
Abstract
Reaching tasks are commonly used in preclinical and clinical studies to assess the acquisition of fine motor skills and recovery of function following stroke. These tasks are often used to assess functional deficits in the absence of quantifying the quality of movement which requires kinematic analysis. To meet this need, this study uses a kinematic analysis in mice performing the Montoya staircase task at 5 and 14 days following a cortical photothrombosis-induced stroke. Following stroke, the mice had reaching impairments associated with sustained deficits including longer, unsmooth, and less individuated paw trajectories. Two weeks after stroke we also detected the emergence of abnormal elbow and shoulder angles, flexion/extensions, and stereotyped kinematic synergies. These data suggest that proximal and distal segments acting in concert is paramount during post-stroke reaching and encourage further analysis of synergies within the translational pipeline of preclinical to clinical studies.
Introduction
Advances in acute stroke treatment have resulted in a significant increase in the number of individuals surviving stroke, yet the trajectory for recovery after stroke has not significantly changed. 1 Stroke remains one of the leading causes of chronic disability, with 80% of patients having motor impairments that often affect the control of the movement of the face, arm, and leg of one side of the body.2,3 These deficits are characterized by the loss of upper motor neuron control over voluntary movements, as well as the emergence of abnormal movement patterns or synergies.4-6
There are multiple definitions for synergy; however, in general, they all refer to the spatiotemporal pattern of coordinated activation of the limbs, muscles, or joints involved in the performance of a movement. 6 These synergy patterns are determined early in life 7 and are able to generate complex motor commands, but are sensitive to modification by training, and cortical injury such as stroke.8-10 The two main synergies of the upper limbs following stroke include flexion synergy, which is characterized by simultaneous shoulder abduction and elbow flexion, as well as extension synergy, which is characterized by simultaneous shoulder adduction and elbow extension. 6 These proximal upper limb synergies reduce the precision and smoothness of the movements and limit the ability to coordinate movement in flexible and adaptable patterns.
In the clinic, the quantification of stereotyped movements and abnormal synergies are often measured using functional ordinal scale measures such as the Fugl-Meyer.11,12 In addition to the use of these functional measurements, the quantification of the quality of movement through fine-grained kinematic analysis has been recommended by the second International Stroke Recovery and Rehabilitation Roundtable taskforce. 13 This recommendation is based on the need to distinguish movements that are responsible for restitution versus compensation, to aid in the development of rehabilitation approaches that target processes underlying motor control and recovery post-injury.13-15
Similar to clinical research, preclinical research has mainly relied on functional outcome measures and there is a growing interest in measuring kinematic movements to enhance translation from the bench to bedside.16,17 Due to the striking similarities between human and rodent reaching movements in grasping tasks, unilateral skilled reaching tasks such as the staircase test and single pellet reaching task are often utilized in both mice and rats.18-23 Both of these tasks are sensitive to detect functional deficits post-stroke as assessed by a reduction in the number of pellets retrieved or reaches performed.18,22,24-31 Kinematic analysis of reaching on these tasks has also illustrated abnormal quantitative distal paw and qualitative proximal movement patterns post-stroke.30,32-36 Additionally, we have previously shown that rats performing the staircase task have abnormal kinematic synergies with inefficient reaching trajectories at 7 days after stroke including elbow flexion, shoulder adduction, and shoulder rotation. 25 How these synergies modify with time after a stroke and whether similar deficits in synergies occur post-stroke in mice remains to be determined. This study, therefore, tested if kinematic distal and proximal movement deficits and/or abnormal kinematic synergies occurred in mice performing the staircase test at 5 and 14 days after a focal stroke.
Materials and Methods
Experimental Design
All procedures were approved by the University of Ottawa’s Animal Care Committee in accordance with guidelines set by the Canadian Council on Animal Care. Fourteen adult male C57BL/6 mice (Charles River, 15–20 g) were single-housed on a 12 hr light/dark cycle (7
Montoya Staircase Skilled Reaching Test
This study used the Montoya staircase test 20 in order to utilize the same kinematic methodologies we established in rats to test our hypothesis that similar abnormal movements and synergies would generalize to mice. 25 Additionally, the staircase test was used, instead of the single pellet reaching task since it allows for analysis of reach-to-grasp movements at 3 separate targets, as opposed to the same location in every reaching event. 23 The mice performed the staircase test within the Behavioral Core Facility at the University of Ottawa. Each day the mice were habituated to the testing room for 30 minutes. Two weeks before surgery, mice were trained (1 30 min trial/day for 14 days) to reach for food pellets (Test Diet® - St Louis, MO-USA). For the first 3 days of training, the top 2 steps (7 and 8) were baited with 1 pellet to encourage eating. After day 3, the pellets were removed from steps 7 and 8 since the mice can use their tongue to reach these steps, while the remaining steps (1-6) were baited with 3 pellets. The 9/14 mice that met the a priori criteria of a minimum of 60% of the pellets eaten during the last 2 days of pre-training were tested before surgery to establish baseline performance. The mice were retested for reaching ability starting 5, 14, and 42 days after surgery. At each of these timepoints there were 4 days of staircase testing with 1 trial/day. Day 1 was used for re-habituation, day 2 and 3 provided an average of their reaching performance, and day 4 was used for kinematic assessment.
Photothrombosis Stroke Surgery and Histology
A stroke was induced in the forelimb motor cortex contralateral to the preferred paw (AP +0.7, ML 2.5) using the photothombosis (PT) method, as previously described. 37 Briefly, mice were anesthetized by inhalation of isoflurane (5% induction, 1.5% maintenance, and 1 L/min oxygen), followed by an intraperitoneal injection of rose Bengal (Sigma, R3877-5G; 10 mg/mL, sterile water) given 5 minutes prior to light application (10 minutes, 3 cm from skull surface, 532 nm wavelength, ∼20 mW power, Beta Instruments). Body temperature was maintained between 36°C and 37.5°C using a rectal probe and feedback blanket (Harvard Apparatus). Mice received 2% transdermal bupivacaine as an analgesic immediately following and 4 hours following surgery.
Animals were anesthetized and perfused, with the brains cryoprotected and sectioned on a microtome (35 μm) as previously described. 37 Cresyl-violet staining was performed on mounted tissue sections and images of the sections were taken with an Aperio Slide Scanner at 20x magnification and exported using the Scanscope software. Blinded measurements for stroke volume were calculated using ImageJ (NIH; 2012, pixels per mm on 20x images).
Kinematic Assessment and Analysis
The mouse staircase apparatus was adapted for kinematic analysis through the use of cameras, calibration frames, and reflective markers similar to our published methodology used in rats.
25
Briefly, as shown in Figure 1A, we reconstructed a four-point kinematic model using markers positioned at the upper shoulder, lower shoulder, elbow, and wrist. Prior to filming, animals were anesthetized using isofluorane (4% induction, 2% maintenance, 1 L/min oxygen) in order to place reflective markers using superglue (Loctite®, gel, Henkel Corp, USA). Ten to fifteen minutes after anesthesia, the mice were placed into the staircase and a 30-minute session was recorded (Sony Handycam®, model HDR-PJ380, 60 fps, USA). Kinematic model and analysis. (A) Montoya staircase apparatus and kinematic model: a four-point 2D kinematic model is used to reconstruct upper shoulder (trunk), lower shoulder (shoulder joint), elbow, and wrist segments. (B) Time-domain features were extracted from linear and angular waveforms; for example, maximum, minimum, negative and positive increments, root mean square (RMS), first and last value. (C) Synergy analysis was conducted considering the frame-by-frame interaction between waveforms. Briefly, we used the first derivative to detect the instantaneous variation of the multi-articular movement, thus, if the limb is advancing and the elbow is flexing the algorithm output was a percent calculated by limb advance—elbow flexion.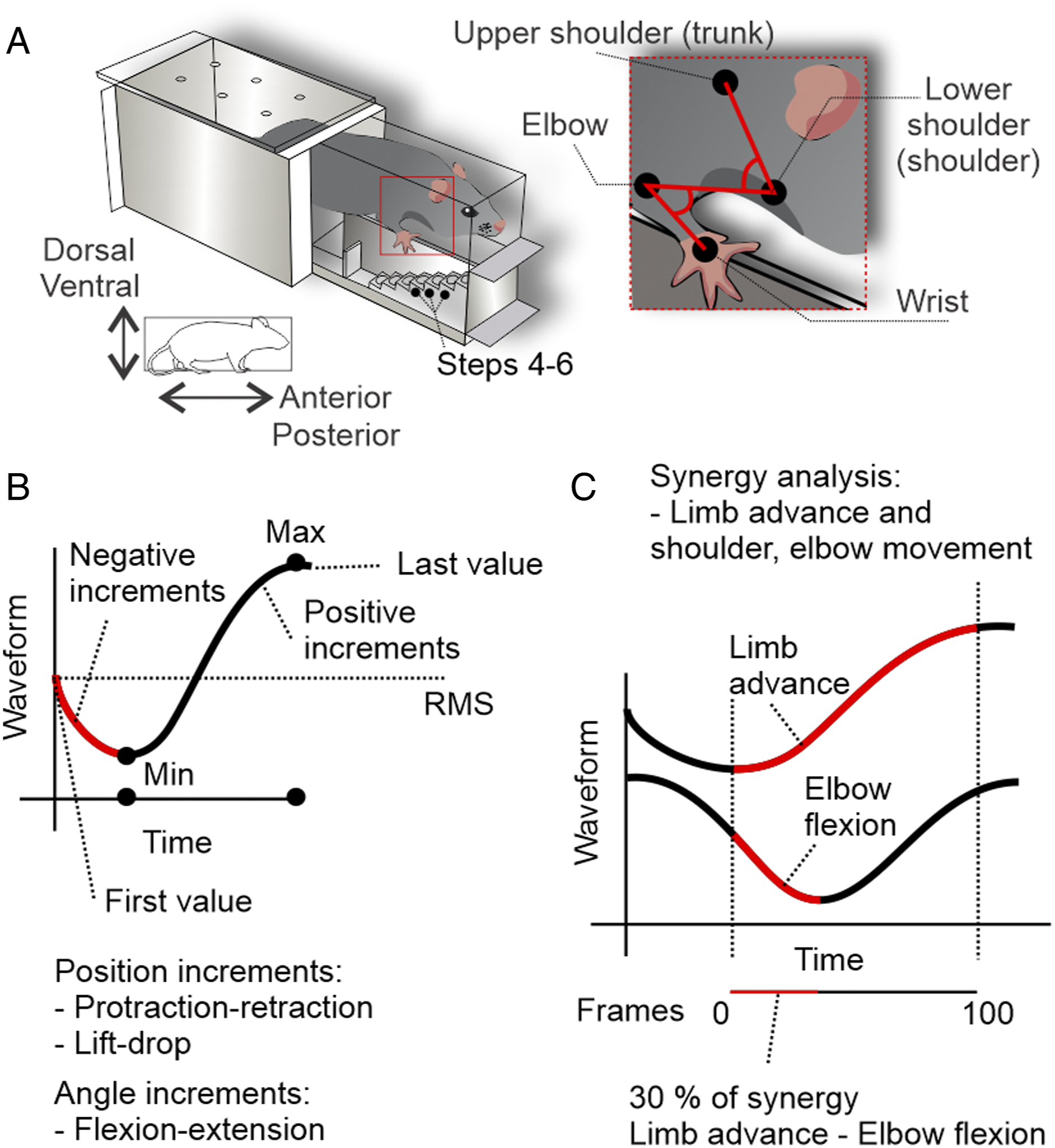
All clearly recorded reaches to steps 4, 5, and 6 were cropped for kinematic analysis using Adobe Premiere Pro® software (Adobe Systems Inc, USA). Kinematic analysis was performed on the 9 mice that had PT surgery. At the pre-stroke, 5 days post-stroke, and 14 days post-stroke time point, a mean of 29.4, 26.9, and 19.2 reaches were analyzed per animal, respectively. At 42 days post-stroke 4/9 mice did not have 5 clearly recorded reaches and therefore for the remaining 5 mice, a mean of 12.2 reaches were analyzed per animal, which were included in the supplementary data.
All video data were digitized using open-source SkillSpector® 1.3.2 software. Raw marker coordinates and joint angles were exported as an ASCII file. LabVIEW® 8.5 custom software routines were developed to analyze kinematic data. Displacement data were normalized by target coordinates (i.e. steps 4, 5 or 6 which all had reflective tape), with all displacement data reported relative to the final endpoint of the reaching.
The position and angle, velocity and acceleration profiles, kinematic synergy, and joint individuation analysis between limb advance, limb lift, and limb drop, and other body segment movement changes, were calculated as described previously. 25 Briefly, a full waveform analysis was conducted in order to extract signal maximum, minimum, time of maximum, time of minimum, the first value (aim), and last value (grasp) (Figure 1B). Euclidean distances were used to calculate path lengths of proximal (trunk, shoulder, and elbow) and distal joints (wrist). Movement segmentation for the shoulder and elbow was measured as the ratio of the distal path length to the proximal path length, thus higher values represent less individuated joint movement. The relation between wrist movements and joint angles was defined as “kinematic synergy” (Figure 1C), whereas the relation between wrist movements, and marker displacements was defined as “joint individuation deficit.”
Statistical Analysis
Data are presented as mean ± SEM. All variables were tested for normality using the Shapiro–Wilk test followed by one-way or two-way ANOVAs and Tukey post-hoc tests. The coefficient of variation analysis was performed to quantify intra-subject variability (SD/Mean). The error and degrees of freedom were adjusted using Greenhouse–Geisser correction for the One-way repeated measure ANOVA. For the principal component analysis (PCA) the data were transformed into z-scores to avoid magnitude artifacts over principal components. To test the ability of our PCA model to separate clusters of animals a Wilcoxon non-parametric statistical test was performed. All statistical analyses were performed using GraphPad Prism 6.01 software and LabVIEW 8.5 software (PCA - Machine learning add-on). Statistical significance was set at P < .05.
Results
Persistent Impairments in the Staircase Test Following Stroke
Daily training of naïve mice on the staircase task resulted in a significant increase in the average number of pellets retrieved over 14 days (Figure 2A; Ftime(4.7, 37.56) = 11.4; P < .001). In total 9/14 (64%) mice met the a priori criteria defined by retrieval of 60% of the pellets and received a PT-induced stroke. The infarcts were targeted to the motor cortex and at the endpoint (45 days post-stroke) had a range in volumes of 0.12 mm3 to 1.44 mm3 with an average infarct volume of 0.69 mm3 ± 0.14 mm3 (mean ± SEM) (Figure 2B). PT stroke induces sustained deficits in staircase performance. (A) The number of pellets retrieved over the 14 days of training on the staircase prior to stroke significantly increased over time. (B) Representative microphotographs of an average lesion size stained using cresyl-violet staining and representative image of the minimum (black), average (gray), and maximum (light gray) stroke volumes at 45 days post-stroke. There was a significant reduction in the (C) number of total pellets retrieved, as well as (D) relative percentage of pellets retrieved post-stroke. This was accompanied by a significant reduction in the (E) number of reach attempts, as well as (F) relative percentage of reaching attempts. Mean ± SEM (n = 9). *P < .05 (pre vs 5 days post-stroke), #P < .05 (pre vs 14 days post-stroke), †P < .05 (5 days vs 14 days post-stroke), **P < .05 (pre vs 42 days post-stroke), and ††P < .05 (5 days vs 42 days post-stroke).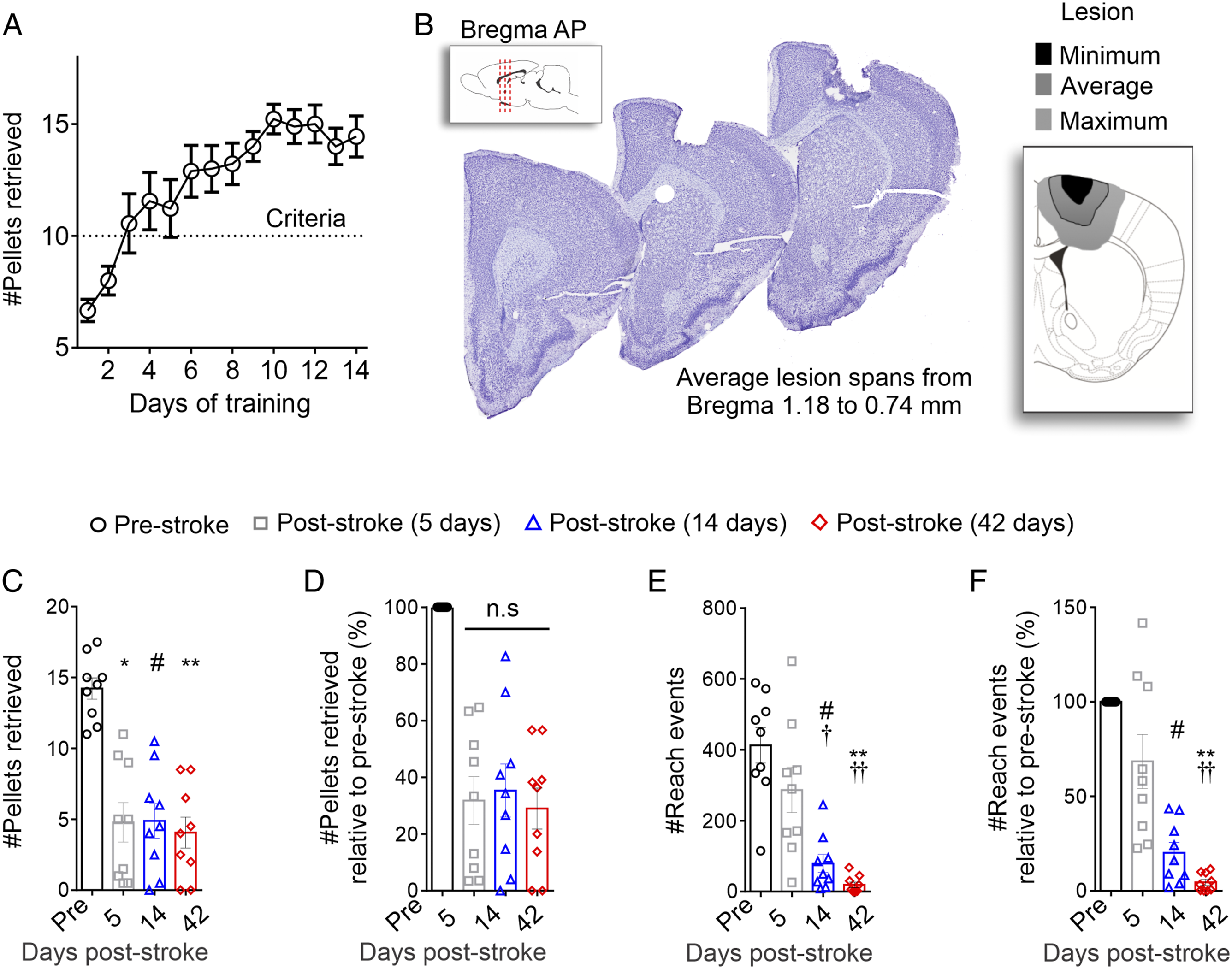
Following stroke, the mice had a significant reduction in the number of pellets retrieved compared to pre-stroke performance (Figure 2c; Ftime (2.6, 20.5) = 25.9, P < .001). Specifically, there were significant deficits at 5, 14, and 42 days after stroke and no significant improvements in performance following stroke. To account for the variability between mice in performance, analysis of the percentage of the pellets retrieved after stroke revealed a significant sustained deficit in performance (Figure 2d; Ftime(2.6, 20.5) = 25.9; P < .001), with an approximate 75% mean reduction in the number of pellets retrieved at 5, 14 and 42 days post-stroke.
The persistent reduction in staircase performance after stroke was accompanied by a significant reduction in the number of reach attempts (Figure 2E; Ftime (2.2, 17.4) = 25.8, P < .001). Unlike the sustained deficits in the number of pellets post-stroke, there was a significant decline in number and relative proportion of reach attempts at day 14 and 42, but not day 5 post-stroke (Figure 2E, Ftime(2.2, 17.4) = 25.8; P < .001; and Figure 2F Ftime(1.2, 9.8) = 31.4; P < .001). Together, these data show that the mice have a significant and sustained deficit in the number of pellets retrieved accompanied by a decline in reach attempts over time following stroke.
Smoothness Deficits Following Stroke
In order to perform kinematic analysis of the reach movement and coordinates of upper limbs during staircase performance, the mice had reflective markers positioned at the wrist, elbow, lower, and upper shoulder, similar to the methodologies we previously published in rats. 25 We measured the dynamic pattern of movement during the voluntary reach that includes a brief aim position, followed by limb lift, advance, drop, and ending in the grasp position at the time of pellet contact. For a visual summary of the kinematic findings as described below, please review Supplementary video 1 for a representative video of an animal pre-stroke and 14 days post-stroke. Analysis of the kinematic movement was done for data collected at baseline, day 5 and 14 post-stroke. Due to the low number of reach attempts and reduced statistical power for the analysis of reach attempts at 42 days post-stroke, a brief preliminary analysis was performed as shown in Supplementary Figure 1.
The paw movement was measured based on the movement of the wrist marker during reaching behavior on the staircase test from the aim position (labeled as 0% of reach) to the final grasp position (labeled as 100% of reach) (Figure 3A). The vertical acceleration profiles illustrated a well-defined pattern pre-stroke, with few vertical velocity adjustments as identified by acceleration zero crossings, as shown by the representative image in Figure 3B. In contrast, there were significantly more frequent velocity adjustments at 5 (Figure 3C) and 14 (Figure 3D) days following stroke (Figure 3E: Ftime(1.6, 12.8) = 11.3, P = .002). This increase was accompanied by significantly more vertical paw trajectory adjustments at 14 days after stroke (Figure 3F: Ftime(1.8, 14.5) = 14.0, P < .001) and a significant increase in the vertical velocity peak of the paw at 5 and 14 days after stroke (Figure 3G, Ftime(1.9, 15.4) = 10.9, P = .001). These findings demonstrate that the mice have less smooth paw movements, characterized by more changes in direction and speed, when reaching in the staircase task after stroke. These findings are also consistent with the reduction in the smoothness of reaching movements reported in mice during the single-pellet reaching task,
32
and our findings in rats performing the staircase task after stroke.
25
Together the results support the theory that across different species and tasks there is a generalized reduction in the smoothness of movements post-stroke. Reduced smoothness following stroke is defined by increased vertical velocity, trajectory adjustments and peak velocity during reaching. (A) Paw trajectory smoothness is quantified by the number of changes in direction (trajectory adjustment) or velocity (velocity adjustments) of the wrist. Acceleration data during the reach from a representative animal at pre-stroke (B; n = 29 trials), 5 days (C; n = 30 trials) and 14 days post-stroke (D; n = 29 trials). At 5 (C) and 14 (D) days the acceleration pattern is more variable with several velocity adjustments when compared to pre-stroke. Post-stroke there is a significant increase in the average number of vertical (E) velocity adjustments, (F) trajectory adjustments, and (G) peak velocity. Mean ± SEM (n = 9); *P < .05 (pre vs 5 days post-stroke), #P < .05 (pre vs 14 days post-stroke), †P < .05 (5 days vs 14 days post-stroke).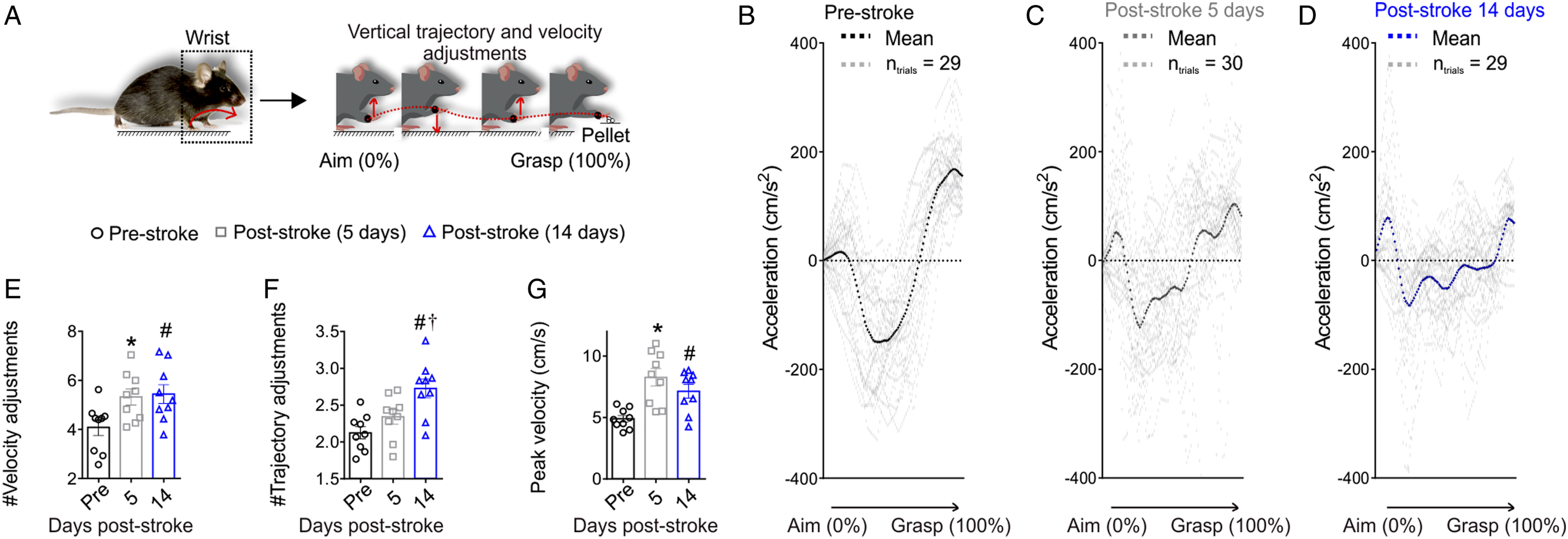
Post-Stroke Trajectories Are Longer and Less Individuated
To test whether the reduction in smoothness was accompanied by stroke-induced changes in the reaching path, shoulder, and wrist path lengths were calculated (Figure 4A). The wrist path lengths were increased at 5 and 14 days post-stroke (Figure 4B; Ftime(2.0, 16.0) = 5.3, P < .001). This was accompanied by a significant increase in shoulder path length after stroke (Figure 4C; Ftime(1.4, 11.0) = 26.7, P < .001). Increased wrist and shoulder path lengths are accompanied by decreased segmentation and increased shoulder protraction during the reach post-stroke. (A) Path length (Euclidian distances) of both the shoulder and wrist were determined during the reach. Both the (B) wrist and (C) shoulder path length is significantly increased following stroke. (D) At 14 days post-stroke there was a significant decrease in the movement segmentation (indicated by an increase in the movement segmentation index), which is the ratio of the wrist to shoulder path lengths. (E) Analysis of shoulder marker displacement through the reach identified an increase in (F) shoulder posterior to anterior movements (shoulder protractions) following stroke (thick lines represent the mean for all animals and thin grey lines represent the standard error). (G) There was a significant increase in average shoulder protraction accompanied by (H) a significant reduction in intra-subject variability post-stroke. Mean ± SEM (n = 9); *P < .05 when compared to Pre, #P < .05 when compared to Pre.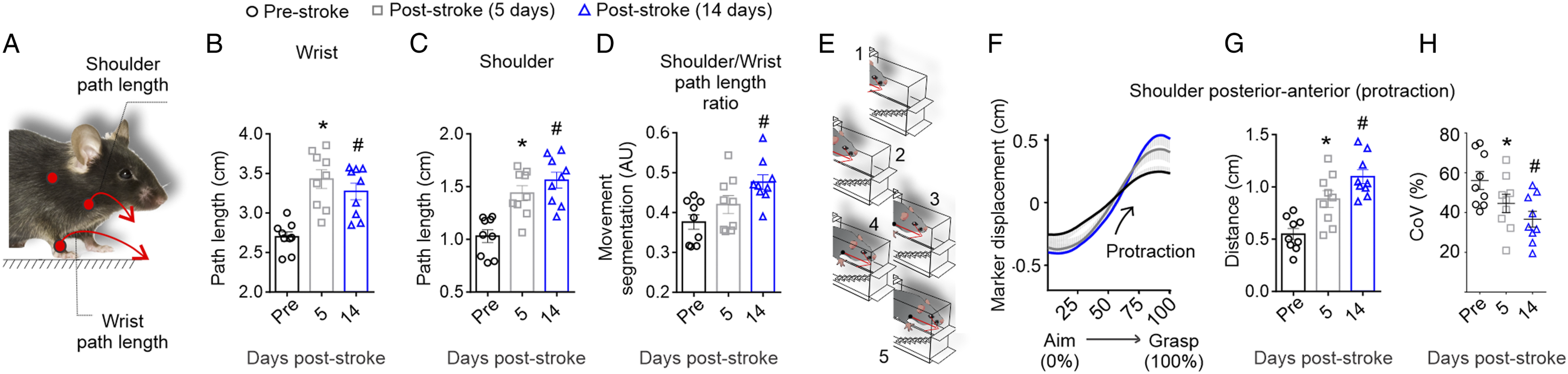
The ratio of measurement between the wrist and shoulder path length was used to determine how much of the movement was segmented. Highly segmented movement refers to the movement of the paw without moving other body parts, such as the elbow or shoulder. This was defined by a movement segmentation score closer to 0, as adapted from Cirstea and Levin (2000).(38) In contrast, lower segmented movement (segmentation score closer to 1) refers to the shoulder/body movement occurring during the advance of the paw. There was significantly less segmentation 14 days after stroke (Figure 4D; Ftime(1.6, 12.8) = 10.1, P = .003). This is further supported by an increase in shoulder protraction (shoulder posterior-anterior movement), as illustrated in Figures 4E and 4F. Quantification of this movement revealed a significant increase in average shoulder protraction distance (Figure 4G; Ftime(1.7, 13.5) = 23.2, P < .001), with an accompanying reduction in the intra-subject variability (Figure 4H; Ftime(1.6, 12.7) = 7.2, P = .01) at 5 and 14 days post-stroke. Overall, these data corroborate that after stroke, mice have a longer paw trajectory when they reach in the staircase, similar to findings shown for single-pellet reach. 32 This data also suggests that the change in paw trajectory measured by the wrist movement, accompanied by an increase in shoulder protraction, could be a compensatory movement.
As shown in Figures 5A and 5B, at the aim position (labeled as 0% of reach) the initial elbow joint angles are more flexed, producing a smaller angle. In contrast, at the grasp position (labeled as 100% of reach) the angles are more extended, producing a larger angle. Analysis of the elbow angles (Figure 5C) revealed that 14 days after stroke, compared to either pre-stroke or 5 days after stroke, there was a significant reduction in elbow angle at aim (Ftime(1.8, 14.8) = 14.7, P < .001) and grasp (Figure 5c; Ftime(1.7, 13.8) = 25.8, P < .001). Similarly, there was a reduction in shoulder angle (Figure 5D) at the aim (Ftime(1.8, 14.3) = 4.4, P = .035) and grasp position (Ftime(1.6, 13.1) = 9.1, P = .005) at 14 days after stroke. Emergence of abnormal elbow and shoulder angles, flexion and extensions, and synergies 14 days post-stroke. (A) Voluntary reaching is preceded by a brief aiming positioning (left panel; 0%) followed by limb lift, advance, and drop (not shown) and ending in pellet grasp positioning (right panel; 100%). B) Elbow joint angle during the reach, lower angles are more flexed, greater angles are more extended. At aiming and grasping the (C) elbow and (D) shoulder are more flexed at 14 days post-stroke. (E) Mean normalized elbow angle from aim to grasp, the negative increments indicate flexion (-Δ) and positive increments extension (+Δ) Similar findings are evident for (F and G) elbow and (H and I) shoulder angular excursions.. (J-O) During limb advance, consistent abnormal flexor synergies as measured by the shoulder and elbow joints are used at 14 days after stroke. (J) Analysis of both the elbow and shoulder during limb advance revealed an increase in (K) shoulder flexion synergies accompanied by (L) a significant reduction in intra-subject variability by 14 days post-stroke. (M) Similarly there was an increase in (N) elbow flexion synergies accompanied by (O) a significant reduction in intra-subject variability by 14 days post-stroke. (P) Analysis of the shoulder ventro-dorsal movement during limb advance revealed an increase in (Q) shoulder lift accompanied by (R) a significant reduction in intra-subject variability by 14 days post-stroke. Mean ± SEM (n = 9); *P < .05 (pre vs 5 days post-stroke), #P < .05 (pre vs 14 days post-stroke), †P < .05 (5 days vs 14 days post-stroke).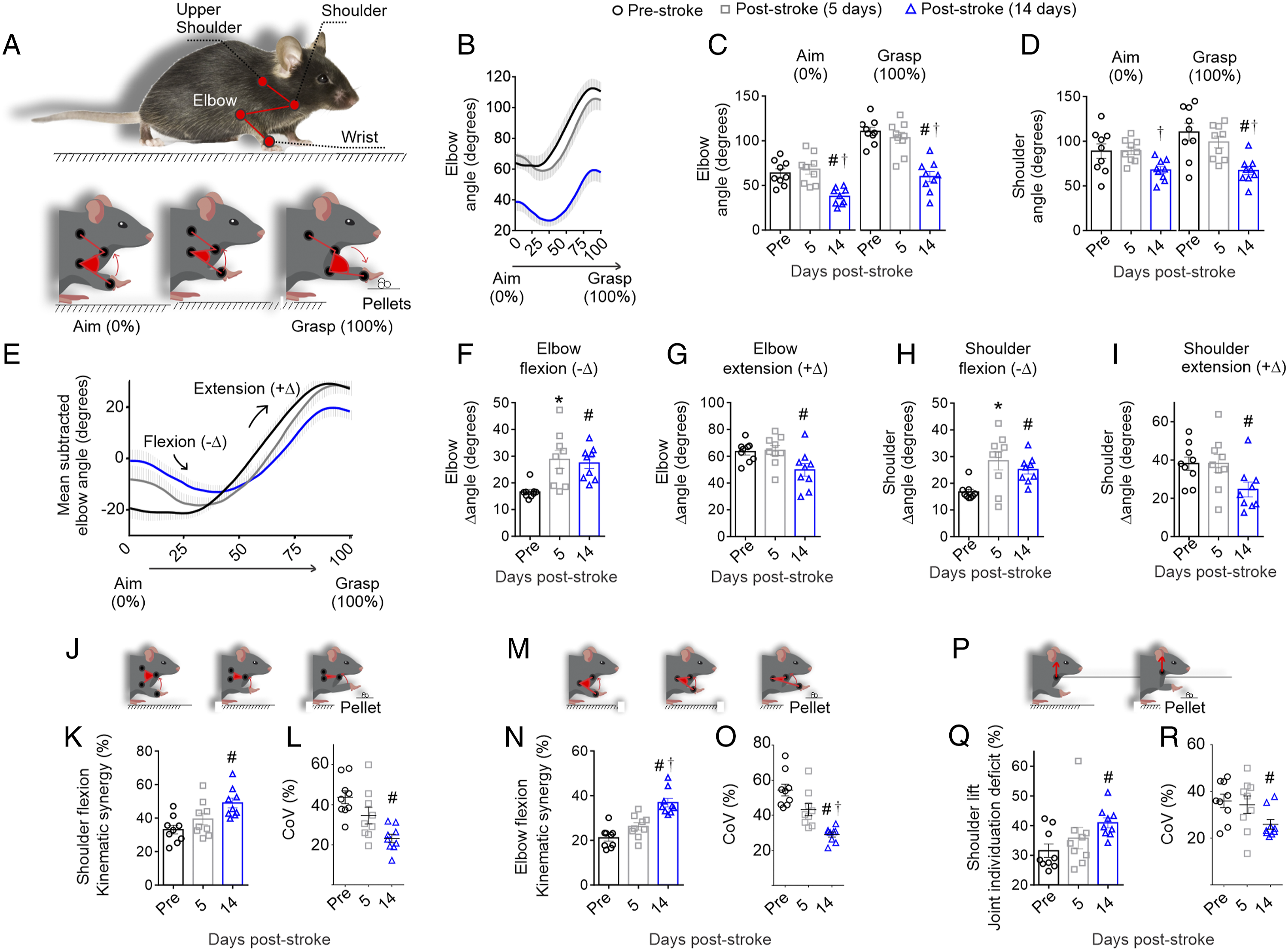
These analyses of the angles of the elbow and shoulder at aim and grasp were conducted using a one-way ANOVA for each measure testing the effect of time. In order to further analyze if there were interactions between time and task phase (i.e., aim or grasp), a 2-way ANOVA was also completed on this dataset. There was a main effect of time and task phase in the shoulder (Ftime(2,16) = 7.0, P = .006; Fphase(1,8) = 28.3, P < .001) and the elbow (Ftime(2,16) = 22.0, P < .001; Fphase(1,8) = 284.1, P < .001). There was also a significant interaction between time and task phase for both joints (shoulder: Ftime*phase(2,16) = 12.2, P < .001; elbow: Ftime*phase(2,16) = 16.9, P < .001), which indicates that differences between aim and grasp angles occurred at 14 days post-stroke.
The reduction in aim and grasp angle in both the elbow and shoulder at 14 days after stroke led us to hypothesize that there was increased flexor tonus after stroke. Flexor tonus is defined by an increase in flexion and reduction in extension of the joints at the static position, and thus the mean normalized elbow angle from aim to grasp was calculated frame-by-frame to better visualize the negative (flexion) and positive (extension) increments (Figure 5E). In agreement with this hypothesis, at both 5 and 14 days post-stroke, the mice display significantly more elbow flexion (Figure 5F; Ftime(1.5, 11.9) = 9.3, P = .006). At 14 days after stroke this was also accompanied by a significant reduction in elbow extension (Figure 5G; Ftime(1.6, 13.0) = 4.8, P = .03). The increase in tonus was mirrored in the shoulder. The mice had a significant increase in shoulder flexion at 5 and 14 days after stroke (Figure 5H, Ftime(1.1, 9.1) = 8.1, P = .017) and specific reduction in extension at 14 days after stroke (Figure 5I, Ftime(1.9, 15.1) = 5.0, P = .020). Overall, these findings indicate that there are differences between aim and grasp angles in the static elbow and shoulder including a delayed onset of increased flexor tonus occurring at 14 days post-stroke.
The increase in flexion at the static position of aim and grasp suggested that abnormal flexion synergies could be occurring during the limb advance from the aim to grasp. Analysis of both the shoulder and elbow angles during limb advance revealed the mice use an abnormal shoulder flexion synergy to advance the limb (Figure 5J and K; Ftime(1.7, 14.0) = 8.0, P = .006) that occurred with reduced variability by 14 days after stroke (Figure 5I; Ftime(1.2, 9.6) = 7.7, P = .017). This was also accompanied by similar abnormal elbow flexion synergy (Figure 5M and N; Ftime(1.8, 14.2) = 16.1, P < .001) that occurred with reduced variability at day 14 post-stroke (Figure 5O; Ftime(1.4, 11.6) = 16.3, P < .001). These abnormal flexion synergies during limb advance are accompanied by an increase in the shoulder lift synergy, which is measured by the combined individuation deficits to advance the limb without lifting the shoulder (Figure 5P and Q; F time(1.8, 14.4) = 6.3, P = .012). Similar to the other increases in synergies, there is less variability at 14 days post-stroke in the shoulder lift synergy (Figure 5R; F time(1.9, 15.0) = 4.4, P = .034). Together, these results suggest that during the volitional action to reach (i.e., extend the limb) there is an overall increase in flexion and ventro-dorsal movements, which are typically used to move upward and not forward. Additionally, these data extend our previous work in rats, 25 by describing a significant reduction in variation and increase in abnormal stereotyped synergies over time post-stroke.
An overfitted principal component analysis (PCA) model was employed to identify if similar patterns were arising from the 128 outcome variables at the different time points (Supplementary Table 1). As shown in Figure 6A, the PCA analysis identified 2 well-defined clusters of the mice at the pre-stroke (black circles within the dark gray ellipse cluster) and 14 days post-stroke (blue triangles within blue ellipse cluster) timepoint. These clusters also had significant differences in PC1 (Wilcoxon, P = .008) and PC2 (Wilcoxon, P = .016) scores (Figure 6B). The variance between the mice was explained by the analysis of the Euclidian distance of the different distribution of loads within both the first and second components (PC1 and PC2, respectively, Supplementary Table 2). As shown in Figure 6C, the variability in PC1 is related to variables relating to angular kinematics (88.2% of load) and abnormal synergies (72.7% of load). PC1 also retained 100% of the loads related to the number of reaching events, lesion volume, and staircase function, whereas PC2 is related to compensation (74% of load), kinetics, and smoothness (76.5% of load), and to a lesser degree staircase function (50% of load). Overall, these findings suggest there is less variability in the dataset at 14 vs 5 days after stroke and align with a reduction in the variation of abnormal synergies that were observed at 14 days after stroke. Principal components analysis (PCA) reduced the dataset dimension and identified 1 group at pre-stroke and day 14 post-stroke. (A) Data of each subject for pre-stroke (black circles), 5 days (gray squares), and 14 days (blue triangles) post-stroke are represented in a new space defined by PC1 and PC2 (%, percent of explained variance). PCA cluster identified a group of mice pre-stroke (visually grouped in gray ellipse) and another cluster at 14 days post-stroke (triangles visually grouped in blue ellipse). Mice at pre-stroke and 14 days post-stroke displayed (B) a significant difference in PC1 and PC2 scores, with (C) many individual parameters being significant within the PC loads of PC1 and PC2. # in the black parameter box in C identifies a significant difference for both PC1 and PC2. *P < .05.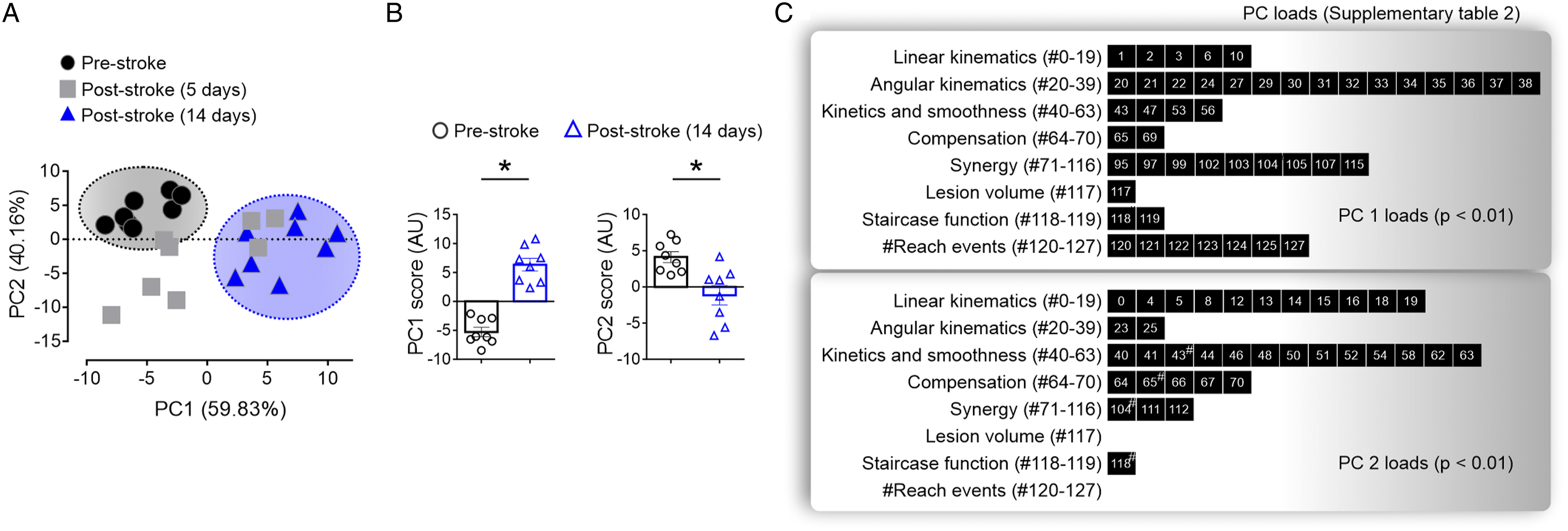
Discussion
This study demonstrates that PT-induced cortical stroke induces dynamic changes in kinematic and synergetic movements. At 5 days after stroke, the kinematic analysis revealed a significant reduction in the smoothness of the reach, as well as an increase in wrist and shoulder path length and shoulder protraction. At 14 days after stroke, these changes were accompanied by limited angular excursions of the shoulder and elbow, and an overall stereotyped increase in shoulder and elbow flexion synergies. This study, therefore identified abnormalities in upper limb synergies in mice that can be used to improve our understanding of processes that may modify recovery.
In mice, the use of kinematic analysis to assess the quality of movement patterns after a stroke has mainly measured distal paw movements, reporting kinematic or kinetic variables of the paw trajectories.32,35,36 Similar to our findings, these studies all report that stroke induces longer and less smooth movements during reaching, as measured during reaching on the single pellet reaching task or head-fixed pulling task on a robotic platform. Our findings are also in agreement with the abnormal wrist trajectories that we reported in the rat endothelin-1 model. 25 The use of these measures is notably aligned with similar metrics of movement time, path length, and velocity that are commonly measured clinically. 14
The deficits in distal movements post-stroke were accompanied by a variety of deficits in proximal joint movements. The mice have an increase in elbow and shoulder flexion during paw aiming at the initiation of movement that was accompanied by an overall reduction in elbow and shoulder extension during the reach-to-grasp movement. These proximal movements are associated with abnormal elbow and shoulder flexor synergies and occur with an increase in shoulder lift synergy, which is suggestive of shoulder and trunk compensation during the limb advance. This data gives quantification to previous qualitative descriptions of shifts in elbow and shoulder position post-stroke in mice, 33 as well as mirror the significant increase in shoulder flexion/lift synergy post-stroke in rats. 25 In the clinic, shoulder flexion/extension kinematic assessments are also often employed, especially for the assessment of 3D pointing tasks. 14 For example, abnormal shoulder synergies after stroke are suggested to occur in an impairment level-dependent manner and are strongly associated with abnormal task performance, as well as lesion size and location.6,39,40 Given the scarcity of literature quantifying proximal movements after stroke in rodents, our data provides a foundation that shows the occurrence of these abnormal proximal movements in mice. Future work should determine longitudinally if these measures vary across preclinical models that vary in size and location of stroke.
Examining both distal and proximal measures concurrently at two time points after stroke provided additional insights. First, this analysis revealed that the relationship between proximal and distal impairments can be ambiguous. For example, although the paw trajectory had an increase in path length post-stroke, the data suggest that increasing flexion synergy and compensation reduces wrist path length. These findings highlight how distal and proximal measures need to both be dissected to determine how they act in concert following stroke. Secondly, proximal deficits were shown to occur after distal deficits in stereotypical patterns. This finding contrasts the lack of a proximal to distal gradient in motor deficits appearing clinically, with upper extremity range of motion analysis showing that the loss of hand function is related to loss of ability to move many segments of the upper extremity.12,41 Thus, future studies will be required to explore the temporal relationship of proximal vs distal deficits to determine if our findings in mice generalize to different species and other types of preclinical stroke models. Lastly, the deficits in synergy at 14 days after stroke were stereotyped with a striking reduction in the intra-mouse variability, as shown by the significant reduction in the coefficient of variation. Although not included in our main results, our preliminary analysis of the few reaches that were performed at 42 days post-stroke, further suggests that these changes are additionally sustained long-term (Supplementary Figure 1). Moreover, the PCA analysis of all the variables analyzed also supports less inter-mouse variability in the outcome measures at day 14 post-stroke, compared to the mice at 5 days after stroke. Although this analysis is preliminary and limited due to small sample size, it demonstrates the utility and the feasibility of analysis of time-series reaching kinematics datasets, and is in alignment with the consensus-based recommendations for stroke recovery trials. 13
Here we show a comprehensive kinematic analysis of a reach-to-grasp task in mice that encompassed most of the variables used in the clinic such as path length, trajectory smoothness, articular angles, and kinematic synergies, that are required to better align clinical and preclinical stroke studies. 16 These findings raise many more questions. Are the changes in kinematic associated outcomes after stroke forms of compensation or recovery? Additionally, how do these changes relate to overall performance? In rats we have shown a lack of correlations between functional deficits in pellet retrieval and changes in kinematic measures at 7 days post-stroke, suggesting that functional performance on the Montoya staircase (reduction in pellets retrieved and reach attempts) does not fully delineate stroke-induced impairments and cannot distinguish between compensation and recovery. 25 This study included analysis at 2 time points post-stroke with the hope of unraveling the nuances between compensation and recovery. Following stroke, the mice had a sustained ∼75% mean reduction in the number of pellets retrieved post-stroke accompanied by a significant decline in the average number of reach attempts over time post-stroke. It is tempting to speculate that, since the mice in this study are using fewer reaching attempts to retrieve the same reduced number of pellets post-stroke, there are performance improvements occurring that are associated with the kinematic changes that could be forms of compensation or recovery. Indeed, compensatory behavior is still poorly understood in rodents and may constitute an alternative route for optimizing functional outcomes after stroke.42,43 Future studies will be therefore required to test the hypothesis of whether the alternations in kinematic measures observed in the mice are forms of compensation or recovery.
This study identifies abnormal kinematic movements and synergies in mice that appear after stroke. Moving forward, the future of this work to develop into larger preclinical studies is bright given recent advances in literature. In naïve mice, significant gains are being made in understanding the fundamental mechanisms for how motor sequences are generated,44-46 which will benefit preclinical work in stroke recovery by providing a road map to identify how stroke modified these processes. Secondly, behavioral kinematic analysis has made it feasible to complete larger sample sizes required through discovery, validation, and automation of 3D markerless tracking systems such as DeepLabCut, 47 use of synthetic animated mice to generate training data for behavioral analysis, 48 as well as creation of home-cage group housed reaching tasks. 49 Thus, preclinical studies will be able to test larger sample sizes, a variety of stroke models, and different reaching tasks, with the goal to produce targeted therapies to improve reaching impairments in stroke survivors.
Supplemental Material
sj-mp4-1-nnr-10.1177_15459683211058174 – Supplemental Material for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke
Supplemental Material, sj-mp4-1-nnr-10.1177_15459683211058174 for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke by Gustavo Balbinot, Sebastien Denize and Diane C. Lagace in Neurorehabilitation and Neural Repair
Supplemental Material
sj-mov-2-nnr-10.1177_15459683211058174 – Supplemental Material for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke
Supplemental Material, sj-mov-2-nnr-10.1177_15459683211058174 for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke by Gustavo Balbinot, Sebastien Denize and Diane C. Lagace in Neurorehabilitation and Neural Repair
Supplemental Material
sj-pdf-3-nnr-10.1177_15459683211058174 – Supplemental Material for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke
Supplemental Material, sj-pdf-3-nnr-10.1177_15459683211058174 for The Emergence of Stereotyped Kinematic Synergies when Mice Reach to Grasp Following Stroke by Gustavo Balbinot, Sebastien Denize and Diane C. Lagace in Neurorehabilitation and Neural Repair
Footnotes
Acknowledgments
We thank Dr. Greg Silasi for assistance in establishing data acquisition, Anthony Carter Biomedical Research Manager with the Canadian Partnership for Stroke Recovery (CPSR) for performing the stroke surgeries, and Dr. Dale Corbett for critical discussions and review of the manuscript. This work was supported by a grant from Canadian Institute of Health Research (DL), and the Canadian Government - Emerging Leaders in the Americas Program (ELAP; CIC #509/2016; GB) and by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES/Brazil, GB).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.