
Abstract
Background
We previously demonstrated that deep brain stimulation (DBS) of lateral cerebellar nucleus (LCN) can enhance motor recovery and functional reorganization of perilesional cortex in rodent models of stroke or TBI.
Objective
Considering the treatment-related neuroplasticity observed at the perilesional cortex, we hypothesize that chronic LCN DBS-enhanced motor recovery observed will carry-over even after DBS has been deactivated.
Methods
Here, we directly tested the enduring effects of LCN DBS in male Long Evans rats that underwent controlled cortical impact (CCI) injury targeting sensorimotor cortex opposite their dominant forepaw followed by unilateral implantation of a macroelectrode into the LCN opposite the lesion. Animals were randomized to DBS or sham treatment for 4 weeks during which the motor performance were characterize by behavioral metrics. After 4 weeks, stimulation was turned off, with assessments continuing for an additional 2 weeks. Afterward, all animals were euthanized, and tissue was harvested for further analyses.
Results
Treated animals showed significantly greater motor improvement across all behavioral metrics relative to untreated animals during the 4-week treatment, with functional gains persisting across 2-week post-treatment. This motor recovery was associated with the increase in CaMKIIα and BDNF positive cell density across perilesional cortex in treated animals.
Conclusions
LCN DBS enhanced post-TBI motor recovery, the effect of which was persisted up to 2 weeks beyond stimulation offset. Such evidence should be considered in relation to future translational efforts as, unlike typical DBS applications, treatment may only need to be provided until such time as a new function plateau is achieved.
Keywords
Introduction
Traumatic brain injury (TBI) disrupts normal brain function through primary (mechanical damage) and secondary (cascades of metabolic, cellular, and molecular processes) injury mechanisms,1,2 with chronic sequelae across sensorimotor, cognitive, and behavioral domains contributing to long-term disability.3-5 Although cognitive and affective abnormalities are more commonly thought of in relation to such injuries,4,6 persistent sensorimotor dysfunction may also occur, including impaired motor control, loss of muscle tone, balance, and coordination.7-11 Standard-of-care treatment generally involves inpatient and outpatient rehabilitation; however, recovery is often limited and many patients end up in assisted-living or residential care facilities.12,13 Given the limitations of current rehabilitation efforts, there is substantial need to identify and develop effective therapeutic approaches to promote significant and long-lasting behavioral improvements beyond what typically can be achieved.7,11,14
Neurostimulation-based approaches to enhance chronic rehabilitation typically targets surviving brain tissue, with either non-invasive or invasive techniques.15-18 Our group has previously demonstrated that deep brain stimulation (DBS) of the dentatothalamocortical (DTC) pathway enhances the excitability of spared, perilesional cerebral cortex 19 and promotes functional recovery in preclinical models of acquired brain injury.20-22 To achieve chronic DTC pathway activation, we target the lateral cerebellar nucleus (LCN; the rodent homologue of the human dentate nucleus) given its dense, glutamatergic efferent projections to both medial and ventrolateral thalamic nuclei. These projections uniquely position the LCN to influence thalamocortical interactions across widespread prefrontal, frontal, and parietal cortical circuits,23-25 areas that typically represent the perilesional zone following stroke or sensorimotor TBI. Given the link between the incidence of functional reorganization and changes in excitability across these spared cortical regions, we hypothesized that chronic upregulation of the net-excitatory DTC pathway should promote functional recovery. 26 To date, we have shown that LCN DBS modulates cerebral cortical excitability in both naïve 19 and post-stroke animals, 27 with the magnitude and persistence of the effect sensitive to stimulus frequency. Moreover, in preclinical models of acquired brain injury, we have shown that treatment-related behavioral improvements are accompanied by increased expression of long-term potentiation markers, synaptogenesis, and neurogenesis in perilesional cerebral cortex.20-22,28
Specific to TBI, our group has shown previously that LCN DBS was associated with enhancement of both motor 29 and cognitive 30 rehabilitation using a sensorimotor fluid percussion and bilateral, prefrontal controlled cortical impact (CCI) model of TBI, respectively. In both instances, however, evaluation of benefit was limited to the duration of treatment delivery and no data are available to support the long-term, post-treatment carry-over of benefits achieved with LCN DBS. Such insight has significant translational implications as it would guide the development and design of human trials, particularly with regard to determining whether DBS treatment would need to be delivered indefinitely to maintain benefit or, alternatively, could be short-term in nature and applied only until a new behavioral plateau is achieved. In light of our previous findings, which support underlying enhancement of cellular and molecular mechanisms consistent with plastic reorganization, 29 we hypothesize that treatment-related benefits will persist beyond cessation of stimulation. Here, we directly evaluate the magnitude and persistence of motor rehabilitation and cellular effects of LCN DBS using a unilateral, sensorimotor CCI TBI model in rats.
Methods and Materials
Animals
Twenty-two male Long Evans rats, at about 3- to 4-month-old, weighing 225-250 g at study onset, were housed on a 12:12, reversed light/dark cycle in a climate-controlled facility. All experiments were conducted under a protocol approved by the Institutional Animal Care and Use Committee of the Cleveland Clinic.
Experimental Design—Overview
As summarized in Figure 1A, animals underwent initial acclimation, pre-training, and baseline assessments over a period of 4 weeks (weeks 1-4). Assessments included the pasta matrix task (primary) as well as the cylinder and horizontal ladder tasks. Once baseline testing was complete, each underwent a stereotaxic-guided CCI injury centered over the sensorimotor cortex contralateral to the preferred forelimb (week 5). After a 2-week recovery period (weeks 5-6), a second surgical procedure was performed to implant a concentric, bipolar macroelectrode (MS306; Protech, TX USA) in the LCN contralateral to the CCI-lesioned hemisphere (ipsilateral to the dominant forepaw). After an additional, 1-week recovery period, assessments were repeated to quantify the post-CCI motor deficits. Data from the pasta matrix task were used to assign, pseudo-randomly (ie, severity-matching), animals to 1 of 2 experimental groups: STIM− and STIM+. Thereafter, all animals were attached to an in-cage tethering system. Electrical stimulation was activated only for those animals in the STIM+ group for the next 4 weeks (weeks 9-12). Throughout that time, pasta matrix testing continued at 3 days per week, with cylinder and horizontal ladder testing repeated during the fourth week of treatment (week 12). Once the treatment phase was complete, stimulation was turned OFF for those animals assigned to the STIM+ group. Carry-over effects were examined by continuing pasta matrix testing for an additional 2 weeks (weeks 13-14), with cylinder and horizontal ladder testing repeated again during week 14. At the end of week 14, all animals were euthanized, and brain tissue was harvested and preserved for further analysis by histology.
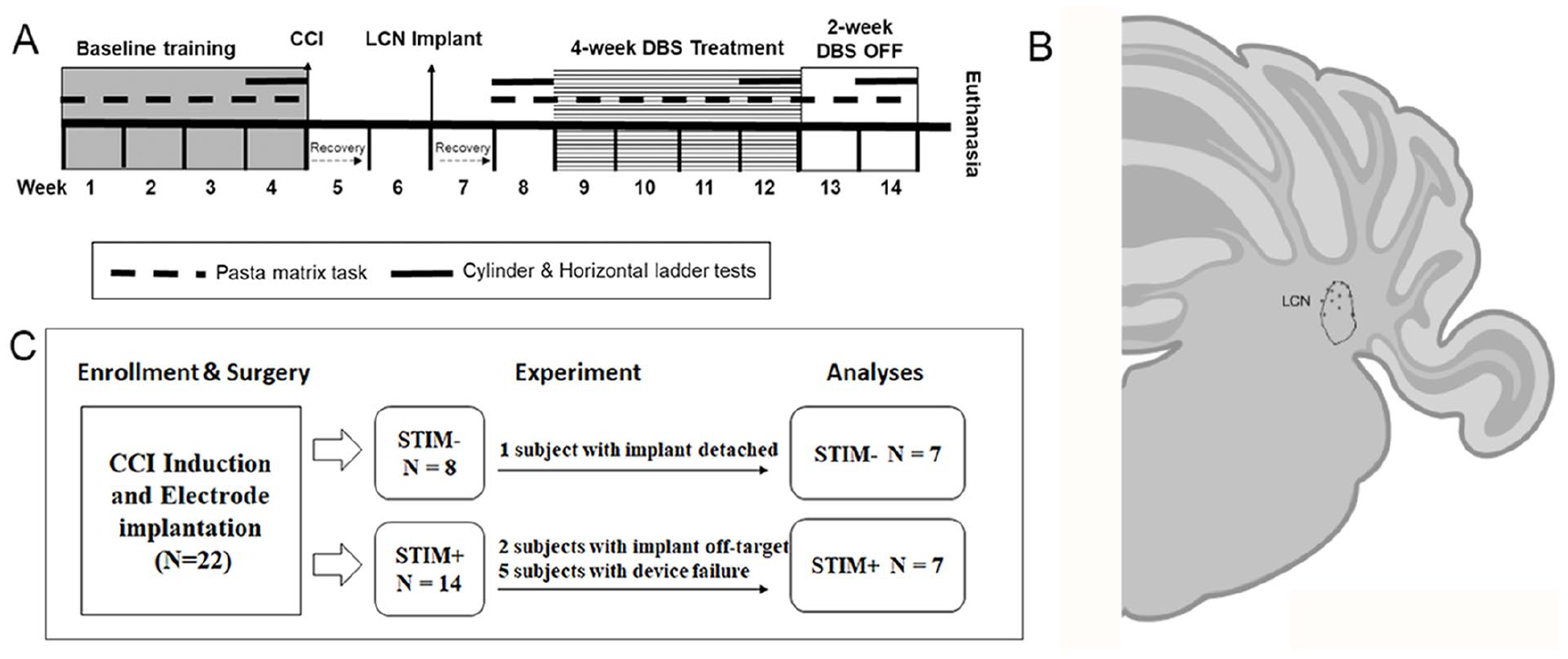
(A) Experimental timeline. (B) Electrode placement. The location of the electrode tips observed in cerebellar histology co-registered to a coronal section, created with BioRender.com, of the panel at approximately -11.28mm from Bregma. 31 (C) Elimination of experimental subjects.
Motor Assessments
All assessments were performed during the active phase of the animals’ circadian cycle under red lighting (200 Lux). Animals were food restricted at 11-12.5 g/day during task assessments to encourage performance on the food reward-based, pasta matrix task (weeks 1-4 & 8-14). The cylinder and horizontal ladder tasks were performed at 4 different time points, including the pre-CCI (naïve) baseline (week 4), 2 weeks post-CCI (week 8), during the fourth week of DBS treatment or sham (week 12) and at the end of the second week of the carry-over phase (week 14; Figure 1A).
The pasta matrix task, an index of skilled reaching and forepaw dexterity, 32 was performed once per day, 3 days per week. Briefly, a rectangular block (7.5 cm wide × 12 cm long × 2.5 cm tall) with a matrix of pre-drilled holes spaced 2 mm apart (2.5 cm deep) was pre-filled with 5 cm long pasta pieces (~1.5 mm thick) and secured to the front panel of the home cage. During each 8-minutes session, the animal was able to reach through a narrow, 1-cm wide vertical slit in order to grasp, break, and retrieve pasta pieces. Initially, pasta was placed on both sides of the matrix relative to the vertical slit and forepaw use preference was established by comparing the left- versus right-side retrieval numbers for each animal. Thereafter, pasta was loaded only on the side accessible using the dominant forepaw. Animals that failed to break a minimum of 15 pieces of pasta at behavioral plateau (week 4) were removed from the study (training failure). The pre-CCI baseline (# of pieces broken) was obtained following behavioral plateau (week 4), prior to the CCI induction, while the post-CCI, pre-treatment baseline was derived from data collected 1 week after the electrode implantation. Testing continued throughout the 4-week DBS treatment phase with repeat evaluations performed once per week during the 2-week, carry-over phase. The average performance per week was documented.
The cylinder test is used to quantify spontaneous forepaw use during rearing. 33 Briefly, the animal was removed from the home cage and placed in a high-walled cylinder for 8 minutes. Spontaneous rearing activity was recorded using a camcorder mounted above the cylinder. An operator who was blinded to group assignment recorded the number of either left or right forepaw touches against the wall of the cylinder during rearing. Forepaw preference was indexed as:
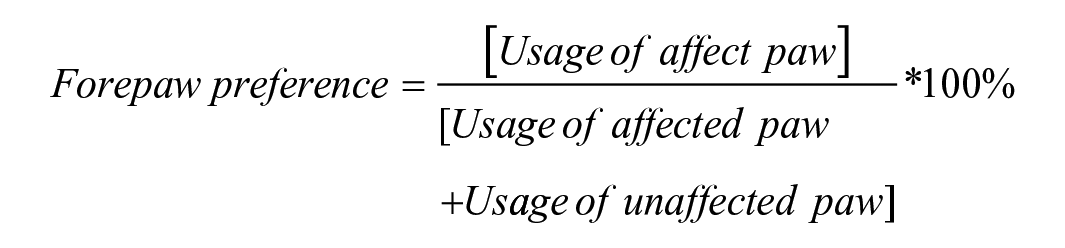
The horizontal ladder test, slightly modified from a previous report, 34 was used to evaluate motor coordination of each of the 4 limbs during walking. During testing, the animal was placed at 1 end of a 60 cm long horizontal ladder with random inter-rung spacing (from 1 to 2 cm). Each animal was required to traverse the ladder in a single direction a total of 4 times per session (the first 3 as acclimation trials, with data derived only from the final pass). Crossings were videotaped for off-line analysis, during which a blinded reviewer recorded the number of fore- and hind-paw misplacements. Misplacement of paw was defined as a failure of the palmer surface to contact the rungs combined with being grasped on the rung with the digits and palmer surface of the paw, or the paws slipped through the area between the rungs.
CCI Induction
Animals were anesthetized using ketamine (50 mg/kg, ip) and dexmedetomidine (0.15 mg/kg) followed by placement in a stereotaxic frame (Kopf Instruments, Tujunga, CA USA). The scalp was infiltrated with 0.25% bupivacaine followed by a 1-inch midline incision. A 6.0 mm in diameter circular craniotomy was created over the motor cortex opposite to the dominant forepaw (centered at AP: +1.0 mm; ML: ±3.0 mm, relative to bregma). The 5 mm-diameter impactor of the CCI system (Impact One, Leica, IL) was placed on the surface of the sensorimotor without depressing brain tissue. The parameters for impact were pre-set (impact speed: 2.25 m/s, depth advanced 2.5 mm and dwell time 0.5 seconds) and applied uniformly across animals. After CCI induction, the craniotomy was covered with sterile cellulose paper and surgical mesh, then secured using VetBond. The scalp wound was sutured, and anesthesia was reversed by injection of atipemezole (1 mg/kg, i.p.). Post-operative care included routine administration of buprenorphine (0.05 mg/kg, s.c.; 2 days post-surgery) and cephazolin (15 mg/kg, s.c.) accompanied by regular monitoring for weight loss and signs of pain and distress.
Electrode Implantation
Two weeks after the CCI induction, all animals underwent a second surgery for insertion of the LCN DBS electrode (MS306, Protech, VA, USA). Briefly, using the same protocol described above, animals were anesthetized and secured in the stereotaxic frame. The scalp wound was reopened and a bur hole created for the insertion of the electrodes into the contralesional LCN (AP: −2.30 mm; ML: ±3.6 mm; and DV: +3.7 mm; relative to the interaural point). Once in position, the electrode was secured by dental acrylic and 4 stainless screws anchoring were mounted into the exposed calvaria to serve as anchor points. Post-operative care was identical to that described for the CCI procedure above.
Post-CCI Baseline and Randomization
Two weeks after electrode implantation, all animals underwent behavioral testing to determine eligibility for continuation, facilitate group assignment, and establish the post-CCI, motor function baseline. As per our prior studies, it was established that animals failing to demonstrate at least a 30% reduction on the pasta matrix testing were considered lesioning failures and were withdrawn from the study, however all animals enrolled in the current study exceeded that threshold and were retained. Thereafter, group assignment was determined by severity-matching of the post-CCI performance on the pasta matrix task (pseudo-randomization).
LCN DBS Titration, Treatment Phase and Carry-Over Phase
Immediately prior to the start of the treatment phase, electrode impedance was tested to ensure electrical integrity. Animals with impedance values greater than 50 kΩ were removed from the study. Thereafter, the LCN DBS-induced motor side-effect threshold was established through visual observation by an experienced investigator as described previously.21,30 Briefly, 30 Hz DBS was delivered in the form of charge-balanced, square wave pulses (pulse width: 400 µs) using a constant-current biphasic stimulus isolation unit (SIU; Model BSI-2A, Microprobes, MD) controlled by a stimulator (Master-9, AMPI, Israel). The pulse amplitude was increase by 10-20 µA and the action of the paws ipsilateral to the implanted electrodes were observed. The increase of pulse amplitude was stopped when the paws were twitching or shaking, which was at the motor threshold. Pulse amplitude was individually set for each animal at 80% of the value of motor threshold. At the start of the treatment phase (week 9), animals in the STIM+ group received DBS for 12 hours daily during the active phase of the dark/light cycle. STIM− animals were similarly tethered to the commutator but no stimulation was delivered. The integrity of the electrodes and cables was regularly monitored through periodic impedance checks. Following the 4-week treatment phase, animals in the STIM+ group had their SIUs turned off, but still remained tethered, for the 2-week carry-over phase.
Euthanasia and Tissue Processing
At the end of the carry-over phase, all animals were euthanized with 100 mg/kg pentobarbital (i.p.) followed by transcardiac infusion of cold normal PBS (1X) and cold 4% paraformaldehyde. The brains were harvested and fixed for 24 hours before cryoprotection in 30% sucrose solution in PBS. Brain blocks were initially stored at −80°C followed by cryostat sectioning of cortical and cerebellar tissue in 30 µm sections collected onto gelatin-coated slides.
Volume measuring and validation of electrode placement—Nissl staining.
Cortical sections containing the CCI lesion at the sensorimotor cortex and cerebellar sections containing the electrode tracks were stained with 0.4% cresyl violet (buffered with 0.4M acetate) for 5 minutes followed by dehydration in ascending percentages of ethanol and defat in xylene. CCI lesion volume was measured using an in-house, MatLab-based program, SLICE. 31 The electrode tracks of each experimental animals on the cerebellar sections were examined under the bright-field microscope and the location of tips were co-registered onto a rat brain atlas 35 (Figure 1B).
Immunohistochemistry (IHC)
To characterize potential LCN DBS-mediated changes in perilesional cortex, which was defined as the area 100 µm away from the border of the CCI lesion core, locating at the residual sensory and secondary motor cortical regions post-CCI, we used IHC techniques to compare the expression of BDNF and CaMKIIα between treated and untreated cohorts. Briefly, the cerebral sections were treated with goat serum containing blocking buffer after the citrate-based antigen retrieval and peroxidase quenching. Cerebral sections were then incubated with the primary antibodies for BDNF (n = 5 per animal; rabbit anti-BDNF, 1:500; Abcam, MA) or CaMKIIα (n = 5 per animal; rabbit anti-CaMKIIα; 1:500; MilliporeSigma, MA) overnight at 4°C. The secondary antibody, biotinylated anti-rabbit IgG, was incubated for 1 hour followed by the incubation of the avidin-biotin complex for an additional 1 hour. The immunocomplex was visualized by developing into brown color in diaminobenzene (DAB) reaction. After mounting the coverslips, the IHC was analyzed under a bright-field microscope.
Image Analysis
The 10X images of the perilesional area on the immunostained sections were taken with a bright-field microscope (DM6 B, Leica). The numbers of the BDNF and CaMKIIα positive cells at a pre-defined area (approximately 100 × 100 µm), within 100 µm from the borders of the lesion core, were manually counted. A minimum of 5 sections was analyzed for each subject by an individual blinded to the group assignment. The data are presented as density of cells ±S.E.M. (/µm2).
Statistics Analyses
Independent samples t-tests were used to confirm that pre-treatment performance on pasta matrix, cylinder, and ladder rung walking tasks was not significantly different between the STIM+ and STIM− groups. Two random intercepts mixed effects models were then applied to test for group differences in performance for each behavioral task—1 including post-CCI baseline through treatment week 4 data, and 1 including post-CCI baseline through treatment week 6 data. An unstructured covariance matrix was implemented and time was treated as linear for all models. Subject and time point were considered random effects, while treatment group was a fixed effect. A difference in group rate of behavioral task improvement over time was tested with a time-by-group interaction term in each of the models. Analyses were conducted using SAS Studio. P values of <.05 were considered significant.
The density of BDNF and CaMKIIα positive cells in both STIM− and STIM+ groups were compared using 1-way ANOVA and Bonferroni’s post hoc test (GraphPad Prism). P values of <.05 or .0001 were considered as statistically significant.
Results
A total of 8 animals were withdrawn over the course of the study: 1 due to premature implant failure, 2 due to off-target LCN DBS electrode placement, and an additional 5 as a result of failure of their SIUs during the course of the study. This yielded a total of 7 animals in each group for statistical analysis (Figure 1C). Impedance values across all animals ranged from 22 to 46 kΩ and final pulse amplitude in the STIM+ cohort ranged from 102 to 189 µA.
Recovery Effect of LCN DBS on the Motor Functions Following CCI
As shown in Figure 2, both groups showed an initial, 73% reduction in the number of broken pasta pieces at 4 weeks post-CCI, supporting the effect of the sensorimotor CCI on motor function. Thereafter, the mean number of broken pasta pieces in the untreated, STIM− group remained relatively stable throughout the 4-week treatment period (up to DBS Wk4). In contrast, animals in the STIM+ group showed an increase in the number of pasta pieces broken at a rate 2.2 times per week more than that observed for the STIM− group across the treatment period (P < .001, Figure 2). This culminated in a 37% increase in the number of pieces retrieved by week 4 relative to baseline, supporting an effect of LCN DBS on motor recovery.
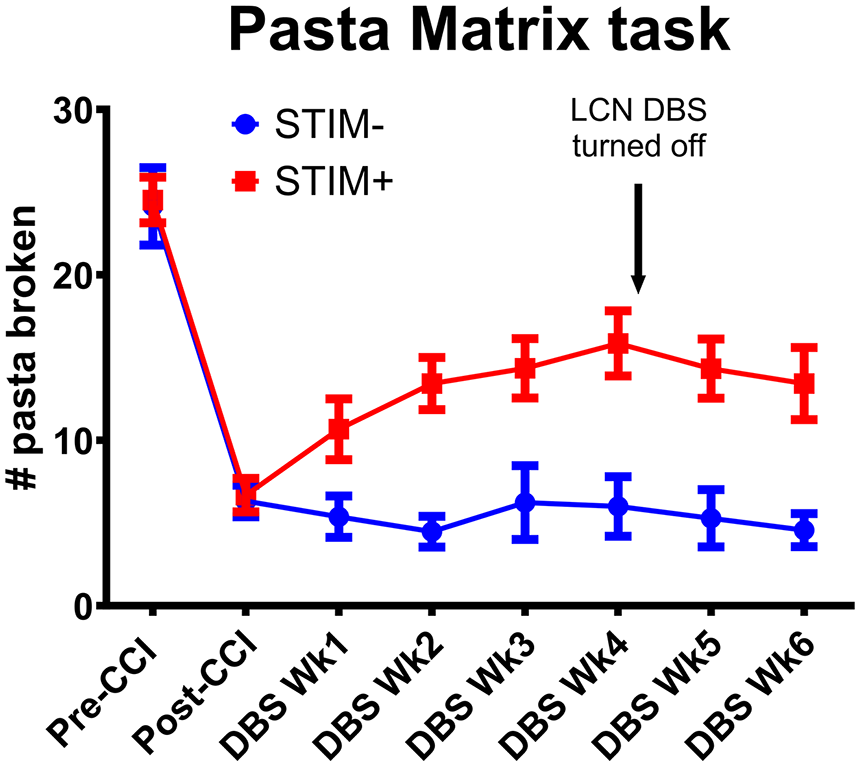
Effect of LCN DBS on post-CCI pasta matrix task performance. Following the CCI induction at the sensorimotor cortex, performance on the pasta matrix task significantly decreased for both STIM− (blue) and STIM+ (red) cohorts. After 4 weeks of LCN DBS treatment, mean performance was significantly higher in STIM+ versus STIM− animals. Following the offset of the LCN DBS, this difference in task performance persisted between cohorts (DBS Wk6). Group differences at the DBS treatment phase: P < .001 and at the carry-over phase: P < .001 (by random intercepts mixed effects models).
The above finding was supported further by similar improvements on the secondary motor metrics. As depicted in Figure 3, both groups demonstrated a marked reduction in use of the predominant forelimb post-CCI. During treatment, use of affected forelimb by animals in the STIM+ group increased by 2.5% points more per week as compared to those in the STIM− group, yielding a week 4 mean difference of 11.88% (DBS Wk4; STIM−: 33.86% vs STIM+: 45.74%, group x time interaction P < .001; Figure 3). Notably, there was no significant difference in the incidence of spontaneous rearing activity between cohorts throughout the experimental phase, as indexed by the total number of rears observed at each test session (Figure S2). Finally, CCI increased the misplacement of both the fore- and hindpaws on the horizontal ladder walking test (Figure 4). Recovery was significant in the STIM+ group, which showed reduced misplacements of the predominant forepaw in comparison to the STIM− group (DBS Wk4; STIM−: 2.86 vs STIM+: 1.71, group × time interaction: 0.36 slips per week, P = .037; Figure 4A). A similar effect was observed for hindpaw misplacements (DBS Wk4; STIM−: 1.86 vs STIM+: 1.00, group x time interaction: 0.43 slips per week, P = .040; Figure 4B).
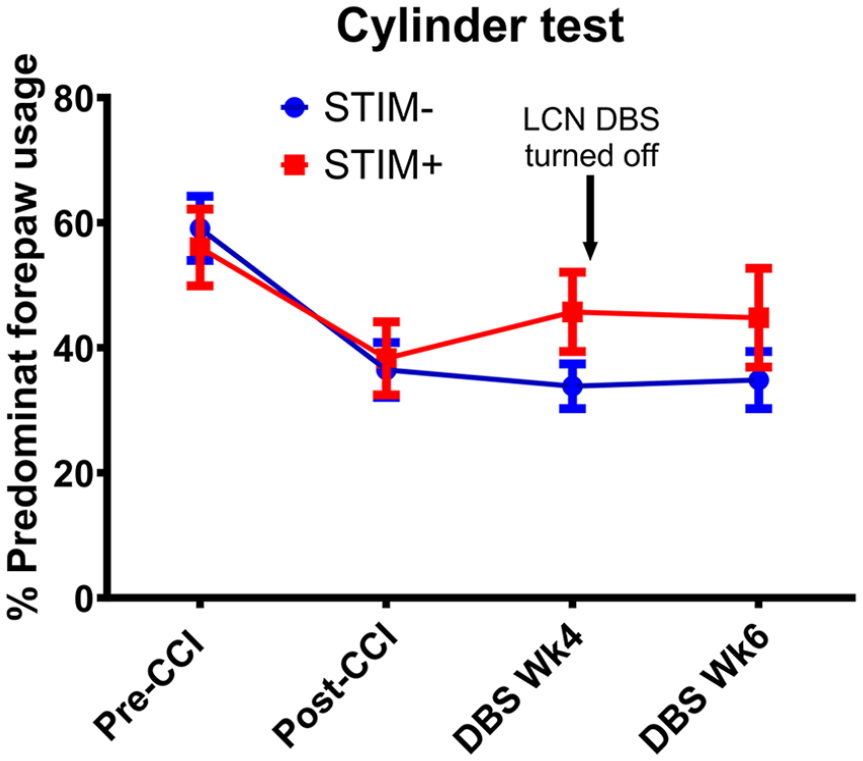
Effect of LCN DBS on spontaneous rearing behavior. CCI resulted in a decrease in dominant forepaw use in both STIM− (blue) and STIM+ (red) groups. In contrast to untreated animals, STIM+ animals showed an increase in dominant forepaw use at week 4. This effect persisted through week 6, 2 weeks after stimulation was turned OFF for the originally STIM+ animals. Group difference at the DBS treatment phase: P < .001 and at the carry-over phase: P < .001 (by random intercepts mixed effects models).
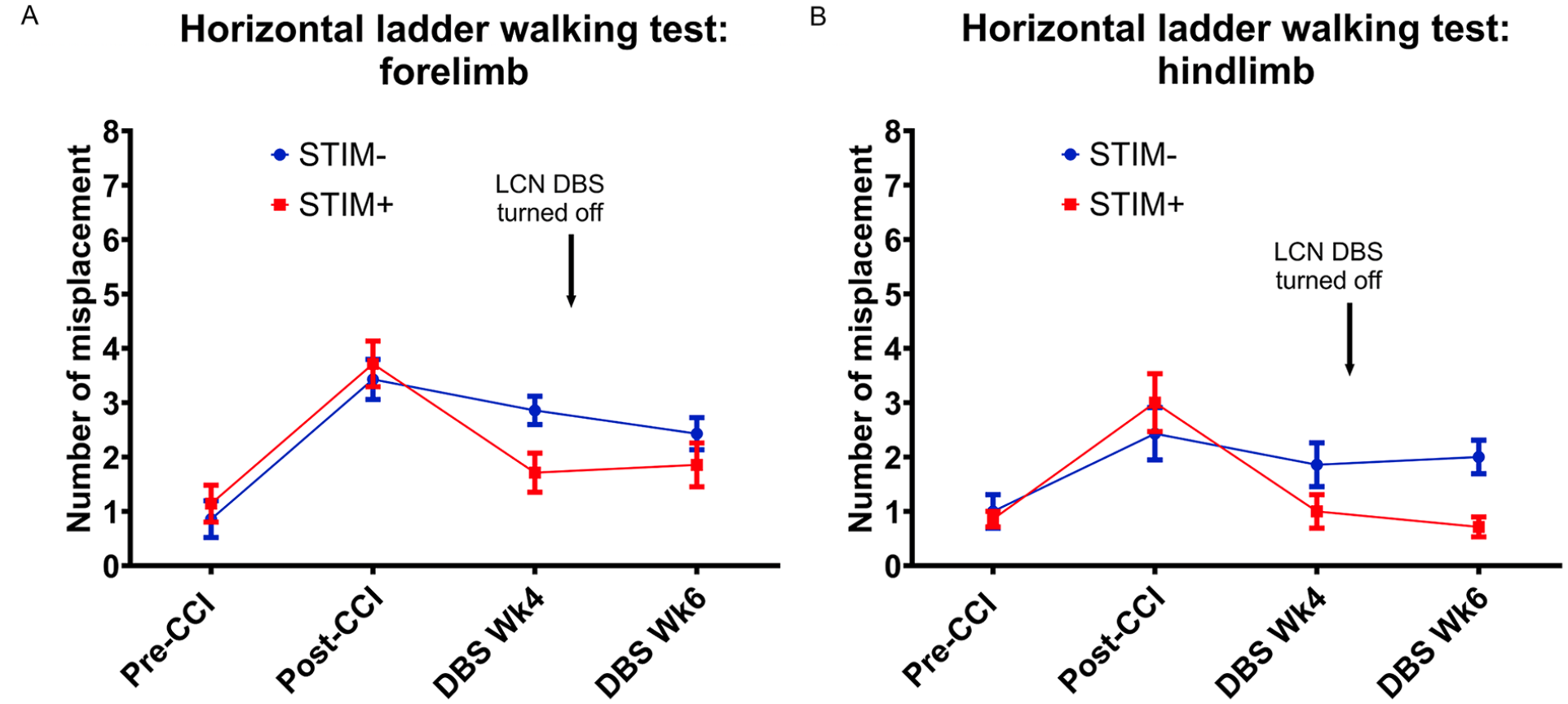
Effect of LCN DBS on post-CCI horizontal ladder walking. Post-CCI animals showed a marked increase in the number of forelimb (A) and hindlimb (B) misplacements for the dominant hemibody. Although both cohorts showed a trend towards improved performance by week 4, the effect only achieved significance for the STIM+ cohort. Following DBS deactivation (STIM+ DBS Wk6), the improvement of hindlimbs was still significant while that of forelimbs was not when compared to the STIM− groups. Group differences of forepaw at the DBS treatment phase: P = .037 and at the carry-over phase P = .07. Group differences of hindpaw at the DBS treatment phase: P = .040 and at the carry-over phase P = .003 (by random intercepts mixed effects models).
Carry-Over Effect of LCN DBS on the Motor Recovery
At the end of the treatment phase, stimulation was turned off for those animals assigned to the STIM+ group in order to determine if treatment-related gains would be maintained in the absence of ongoing stimulus delivery. A significant difference in pasta broken was still observed between the STIM+ and STIM− groups at DBS Wk6_carry-over, with STIM+ group breaking pasta at an average rate of 1.2 more pieces per week than the STIM− group (group × time interaction: P < .001, Figure 2). In the cylinder test, use of the predominant forelimb in the STIM+ group remained stable at the period of DBS Wk4 to Wk6_carry-over, which resulted in a rate of improvement significantly greater than that of the STIM− group from post-CCI baseline through the carry-over period. The STIM+ group increased predominant forepaw use by 1.5% points more per week on average than the STIM− group (group × time interaction: P < .001; Figure 3). Similarly, on the horizontal ladder test, the improvement in predominant hindpaw performance persisted between DBS Wk4 and Wk6_carry-over for animals in the STIM+ group, which resulted in a rate of improvement significantly higher than that in the STIM− group when measuring from post-CCI baseline through the carry-over period (group × time interaction: P = .0030, Figure 4A). However, from post-CCI baseline through the carry-over period, there was no evidence of a difference in improvement rates of affected forepaw placement in the ladder rung walking task between the STIM+ and STIM—groups. Animals in the STIM+ group committed only 0.17 fewer predominant forelimb slips on average than the STIM− group (group x time interaction: P = .070; Figure 4B).
LCN DBS-Mediated Increase of Expression of BDNF and CaMKIIα
Following the LCN DBS treatment, there was a significant increase in the density of BDNF positive cells in the perilesional cortex for animals in the STIM+ group when comparing to the STIM− group (One-way ANOVA: df = 3, F = 16.51, P < .0001, Bonferroni test: STIM+ ipsilateral: 0.55 ± 0.05/µm2 vs STIM− ipsilateral: 0.37 ± 0.07/µm2, P < .05; Figure 5A-B). Similarly, the density of CaMKIIα positive cells in the ipsilesional cortex of STIM+ animals was significantly greater than that of STIM− animals (One-way ANOVA: df = 3, F = 69.45, P < .0001, Bonferroni test: STIM+ ipsilateral: 1.40 ± 0.08/µm2 vs STIM− ipsilateral: 0.52 ± 0.04/µm2, P < .0001; Figure 6A-B). These findings support the persistence of stimulation-related increases in BDNF and CaMKIIα for at least 2 weeks following the offset of stimulation.
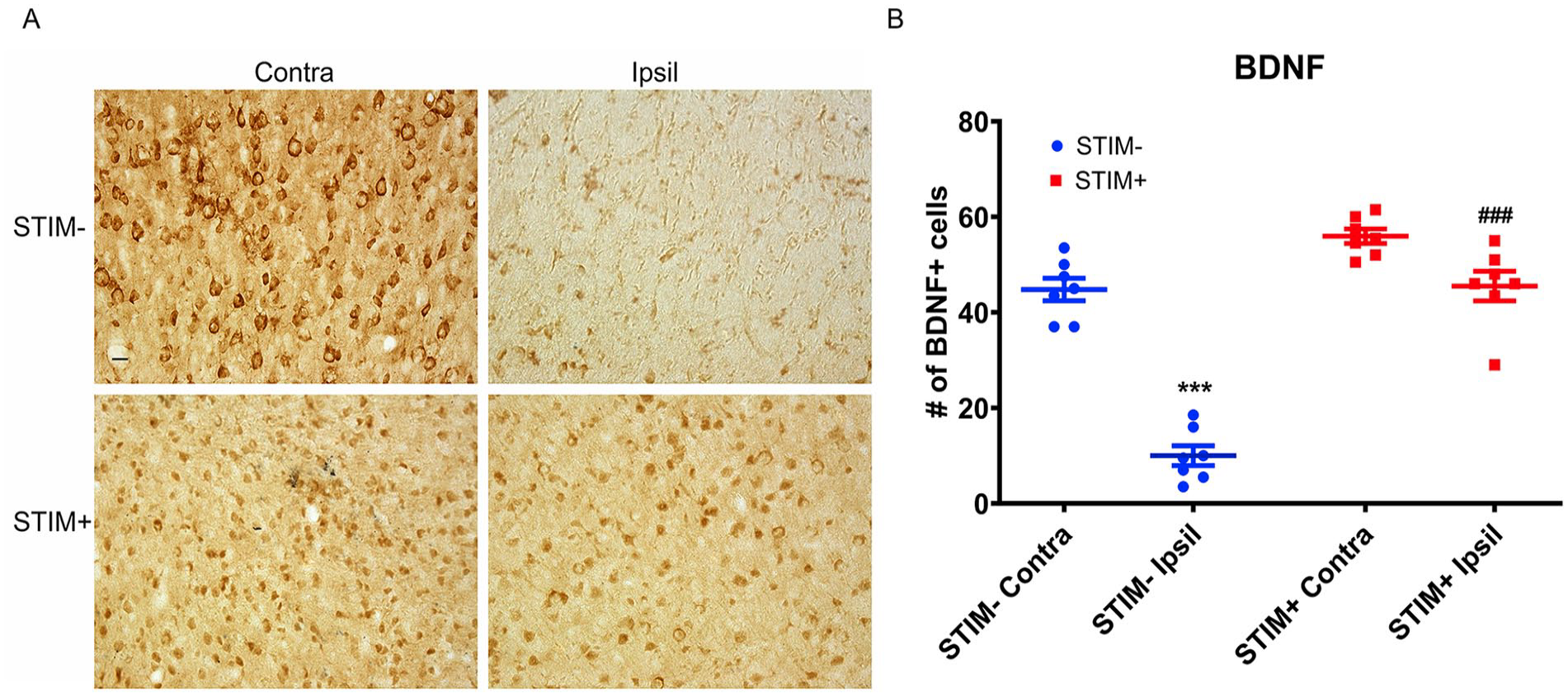
Effect of LCN DBS on BDNF-positive cell density. (A) The density of BDNF-positive cells in the ipsilesional (Ipsil), perilesional cortex of STIM+ significantly greater than that of STIM−. Data was presented as density of cells/µm2±S.E.M., analyzed by one-way ANOVA followed by Bonferroni’ post hoc test. * P < .05 when compared to the STIM− Ipsil while ### P < .0001 when compared to the STIM+ Contra. (B) The representative photomicrographs demonstrated the BDNF-positive cells in both ipsilesional (Ipsil) and contralesional (Contra) perilesional motor cortex of STIM− and STIM+ animals. Scale bar is 100 µm.
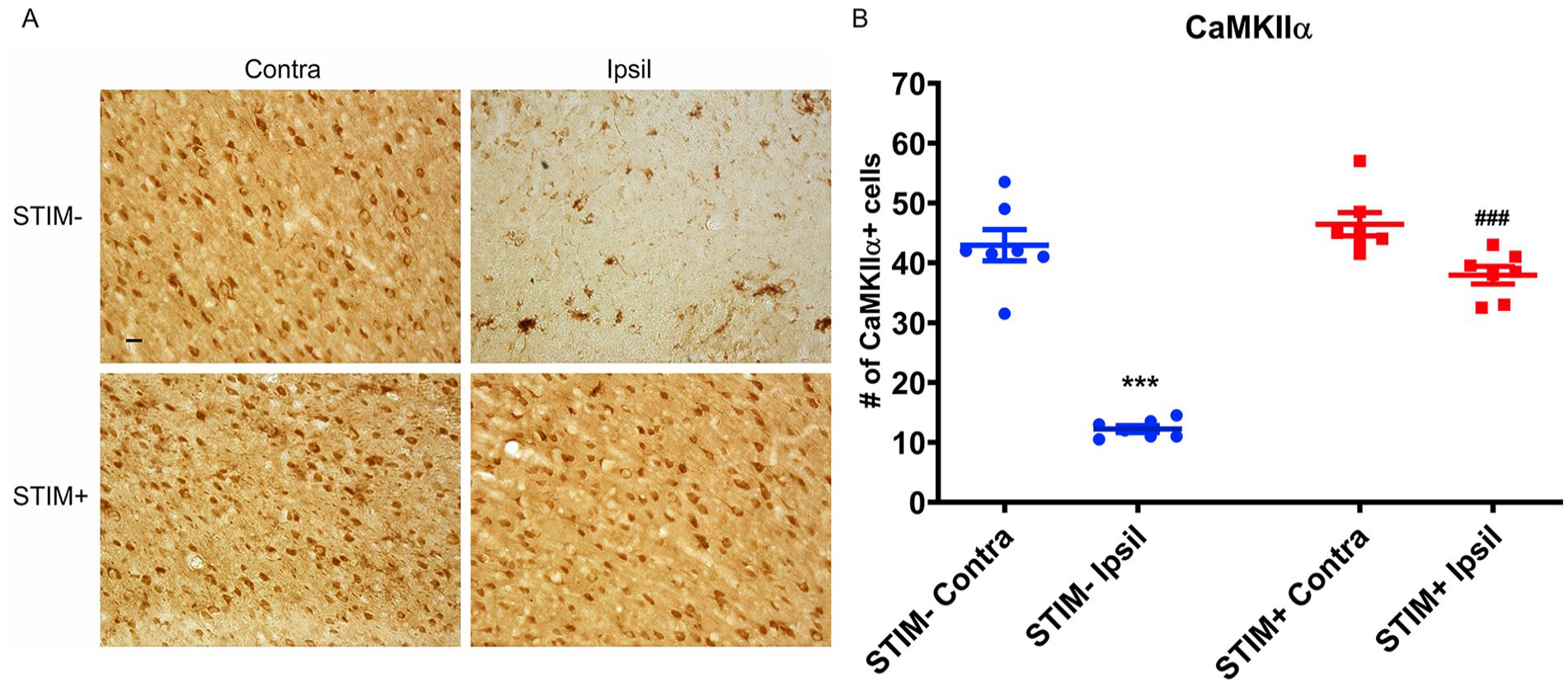
Effect of LCN DBS on CaMKIIα-positive cell density. (A) The density of CaMKIIα-positive cells in the ipsilesional (Ipsil), perilesional cortex of STIM+ significantly greater than that of STIM−. Data was presented as density of cells/µm2±S.E.M., analyzed by one-way ANOVA followed by Bonferroni’ post hoc test. *** P < .0001 when compared to the STIM− Ipsil while ### P < .0001 when compared to the STIM+ Contra. (B) The representative photomicrographs demonstrated the CaMKIIα-positive cells in both ipsilesional (Ipsil) and contralesional (Contra) perilesional motor cortex of STIM− and STIM+ animals. Scale bar is 100 µm.
LCN DBS Does Not Affect the Lesion Volume
The volumes of the CCI lesion induced at the sensorimotor cortex of experimental animals in both STIM− and STIM+ groups were not found to differ significantly (STIM−: 35.57 ± 1.23 mm3 vs STIM+: 36.03 ± 1.10 mm3, P = .7846; Figure S1), suggesting that LCN DBS does not affect lesion volume.
Discussion
Durability of beneficial effects is an essential element of any post-TBI rehabilitation strategy. Our current results provide not only further evidence for a role of stimulating the ascending cerebellar output circuits in enhancing post-TBI motor recovery but, importantly, support proof-of-principle demonstration of durable change. Specifically, 4 weeks of LCN DBS, initiated 4 weeks after CCI induction, improved motor performance across all 3 motor outcome metrics, consistent with our prior findings. 29 The therapeutic benefit was maintained after cessation of treatment, with maintenance of performance across all 3 metrics. The findings are consistent with our hypothesis, based on treatment-enhanced cellular and molecular perilesional reorganization in both chronic post-stroke and post-TBI models.21,22,29,30 The findings are also consistent with carry-over effects observed in our recent first-in-man human translation of DN-DBS for post-stroke rehabilitation. 36 In the present study, similar changes were found to persist at the end of the 2-week carry-over phase, specifically including up-regulation of BDNF and CaMKIIα at the perilesional cortex in the treated group.
The carry-over effect of LCN DBS on motor recovery possibly relies on treatment-induced cellular and molecular modification s across the perilesional area. Chronic LCN DBS-associated motor recovery in the current CCI TBI model was linked to evidence of changes that are associated with calcium-dependent neuroplasticity, a finding that is further consistent with our prior work which revealed evidence of synaptogenesis, in perilesional cortex of LCN DBS-treated, post-stroke animals.21,29 Combined, these data support that treatment-related benefits, once achieved, should be maintained even after stimulation has been discontinued. Notably, although this effect contrasts markedly with most current DBS indications, where continuous stimulation is required to maintain benefit and cessation of stimulation results in motor sign recurrence within seconds to minutes, 37 there is evidence of stimulation-induced acute and long-term cellular changes in certain instances.38,39
The transcranial alternating current stimulation at alpha frequency induced a carry-over effect of EEG power at the occipital cortex of healthy human in the range of the stimulation frequency, which is potentially dependent on the spike-timing dependent reorganization of neural activities.40,41 On the other hand, the DBS-mediated cellular and molecular reorganization plays an important role of disease amelioration or symptom-relieving effect in certain diseases. Single-pulse DBS of the internal segment of the globus pallidus (GPi) in patients with dystonia produces long-term potentiation-like neuronal activities, assessed by repetitive pairs of GPi DBS and transcranial magnetic stimulation onto the primary motor cortex and has been argued to play a mechanistic role in GPi-DBS mediated improvements in that population. 42 Furthermore, DBS to the subthalamic nucleus increased the motor evoked potential of the Parkinson’s disease patients, promoting the long-term potentiation in the motor cortex. 43 In preclinical studies, DBS delivered to the mesencephalic locomotor region in a rat spinal cord injury model promoted the expression of PSD-95 and BDNF in the lesioned spinal tissue associated with the improvement of locomotor functions. 44 The DBS of the nucleus accumbens of rats remotely proliferated the expression of tyrosine hydroxylase and the dendritic length in the prefrontal cortex associated with the anxiolytic effect. 45 Taken together, these findings support the potential for DBS to yield cellular and molecular changes that, similar to the current findings, may play a role in achieving beneficial effects that persist beyond the actual delivery of stimulation.
Cellular and Molecular Mechanisms
Loss of motor control was compensated by the treatment-related synaptogenesis and the sprouting of new fibers. 46 These calcium-dependent cellular modifications are associated with reorganization of preserved perilesional tissue, compensating for loss of neural transmission in the injured area.46-48 We propose that chronic LCN DBS upregulates neural activity across the ascending, glutamatergic DTC pathway, thereby creating conditions that promote calcium/CaMKII-dependent synaptogenesis 21 and BDNF expression. 30 Although BDNF promotes plastic reorganization and rehabilitation in both stroke and TBI,49,50 the causal relationship between CaMKIIα and BDNF under the condition of LCN DBS remains unclear. One possibility is that activation of the CaMKIIα-dependent long-term potentiation signaling pathways increases the expression and release of BDNF which signals other adjacent neurons to undergo synaptogenesis and dendritogenesis51,52 and facilitates also the angiogenesis. 53
LCN DBS Effect on Different TBI Models
The current line of work, including its use of a sensorimotor TBI model and emphasis on motor rehabilitation, arose primarily from our prior experience in applying LCN DBS to treat chronic, post-stroke motor deficits.20-22 As noted previously, however, chronic motor deficits can arise post-TBI and negatively impact overall outcomes and quality of life.3,4 To that end, our initial studies in the sensorimotor, fluid percussion injury TBI model showed promise for improving motor outcomes 29 but did not include an examination of the post-treatment carry-over effects reported here. The current work is further distinguished from that prior effort in its use of the CCI model, which was adopted for its higher consistency as regards the size and location of the injury. 54 Moreover, we had since adopted the CCI approach as it provides similarly high consistency in establishing a midline, prefrontal injury model in order to examine cognition-related behaviors, 30 a model that is more suitable for representing post-TBI cognitive deficits in humans.55,56 Additional work is needed, however, to further characterize the effect of therapy on cognitive behaviors post-TBI as well as to examine the overall durability of any such effects.
Limitation
Interpretation of the current data set is limited by the nature of the study and its design trade-offs. Most importantly, the study does not include several cohorts with multiple treatment durations. Such an experiment would require large groups of animals and could have its own downsides, such as the need for non-concurrent cohorts due to space constraints. Nevertheless, additional experiments can, in the future, identify the limits of stimulation carry-over as well as explore the possibility that longer periods of stimulation may drive even longer carry-over effects. Likewise, shorter treatments can be examined, in order to learn the shortest possible treatment durations that will produce non-inferior results. The implication of experiments examining temporal effects of stimulation are significant for human translation, as they can guide cost-effectiveness of treatment and device design.
It cannot be ruled out that LCN DBS could, as its primary effect, increase home cage, spontaneous motor activity, which, by itself, could promote neuroplasticity and motor recovery. Unfortunately, home cage activity was not monitored throughout the study, however the available evidence from the cylinder test suggests that spontaneous behavior (ie, total number of rears observed) did not differ significantly between the treated and untreated animal across the trial.
Finally, interpretation of our findings is limited to male animals, which were selected due to the higher rate of TBI in the male population combined with preliminary data in our laboratory that showed evidence of stronger post-CCI spontaneous recovery in female animals. 57 Further work is underway to characterize that difference in order to power and design comparable work using female animals as part of a future endeavor.
Conclusion
LCN DBS promotes recovery of motor function following sensorimotor CCI injury and is associated with greater expression of CaMKIIα and BDNF in the surviving cortex. These data have significant implications for the design of future translational efforts, as they suggest that DBS targeting DTC pathways may only need to be delivered temporarily until such time as recovery effects have plateaued.
Supplemental Material
sj-docx-1-nnr-10.1177_15459683241277194 – Supplemental material for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury
Supplemental material, sj-docx-1-nnr-10.1177_15459683241277194 for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury by Hugh H. Chan, Brittany M. Fisher, Margaret A. Oimoen, Latavya Chintada, Hemen Khanna, Claire A. Sonneborn, Olivia Hogue, André G. Machado and Kenneth B. Baker in Neurorehabilitation and Neural Repair
Supplemental Material
sj-jpg-3-nnr-10.1177_15459683241277194 – Supplemental material for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury
Supplemental material, sj-jpg-3-nnr-10.1177_15459683241277194 for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury by Hugh H. Chan, Brittany M. Fisher, Margaret A. Oimoen, Latavya Chintada, Hemen Khanna, Claire A. Sonneborn, Olivia Hogue, André G. Machado and Kenneth B. Baker in Neurorehabilitation and Neural Repair
Supplemental Material
sj-tif-2-nnr-10.1177_15459683241277194 – Supplemental material for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury
Supplemental material, sj-tif-2-nnr-10.1177_15459683241277194 for Carry-Over Effect of Deep Cerebellar Stimulation-Mediated Motor Recovery in a Rodent Model of Traumatic Brain Injury by Hugh H. Chan, Brittany M. Fisher, Margaret A. Oimoen, Latavya Chintada, Hemen Khanna, Claire A. Sonneborn, Olivia Hogue, André G. Machado and Kenneth B. Baker in Neurorehabilitation and Neural Repair
Footnotes
Abbreviations
ANOVA: Analysis of variance; AP: anterioposterial; BDNF: Brain Derived Neurotrophic Factor; CaMKIIα: Calmodulin dependent kinase II alpha; CCI: controlled cortical impact; DTC: dentatothalamocortical; DAB: 3,3’-diaminobenzidine; DBS: deep brain stimulation; DV: dorsoventral; GPi: internal segment of the globus pallidus; IHC: immunohistochemistry; LCN: lateral cerebellar nucleus; PBS: phosphate-buffered saline; PSD-95: Postsynaptic density protein-95; SIU: stimulus isolation unit; TBI: traumatic brain injury.
Author Contributions
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Drs. Machado and Baker have intellectual property and distribution rights in Enspire DBS Therapy, Inc., which is a spin-off of the Cleveland Clinic. Drs. Machado and Baker serve on the Scientific Advisory Board of Enspire DBS Therapy, Inc. Dr. Machado holds a patent titled as “Methods of treating medical conditions by neuromodulation of the cerebellar pathways.” US 7,640,063.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from NIH NINDS [R01NS116384]
Supplementary material for this article is available on the Neurorehabilitation & Neural Repair website along with the online version of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.