural integrity using tract-based analysis.
Results
Ageing and TBI were independently associated with reduced structural integrity in multiple regions. However, age moderated the effect of TBI on both fornix integrity and left thalamic volume, such that TBI-related reductions were present in adults in their 40s to 70s and 40s to 60s, respectively, but attenuated in the oldest age groups (80+ and 70+, respectively) relative to controls. Fornix and thalamic reductions correlated with poorer cognitive function, with the largest effects in verbal memory and processing speed.
Conclusions
While TBI and ageing were independently associated with changes in brain structure, their interaction manifested distinctly in the bilateral fornix and left thalamus, with TBI-related structural losses in middle-age that diminished in later life as controls experienced typical age-related declines. In adults who received inpatient rehabilitation, moderate–severe TBI may result in initial structural alterations that do not accelerate age-related declines or compound with ageing processes long-term. Age-dependent fornix and thalamic structural changes were strongly associated with cognitive performance, particularly in verbal memory and processing speed.
Introduction
Traumatic brain injury (TBI) is a significant cause of chronic disability worldwide. 1 As life expectancy rises, along with improvements in healthcare, a growing number of individuals are surviving TBI and experiencing its long-term effects into older adulthood.2-4 However, the intersection between TBI and ageing is poorly understood, with limited insight into whether their combined effects lead to accelerated, additive, or distinct patterns of structural brain changes.
TBI results in persistent structural and functional brain changes.5,6 Proposed mechanisms that may contribute to ongoing structural changes following TBI include environmental impoverishment, cognitive disuse, neuroinflammation, and neuroendocrine dysfunction.7-9 Following moderate–severe TBI, studies have demonstrated significant grey and white matter volume loss that persisted in the subacute (6-24 months post-injury)7,10 and chronic (5-22 years post-injury)11,12 phases. The frontal and temporal lobes are particularly vulnerable to adverse changes following TBI.7,9,12 Normal ageing similarly affects these regions most prominently, characterised by grey and white matter volume loss, neurochemical deficits (eg, dopaminergic), and altered neural activity.13,14 It remains unclear whether the progression of structural brain changes with ageing from middle to older adulthood differs for individuals with TBI, including whether TBI leads to premature ageing or an accelerated decline in brain integrity in the decades following injury. 9 No studies to date have directly examined the interaction between age and chronic TBI in influencing grey or white matter integrity. Notably, many neuroimaging studies either omit age from their analyses or adjust for it as a covariate, limiting our understanding of how TBI and ageing interact.7,11,12
Although some studies suggest that older age is associated with poorer structural outcomes or ageing after TBI, most lack age-matched controls or have not extended beyond 6 months post-TBI.15-17 Control groups are necessary to distinguish the effects of TBI from normal ageing. If TBI exacerbates age-related structural declines, older adults with TBI should exhibit greater structural loss compared to both middle-aged adults with TBI (reflecting age-related differences) and age-matched controls without TBI (indicating TBI-related effects). This requires studying outcomes beyond the acute and subacute recovery phases, to observe ageing processes while minimising confounding effects of ongoing neurological recovery or secondary pathophysiological processes after injury. 18
The relationship between structural brain changes and cognition in chronic TBI is established,11,12,19 but has been scarcely examined in older adults. We previously found cognitive impairments 6 to 33 years post-TBI, that were not exacerbated in older age when compared cross-sectionally to age- and gender-matched controls. 20 Rather, verbal memory was impaired in middle-aged adults with TBI while performance in older adults was comparable to age-matched controls. Whether structural brain changes are related to and follow a similar pattern to cognition after injury or demonstrate an adverse effect of older age remains unknown.
In this study, we cross-sectionally examined the interactive effects of older age and moderate- severe TBI on cortical thickness, subcortical volume, and white matter integrity using MRI, comparing individuals with TBI who received inpatient rehabilitation to a non-TBI control group. We hypothesised that those with TBI would exhibit reduced structural integrity, volume, and cortical thickness, with a more pronounced reduction in older age compared to controls, particularly in the frontal and temporal brain networks, given their susceptibility to both ageing and TBI. Additionally, we investigated whether structural changes associated with ageing post-TBI were related to cognitive function in the chronic phase. Since previous research has linked cognitive performance to specific brain structures following TBI,21,22 we focused on regions and tracts where we found significant age × group interactions, predicting that reduced structural integrity or thickness would correlate with cognitive dysfunction, with domain-specific effects depending on the regions affected.
Methods
Standard Protocol Approvals, Registrations, and Patient Consents
The study conformed to the Declaration of Helsinki and was approved by the Monash Health Human Research Ethics Committee (ID 90582). Written informed consent was obtained from all participants (or guardians of participants) in the study. To date, all participants have had capacity to provide their own informed consent; no participants have required guardian consent due to impairment.
Participants
The TBI group were recruited from inpatient rehabilitation between 1985 and 2019 as part of a longitudinal TBI study that did not include cognitive testing or MRI. 23 Cross-sectional data using the current study’s cognitive and neuroimaging assessments were collected between 2017 and 2024. Inpatient rehabilitation was delivered under a state-wide no-fault accident compensation scheme. The TBI group were medically-confirmed to have sustained a single moderate–severe TBI (Mayo classification) 24 from age 16 years and were at least 5 years post-injury. This threshold was applied to ensure findings reflected the chronic phase rather than ongoing recovery in the initial years after injury, during which secondary neuropathological processes such as apoptosis, Wallerian degeneration, and inflammation are thought to be active and not yet stabilised.17,25,26 Structural losses occurring during this period are therefore considered distinct from the long-term age-related changes of interest in the current study. A convenience sample of non-TBI controls with no history of TBI, concussion, or other loss of consciousness were recruited from the community using advertisements (social media and newspaper). We recruited adults from age 40 years to study ageing with a specific focus on older adults with TBI, since age-related declines often emerge around middle-age or thereafter. 14 Participants with and without TBI were eligible if they had (i) sufficient cognitive capacity, general medical health, and English language skills to complete the study measures, (ii) no severe psychiatric disturbances or chronic substance abuse, and (iii) no premorbid neurological conditions (TBI participants) or no other neurological conditions (non-TBI participants).
To assess potential recruitment bias, we compared key characteristics between our study sample and the broader longitudinal head injury outcome study participants (n = 3075). Participants in the current study did not differ significantly in age at injury, current age, post-traumatic amnesia (PTA), or Glasgow Coma Scale (GCS) scores, but had higher educational attainment. Age-stratified analyses revealed the participants in their 40s, 50s, and 80s had higher educational attainment (all P < .009), whereas no difference was observed for participants in their 60s or 70s. Among participants in their 60s and 70s, those in the current sample had sustained their injuries at a significantly older age (P = .003 and .012, respectively). Participants in their 40s had more severe injuries, indicated by longer PTA duration (P = .049), and lower GCS scores (P = .003). Detailed sample comparisons are provided in Supplemental Table 1.
Measures
Injury-related data were obtained from medical records. PTA duration was acquired using the Westmead Post Traumatic Amnesia Scale. 27 Demographic information was collected via research interviews and online questionnaires. Premorbid intellectual functioning (Intelligence Quotient) was estimated using the Wechsler Test of Adult Reading, 28 which has been identified as an appropriate measure for estimating premorbid IQ and proxy for cognitive reserve in TBI.29,30
Cognitive Measures
We assessed cognitive function using a comprehensive neuropsychological test battery common standardised measures designed to capture all major cognitive domains: information processing speed (WAIS-III Digit Symbol Coding), 31 basic attention and working memory (WAIS-III Digit Span Forwards and Backwards, respectively), 31 verbal learning and memory of a wordlist (Rey Auditory Verbal Learning Test), 32 verbal learning and memory of short stories (WMS-R Logical Memory), 33 visual processing and memory (Rey Complex Figure), 34 phonemic and semantic fluency (Controlled Oral Word Association Test), 35 divided attention (Trail Making Test), 36 and inhibition (Hayling Sentence Completion Test). 37
MRI Acquisition
Imaging was performed using a Siemens Magnetom Skyra 3T scanner (Erlangen, Germany). T1-weighted scans were obtained using these parameters: resolution = 1 mm × 1 mm × 1 mm, repetition time = 2300 ms, echo time = 3.65 ms, field of view = 208 mm × 240 mm × 256 mm, flip angle = 9°, number of slices = 208 slices, slice thickness = 1 mm. The diffusion-weighted sequence was acquired with: resolution = 2.5 mm × 2.5 mm × 2.5 mm, 71 diffusion-weighted images (b = 3000 s/mm2) and 1 nondiffusion-weighted image, repetition time = 8400 ms, echo time = 110 ms, field of view = 150 mm × 240 mm × 240 mm, and number of axial slices = 59. A reverse-phase image was obtained for distortion correction.
Structural Imaging Pre-Processing and Analysis
FreeSurfer version 6.038 was used to estimate cortical thickness. To identify damaged brain regions in individuals with TBI, lesion masks were manually created on T1-weighted MRI images. Lesion masks were then processed through the Virtual Brain Grafting (VBG) package. 39 VBG works by replacing damaged areas with a synthetic ‘virtual brain’, which helps achieve more accurate cortical parcellation by reducing errors from tissue discontinuities or loss. This preprocessing enhanced the reliability of later segmentation and parcellation. The Destrieux atlas in FreeSurfer was used for cortical parcellation. Cortical thickness was measured for each region. Subcortical volumes were also extracted.
Cortical or subcortical regions with partial lesion overlap underwent additional adjustment, whereby we calculated the percentage of lesion overlap as:
Including this correction enabled reliable estimation of cortical and subcortical structure values while reducing lesion-related segmentation errors. It also improved result comparability across participants with different lesion patterns and sizes, strengthening the statistical analysis.
Diffusion Tensor Imaging Pre-Processing and Analysis
Diffusion-weighted imaging data was preprocessed using QSIPrep (version 0.21.4), 40 an automated pipeline specialised for diffusion MRI processing. This involved denoising the raw images, correcting for Gibbs ringing artefacts, and adjusting for motion and eddy currents using FSL’s eddy tool. Reverse phase-encoded b0 images were used to correct susceptibility-induced distortions, and bias field correction was used to address image intensity inhomogeneities across the images.
QSIRecon was then used to reconstruct fibre orientation distributions (FODs), using a single-shell, 3-tissue pipeline for robust modelling of tissue microstructure. Response functions for white matter, grey matter, and cerebrospinal fluid were estimated using a method like the ‘Dhollander’ approach. 41 These tissue-specific response functions were used in the multi-tissue constrained spherical deconvolution algorithm to generate FODs. The accuracy of FOD estimation was enhanced by incorporating global intensity normalisation and calibration steps.
An unbiased, study-specific white matter FOD template was created from 50 TBI participants and 50 controls, with equal gender distribution, using non-linear registration of white matter FODs. Individual FODs were registered to this template, with fixels segmented and reoriented to align accordingly.
This template was then used to perform whole-brain probabilistic tractography to allow connectivity-based fixel enhancement for statistical analysis. The iFOD2 algorithm was used to initially generate 20 million streamlines, which were filtered to 2 million using spherical-deconvolution informed filtering of tractograms to reduce reconstruction biases. The produced fixel metrics including fibre density (FD), log-transformed fibre cross-section (log-fc), and fibre-bundle density and cross-section (FDC), representing micro- and macro-structural properties within white matter fibre bundles, along with traditional diffusion tensor metrics (fractional anisotropy, apparent diffusion coefficient, axial diffusivity, and radial diffusivity).
TractSeg, a deep learning framework, was used to segment the FOD template into 72 predefined white matter tracts. 42 TractSeg is a feasible automated tract segmentation in TBI. 43 Average fixel-based and tensor-based metrics were calculated for each participant across these tracts, representing a detailed assessment of white matter integrity across microstructure and macrostructure.
Statistical Analysis
Statistical analyses were conducted in R (version 4.4.2). Non-TBI controls were selected from a larger sample recruited from the general community using cardinality matching (tolerance = 0.2) to algorithmically select the subset of controls that best matched the TBI group on age and gender. 44 We assessed the success of group matching by comparing groups on demographic variables: t-tests for continuous variables (current age, years of education, and premorbid IQ) and chi-squared tests for categorical variables (gender and age strata by decade: 40-49, 50-59, 60-69, 70-79, and 80-89). Fisher’s exact test was used for primary language (English vs non-English) and ethnicity (White vs non-White) due to the low prevalence of participants who primarily spoke a language other than English or identified with an ethnic background other than White. The association between age and injury variables (time since injury and injury severity) in the TBI group was assessed using linear regression.
For our primary analysis, we conducted linear regressions to assess whether age-related differences in cortical thickness, subcortical volume, and white matter parameters differed between TBI and non-TBI control groups. Individual associations between age or TBI and specific brain structures have been previously documented and were not the focus of this study.11 -14,45 Nevertheless, these individual associations are presented in Supplemental Figure 1. The interaction between age and group (TBI vs Control) determined whether the effect of TBI on brain structures varied across age at assessment. Continuous variables were standardised to provide effect sizes (standardised beta coefficients), interpreted as small (<.29), medium (.30-.49), or large effects (>.50). 46 Corrections for multiple comparisons were conducted following the regressions by controlling the false discovery rate (FDR) at P < .05. 47 Where significant interactions emerged following FDR correction, pairwise comparisons of estimated marginal means between each age decade and group were conducted. Marginal means were calculated at the median of each decade: ages 45, 55, 65, 75, and 85 years for the respective age ranges 40 to 49, 50 to 59, 60 to 69, 70 to 79, and 80 to 89 years. Gender, premorbid IQ, and total intracranial volume were covariates in all regression models.
Finally, to explore whether changes in brain structure with ageing were associated with cognitive function in the chronic period after TBI, linear regressions were performed to examine the relationship between structural integrity and cognitive test scores in the TBI group, controlling for age as an additional covariate. Again, corrections for multiple comparisons were conducted following the regressions by controlling the FDR at P < .05. For white matter tracts, effect sizes are reported for FDC, a metric of overall tract integrity using both microstructural and macrostructural properties, given its sensitivity to changes after acquired brain injury.48,49 Regression results for all other white matter metrics are provided in figures.
To address potential concerns around statistical power in the smallest age strata and increase confidence in our results, a sensitivity analysis was conducted with the 70 to 79 and 80 to 90 years age groups condensed into a single 70 to 90 stratum (median = 80). This condensation affected only the pairwise comparisons of estimated marginal means, as the primary analyses of main effects and interactions retained age as a continuous variable and were therefore unchanged. Results were largely consistent with the primary analyses and are presented in Supplemental Results 1.
Data Availability
De-identified scans from this study will be made available in de-identified format to researchers via the Federal Interagency Traumatic Brain Injury Research database (https://fitbir.nih.gov/). De-identified data from this study are not available in a public archive. Other de-identified data from this study are not available in a public archive and will be made available (as allowable according to institutional IRB standards) by emailing the corresponding author as of June 2025.
Results
Complete sample demographic and injury characteristics are provided in Table 1. After matching, the sample included 123 participants with TBI and 74 non-TBI controls aged between 40 and 90 years. The TBI group was 6 to 33 years post-injury (M = 21.35, standard deviation = 6.97). Older aged TBI participants had shorter time since injury (B = −0.19, P < .001, 95% confidence interval [CI] [−0.29, −0.09]), while injury severity was not related to age (B = −0.44, P = .07, 95% CI [−0.91, 0.03]). The TBI and non-TBI groups did not significantly differ on age at assessment, gender, primary language, or ethnicity after matching. However, non-TBI controls demonstrated higher premorbid IQ and educational attainment (Table 1). Consequently, premorbid IQ was included as a covariate in all analyses, and a sensitivity analysis identical to the primary analysis was conducted with a smaller sample of non-TBI controls who were matched to the TBI group on age, gender, premorbid IQ, and educational attainment. The results of this sensitivity analysis are provided in Supplemental Results 2.
Demographic and Injury Characteristics for Participants.
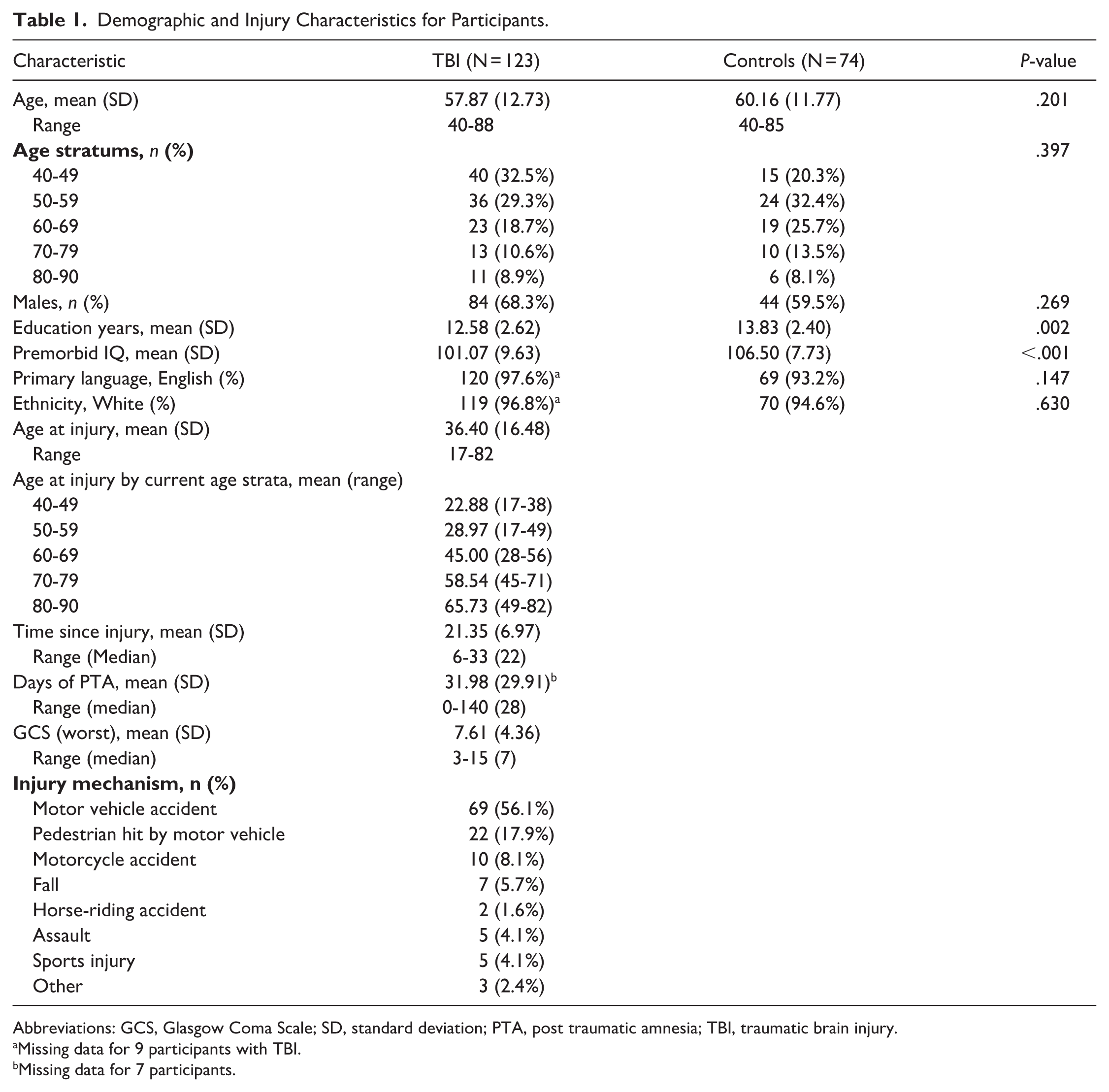
Abbreviations: GCS, Glasgow Coma Scale; SD, standard deviation; PTA, post traumatic amnesia; TBI, traumatic brain injury.
Missing data for 9 participants with TBI.
Missing data for 7 participants.
TBI Reduces Structural Integrity
We first used linear regression to examine group differences in structural integrity, regardless of age. The TBI group had significantly reduced integrity in all 72 tracts in TractSeg, 42 as well as reduced volume in 12 of 14 subcortical structures and reduced cortical thickness 13 of 117 cortical structures compared to non-TBI controls (Supplemental Figure 1).
Structural Integrity Decreases With Ageing
We then examined changes in structural integrity related to age. Across both groups, linear regression revealed that older age was associated with significantly reduced integrity in all 72 tracts in TractSeg, 42 as well as reduced volume in 11 of 14 subcortical structures, and reduced cortical thickness in 44 of 117 cortical structures (Supplemental Figure 1).
Changes to Ageing in the Brain After TBI
We then investigated whether age-related differences in structural integrity varied between the TBI and non-TBI groups. There was a significant age × group interaction for the left and right fornix (β = .20, P = .020, 95% CI [0.09, 0.30] and β = .23, P = 0.033, 95% CI [0.09, 0.36], respectively), and the left thalamus (β = .17, P = .046, 95% CI [0.06, 0.29]; Figure 1). Pairwise comparison revealed volume in the left thalamus was significantly reduced for individuals with TBI in their 40s, 50s, and 60s compared to their same-aged peers without TBI, however there was no significant difference between groups in their 70s and 80s (Table 2). The left and right fornix showed significantly reduced integrity for individuals with TBI in their 40s, 50s, 60s, and 70s, but not individuals in their 80s, compared to their same-aged peers without TBI (Table 2; Figure 1). Interaction effects in the fornix were evident across multiple metrics of microstructural and macrostructural integrity (Figure 2). Further interactions were observed in a range of cortical and subcortical structures; however, these were not significant after applying FDR correction for multiple comparisons (Figure 2). Specifically, no other white matter tracts showed this interaction, whereas uncorrected interactions were observed in multiple cortical thickness regions across both hemispheres and in subcortical structures including the left amygdala, caudate, hippocampus, and putamen, and the right pallidum and thalamus.

Age-specific fornix and thalamic structural changes post-TBI show reductions in middle-ages but not accelerated ageing after injury. (A) Scatter plots showing the interactions between age and group (TBI vs controls) on left fornix, right fornix, and left thalamus, significant after false discovery rate (FDR) correction for multiple comparisons. Individuals with TBI show reduced left thalamic volume compared to non-TBI controls aged 40 to 69, but do not differ from controls at ages 70 to 90. Fornix integrity was significantly reduced in individuals with TBI in their 40s to 70s compared to non-TBI controls, with no differences observed in the 80s age group. Results from FDC are plotted to illustrate the relationship and other white matter metrics followed a similar pattern. (B) Brain regions with significant interaction effects visualised in coronal (top) and sagittal (bottom) planes: left fornix (orange), right fornix (purple), and left thalamus (blue).
Estimated Marginal Means (SE) by Age and Pairwise Comparisons for Age X Group Significant Interactions.
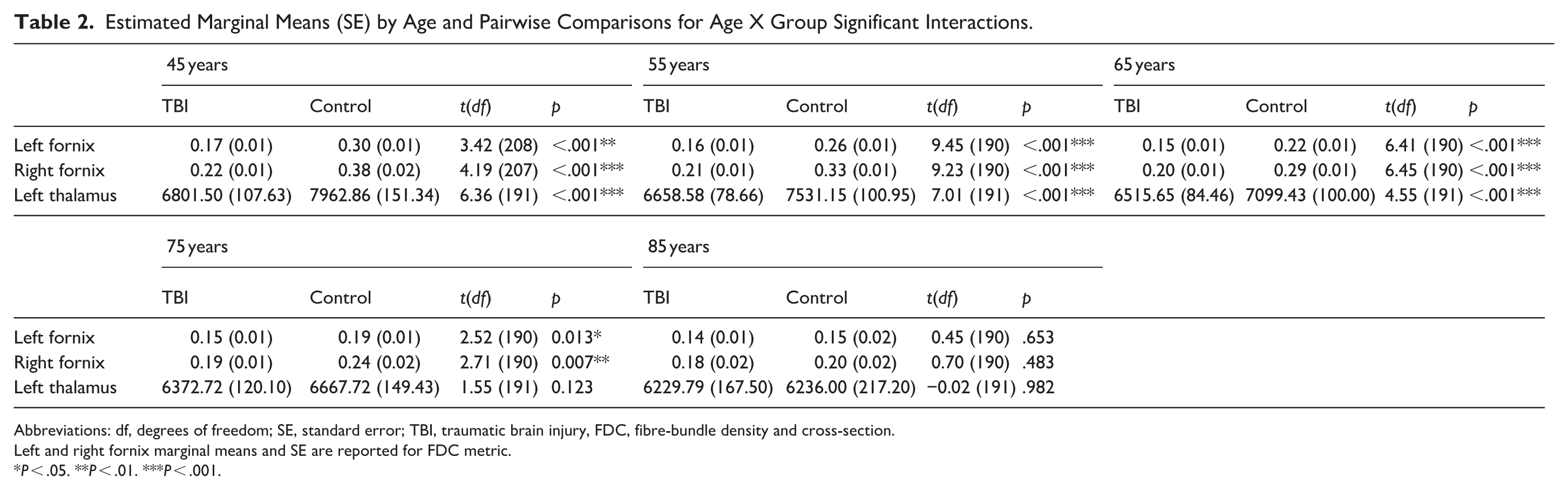
Abbreviations: df, degrees of freedom; SE, standard error; TBI, traumatic brain injury, FDC, fibre-bundle density and cross-section.
Left and right fornix marginal means and SE are reported for FDC metric.
P < .05. **P < .01. ***P < .001.

Age-dependent vulnerability following TBI is region-specific. Summary of age × group interactions from regressions across all (A) white matter tracts and integrity metrics, (B) cortical regions, and (C) subcortical structures. Gender, total intracranial brain volume, and premorbid IQ were covariates. Uncorrected P-values and false discovery rate (FDR)-corrected P-values are provided and denoted by small Fand large circles, respectively. Multiple regions show age-dependent vulnerability following TBI, with the bilateral fornix tracts and left thalamus demonstrating robust significant associations after correction for multiple comparisons. Red circles signify positive associations and blue circles signify negative associations.
Structural Integrity Correlates With Cognition
Next, we used linear regression to assess whether the structures that differed with age in TBI (ie, showed a significant interaction after FDR correction) were related to cognitive function in the TBI group. The characteristics and distribution of scores on cognitive assessment in this cohort have been previously reported. 20 Right fornix integrity was associated with all cognitive domains except auditory attention and immediate visual memory. Left fornix integrity showed similar associations but was additionally not associated with working memory or visuospatial processing. Left thalamic volume was associated with 6 cognitive domains (Figure 3).

Reduced fornix integrity and left thalamic volume are associated with cognitive impairment after TBI. (A) Significant associations were found between left (top) and right (bottom) fornix integrity and cognitive performance after TBI in all domains except basic auditory attentional capacity and immediate visual memory (left and right fornix), and working memory and visuospatial processing (left fornix). Results were largely consistent across metrics of white matter integrity (y-axis). Red circles signify positive associations and blue circles signify negative associations. (B) Significant associations between left (orange) and right (purple) fornix integrity and cognitive performance after TBI. Results from FDC are plotted to illustrate relationships and other white matter metrics followed a similar pattern. (C) Significant associations between cognitive performance after TBI and left thalamus volume (blue) were found for 6 cognitive domains.
Left Fornix
Following TBI, lower FDC in the left fornix was associated with poorer processing speed (β = 1.06, P < .001, 95% CI [0.69, 1.43]), acquisition and delayed recall of verbal stories (β = 0.87, P = .002, 95% CI [0.43, 1.30] and β = .67, P = .038, 95% CI [0.23, 1.12], respectively), acquisition and delayed recall of a verbal wordlist (β = 1.10, P < .001, 95% CI [0.69, 1.51] and β = 1.03, P < .001, 95% CI [0.62, 1.45], respectively), delayed visual memory (β = .78, P = .005, 95% CI [0.35, 1.21]), verbal fluency (phonemic and semantic; β = .84, P = .003, 95% CI [0.37, 1.31] and β = .65, P = .022, 95% CI [0.19, 1.10], respectively), divided attention (β = .99, P = .008, 95% CI [0.57, 1.41]), and inhibition (β = .71, P = .017, 95% CI [0.26, 1.16]).
Right Fornix
In individuals with TBI, lower FDC in the right fornix was relate to poorer working memory (β = 0.53, P = .036, 95%CI [0.18, 0.89]), processing speed (β = .92, P < .001, 95% CI [0.62, 1.21]), acquisition and delayed recall of verbal stories (β = .84, P < .001, 95% CI [0.50, 1.18] and β = .71, P = .002, 95% CI [0.36, 1.06], respectively), acquisition and delayed recall of a verbal wordlist (β = .93, P < .001, 95% CI [0.61, 1.26] and β = .77, P < .001, 95% CI [0.44, 1.11], respectively), visuospatial processing (β = .50, P = .041, 95% CI [0.15, 0.85]), delayed visual memory (β = .67, P = .003, 95% CI [0.33, 1.01]), verbal fluency (phonemic and semantic; β = .64, P = .009, 95% CI [0.28, 0.99] and β = .51, P = .029, 95% CI [0.16, 0.86], respectively), divided attention (β = -.59, P = .006, 95% CI [-0.93, -0.25]), and inhibition (β = -.61, P = .004, 95% CI [-0.85, -0.36]).
Left Thalamus
Following TBI, reduced volume in the left thalamus was associated with poorer processing speed (β = .637, P = .002, 95% CI [0.27, 1.00]), acquisition of verbal stories (β = .593, P = .026, 95% CI [0.18, 1.00]), acquisition and delayed recall of a verbal wordlist (β = .734, P = .002, 95% CI [0.33, 1.13] and β = .610, P = .019, 95% CI [0.20, 1.02], respectively), delayed visual memory (β = .492, P = .046, 95% CI [0.09, 0.90]), and semantic verbal fluency (β = .517, P = .030, 95% CI [0.12, 0.92]). Integrity of the left thalamus was not related to auditory attentional capacity, working memory, delayed recall of verbal stories, visuospatial processing, visual learning, phonemic fluency, divided attention, or inhibition.
Discussion
This study examined the interactive effects of older age and moderate–severe TBI on cortical thickness, subcortical volume, and white matter tract integrity using a non-TBI control group for comparison. As expected, when examined separately, both older age and TBI were associated with reduced tract integrity, subcortical volume and cortical thickness, consistent with previous findings. 50 Alterations to typical age-related structural declines were observed after TBI in the bilateral fornix and left thalamus. Contrary to our hypothesis, however, we did not find a more pronounced structural loss in older adults with TBI. Instead, these alterations resulted in reduced volume and integrity in middle-aged adults with TBI, while older adults with TBI exhibited no significant difference from age-matched controls. Our findings, while cross-sectional, revealed no group-level evidence to support an accelerated ageing process in the brain after TBI.
We did not find a steepened trajectory of structural loss with older age in any grey or white matter region in the TBI group. Rather, we found a more nuanced pattern where TBI may cause an initial, premature decrease in structural integrity that resembles the loss reached in normal ageing, but which does not continue to compound with normal ageing processes. Though we note this pattern was observed only in the bilateral fornix and left thalamus. While many other structures exhibited a similar interaction effect, these did not remain significant after correcting for multiple comparisons, emphasising the need for replication in other samples. We also note that the interaction in the bilateral fornix and left thalamus was only significant prior to correction for multiple comparisons in our sensitivity analysis, while all pairwise comparisons remained significant and showed the same pattern as the main analysis. Due to this, we cannot rule out that our group differences converged in older age because the non-TBI controls had greater educational attainment, a factor associated with better cognitive reserve and ageing.51,52 However, the preserved pattern of results and significant pairwise comparisons more likely suggests that the smaller sample in the sensitivity analysis was not sufficiently powered for the large number of analyses, particularly in the older age groups.
The specificity of the fornix and thalamus may be due to their vulnerability to structural change in both TBI and typical ageing. The fornix and thalamic radiations have been identified as particularly vulnerable in white matter injury following TBI. 53 Similar vulnerability has also been observed in the thalamus in TBI.13,14,54-58 Whereas in typical brain ageing, both the fornix and thalamus are highly susceptible to change, with the thalamus further implicated via significant alterations in frontostriatal circuits with ageing.59,60 This suggests these are key regions for examination in ageing with TBI and clinically may provide a better understanding and determination of individuals at risk of long-term impairments who may be in greater need of ongoing rehabilitation.
Cognitive Correlates of Altered Brain Ageing After TBI
We then examined the cognitive correlates of alterations in the fornices and left thalamus after TBI. As expected, reduced fornix integrity and thalamic volume correlated with multidomain cognitive deficits.57 -59,61,62 The strongest associations were observed for verbal memory and processing speed, with the fornix showing large effect sizes and the left thalamus showing medium effect sizes. These findings emphasise the established role of the fornix and thalamus in these cognitive functions 63 and align with our previous work, in which verbal learning and memory in chronic TBI followed a similar pattern to the present study: middle-aged adults demonstrated more pronounced impairments than older adults in the chronic period following TBI when compared to age-matched controls. 20
Together, these findings reveal age-specific vulnerability periods that could inform optimal intervention timing. Middle age potentially represents a critical window during the chronic period to implement compensatory strategies that preserve function and mitigate the structural and cognitive impacts of TBI. Meanwhile, long-term rehabilitation services for older adults with TBI should integrate relevant compensatory strategies with healthy ageing recommendations and support.66,67 This is not to suggest that chronic TBI does not add functional burden as people age, but rather to emphasise that our cohort showed no evidence of TBI accelerating or adversely affecting ageing processes in the brain.
Limitations
Study limitations should be acknowledged when interpreting these results. The study included adults aged 40 years and above, leaving uncertainty about whether reduced structural integrity in the fornix and thalamus remains stable in younger adults (20-40 years) with chronic TBI. Although future research is needed, there is no evidence suggesting age-specific structural decline during the chronic injury phase before age 40. Older adults in the present sample had shorter time since injury compared to younger participants. This likely reflects survival and recruitment biases in older adults with TBI. Individuals with more severe impairment or comorbidity burden may have died at younger ages, been excluded due to study criteria, or been less likely to participate in research, and thus our results may not generalise to these groups.66,67 This is unlikely to have contributed to the relative stability across age groups in the fornix and left thalamus after TBI, as we observed age-associated structural losses comparable to non-TBI controls across other regions.
Our cross-sectional design limited our ability to conclusively demonstrate or rule out individual trajectories of decline with ageing. Longitudinal prospective studies will be crucial for understanding how TBI and age interact over time and ongoing follow-up data may provide opportunities to validate these findings. We also recognise that the interaction pattern between age and TBI was analysed at a group level, and individuals with TBI may experience variable trajectories with time post-TBI and ageing. Additionally, we note that our study sample does not capture those (with or without TBI) experiencing structural decline attributed to underlying neurodegenerative pathologies.
Our findings should be considered within context. Individuals who sustain a TBI in countries with healthcare systems differing from Australia’s, including accessibility, chronic care management, and funded universal healthcare, may show different outcomes.68,69 Following TBI from motor vehicle-related accidents, individuals in the Australian state of Victoria receive inpatient rehabilitation and ongoing community-based support as justified under a no-fault accident compensation scheme. For non-motor vehicle related TBI, the chronic health care and insurance setting in Australia also differs. This can increase care accessibility, affordability, and continuity of chronic condition management compared to other regions.70-72 Individuals with TBI receiving this care in Australia appear to have better chronic functional and health outcomes compared to those in other systems, 73 which is likely associated with better brain health and cognitive function 74 and may suggest improved outcomes in these domains as well. This aligns with evidence that providing appropriate long-term management improves outcomes in other chronic conditions. 75
Healthcare delivery and insurance systems also play a critical role in ageing outcomes, with substantial international variability in healthcare access among ageing populations.76,77 Disparities in healthcare funding and perceived access exist even within high-incomes nations with universal healthcare, 74 significantly affecting older adults.75,76 These systemic differences may influence ageing trajectories and, consequently, ageing outcomes following TBI. The care models available in Australia may result in relatively healthy ageing with TBI by ensuring consistent access to appropriate interventions, regular monitoring, and more integrated support services. Consequently, healthcare and rehabilitation characteristics should be considered when interpreting and comparing TBI outcomes across different regions and countries.
Conclusions
This study provides important insights into the relationship between TBI and ageing, demonstrating that moderate–severe TBI does not necessarily accelerate normal age-related structural decline but rather may cause initial structural alterations that do not compound with normal ageing processes in the long-term. Our findings specifically highlight the bilateral fornix and left thalamus as regions with structural changes after TBI that mirror those eventually seen in typical ageing. Middle-aged adults with TBI showed reductions in the integrity and volume of these structures relative to age-matched controls that diminished in older adults as controls experienced typical age-related decline; a pattern distinct from other brain structures that showed consistent TBI-related reductions across all ages. These structural alterations correlated meaningfully with cognitive performance, particularly verbal memory and processing speed. While longitudinal follow-up data is required to further validate these findings, our results challenge the notion of accelerated ageing after TBI and underscore the importance of age-tailored rehabilitation strategies that address potential vulnerability periods, particularly for interventions targeting verbal memory and processing speed deficits associated with structural changes in the fornix and thalamus.
Supplemental Material
sj-pdf-1-nnr-10.1177_15459683261442198 – Supplemental material for The Relationship Between Age, Brain Structure, and Cognition in Chronic Traumatic Brain Injury
Supplemental material, sj-pdf-1-nnr-10.1177_15459683261442198 for The Relationship Between Age, Brain Structure, and Cognition in Chronic Traumatic Brain Injury by Amber Ayton, Jennie Ponsford, Amelia J. Hicks, Rachael Knott, Gavin Gan and Gershon Spitz in Neurorehabilitation and Neural Repair
Footnotes
Acknowledgements
We thank the staff and participants of the Monash-Epworth Rehabilitation Research Centre for their valuable contributions.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Health and Medical Research Council (NHMRC) under Grant APP 1127007.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplementary material for this article is available on the Neurorehabilitation & Neural Repair website along with the online version of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.