
Abstract
Defective muscle coordination for balance recovery may contribute to stroke survivors’ propensity for falling. Thus, we investigated deficits in muscle coordination for postural control and their association to body sway following balance perturbations in people with stroke. Specifically, we compared the automatic postural responses of 8 leg and trunk muscles recorded bilaterally in unimpaired individuals and those with mild to moderate impairments after unilateral supratentorial lesions (>6 months). These responses were elicited by unexpected floor translations in 12 directions. We extracted motor modules (ie, muscle synergies) for each leg using nonnegative matrix factorization. We also determined the magnitude of perturbation-induced body sway using a single-link inverted pendulum model. Whereas the number of motor modules for balance was not affected by stroke, those formed by muscles with long latency responses were replaced by atypically structured paretic motor modules (atypical muscle groupings), which hints at direct cerebral involvement in long-latency feedback responses. Other paretic motor modules had intact structure but were poorly recruited, which is indicative of indirect cerebral control of balance. Importantly, these paretic deficits were strongly associated with postural instability in the preferred activation direction of the impaired motor modules. Finally, these deficiencies were heterogeneously distributed across stroke survivors with lesions in distinct locations, suggesting that different cerebral substrates may contribute to balance control. In conclusion, muscle coordination deficits in the paretic limb of stroke survivors result in direction-specific postural instability, which highlights the importance of targeted interventions to address patient-specific balance impairments.
Introduction
Proper balance control is essential for mobility and activities of daily living.1-5 When balance is perturbed, fast “automatic” postural muscle responses are the first protective mechanism to restore balance. These postural responses are highly coordinated and tuned to the direction of the balance perturbation.6-8 Studies in decerebrate cats have demonstrated that the brainstem harbors the circuitry for these responses in these animals.9,10 Yet the pronounced balance deficits in individuals with hemisphere lesions11,12 suggest that cerebral structures are also involved in human postural control. This idea is further supported by electroencephalogram studies revealing broad cortical activity in response to balance perturbations. 13 However, the contribution of cerebral hemispheres to balance control is not fully understood.13-15
The role of cerebral structures in human postural responses can be studied using stroke as a disease model. People with stroke demonstrate smaller and delayed postural responses to balance perturbations compared with unimpaired individuals.11,16-19 Delays in postural responses are, however, not the same for all muscles, indicating that coordination of muscle activity for maintaining balance is disrupted after stroke.11,18 Moreover, the deficits in muscle responses are different across stroke survivors because of the known heterogeneity of the disease. 18 Consequently, it is difficult to capture the distinct muscle coordination deficits that can exist after stroke with conventional electromyography (EMG) analysis techniques that assess activity of individual muscles independently. Hence, multivariate analytical tools are needed to identify stroke-related deficiencies in muscle coordination for balance control across different muscles.
We propose to characterize stroke-related muscle coordination deficits of postural responses with factorization algorithms identifying muscle synergies or motor modules.20-22 In these analytical techniques, a muscle synergy is defined as a group of muscles activated together, but these are not necessarily the pathological poststroke synergies described before. 23 Previous studies that have applied this technique to poststroke gait were able to identify stroke-related deficits in muscle coordination and their detrimental effects on functional performance.24,25 It is unknown, however, how stroke affects motor modules for postural control.
In the present study, we aimed to identify stroke-related deficits in motor modules used for balance and their functional consequences. We hypothesized that people with stroke would demonstrate deficits in the structure of motor modules and their temporal recruitment based on the impaired amplitude and timing of postural responses reported in individual muscles. 26 We also hypothesized that each deficit would result in a distinct pattern of direction-specific postural instability given that individual muscle synergies for balance have been shown to counteract specific perturbation directions.7,27,28 Finally, we hypothesized that stroke-related deficits would be heterogeneously distributed across stroke survivors because of the heterogeneity of the disease. We believe that these deficits could become important targets for balance rehabilitation interventions poststroke.
Materials and Methods
Participants
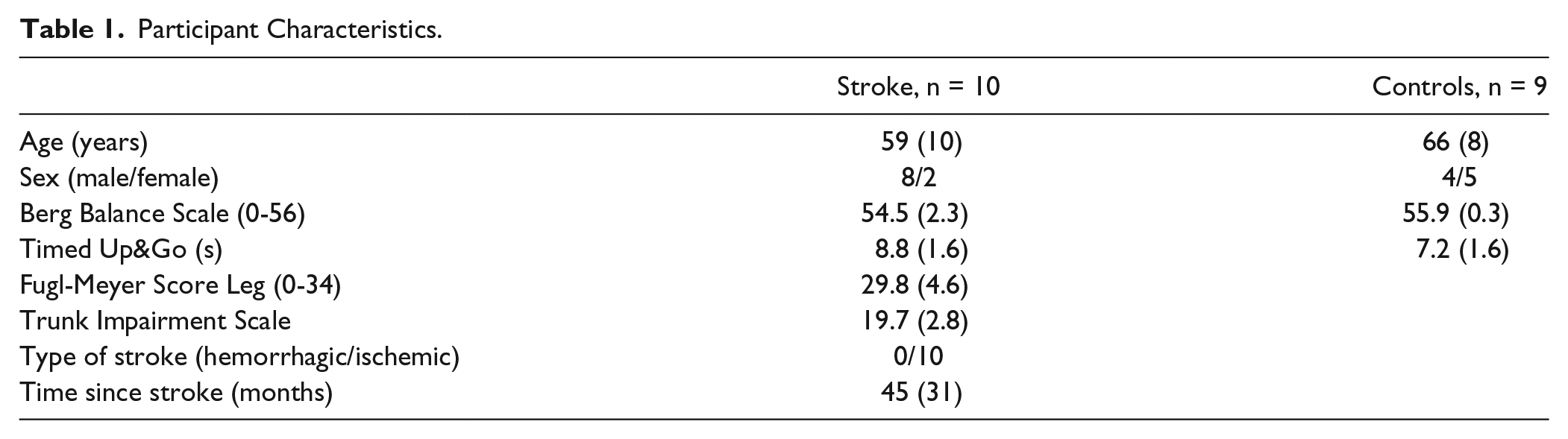
A total of 10 people with mild to moderate motor impairments caused by unilateral supratentorial lesions (>6 months) and 9 healthy controls participated (Table 1). Participants had to be able to stand and walk independently or under supervision (Functional Ambulation Categories ≥ 3). Individuals with neurological (except stroke), cognitive (Mini Mental State Examination < 24), or musculoskeletal impairments were excluded. Written informed consent was obtained from all participants. The study was approved by the Medical Ethical Board of the region Arnhem-Nijmegen (The Netherlands) and conducted in accordance with the Declaration of Helsinki.
Participant Characteristics.
Experimental Setup and Protocol
Participants stood with their feet 4.5 cm apart on a movable platform that unexpectedly translated in each of 12 evenly distributed directions (Supplementary Figure 1). Perturbation directions were defined with respect to each leg such that 0°, 90°, 180°, and 270° represented lateral, forward, medial, and backward translations, respectively. The platform translated with an acceleration of either 0.5 m/s2 (low intensity) or 1.5 m/s2 (high intensity) for 300 ms followed by a constant velocity phase of 500 ms and deceleration phase of 300 ms (Supplementary Figure 1). Low-intensity perturbations in all directions were applied first to induce feet-in-place responses. Subsequently, high-intensity perturbations were applied to investigate stepping responses. There were at least 2 to 4 trials collected per direction for each intensity. This article focuses on the analysis of the feet-in-place responses. EMG activity for the high-intensity perturbations was only used for normalization purposes as indicated below. The total number of collected feet-in-place trials was 49 ± 10 for controls and 44 ± 8 for people with stroke. For safety reasons, all participants wore a harness, and stroke survivors wore an ankle brace (ASO, Medical Specialities, Wadesboro, NC) on the paretic side, which provided only minimal support.
Activity of the following muscles was recorded bilaterally at 2000 Hz: erector spinae (ERSP), gluteus medius (GLUT), biceps femoris (BFEM), semitendinosis (SEMT), rectus femoris (RFEM), peroneus longus (PER), tibialis anterior (TA), and soleus (SOL). Additionally, 3D kinematics were recorded at 100 Hz with a motion analysis system (Vicon Motion Systems, UK). The start of the perturbation was determined with a digital trigger signal.
All participants underwent clinical assessment prior to the balance task. These included Fugl-Meyer leg motor score, 23 Trunk Impairment Scale, 29 and Berg Balance assessment. 30
Data Analysis
EMG Analysis
We systematically controlled the quality of the EMG signals to minimize the effect of movement artifacts on our results (see Supplementary Information 1 for details). EMG data that passed the quality control were band-pass filtered at 35 to 450 Hz, de-meaned, rectified, and low-pass filtered at 40 Hz. 7 Muscle activity during the postural response (PR) was characterized as the average activity over 3 consecutive 75-ms time bins (PR1, PR2, and PR3).7,31,32 Because stroke survivors typically have delayed PRs,11,18 we shifted the start of the first time bin (PR1) for all muscles within a leg to the first onset of muscle activity observed across all perturbation directions and muscles of this leg.33,34 As such, the relative timing of activity across muscles within a leg was preserved after this temporal shift. This procedure was done separately for each leg and each perturbation intensity.
Muscle activity during the PR (PR1, PR2, and PR3) in each muscle was normalized taking into account all directions and perturbation intensities. Specifically, each muscle’s activity was divided by the root mean square of all data bins for standing responses of that muscle. We then shifted the normalized data by subtracting its lowest value to avoid overrepresentation of muscles that showed little modulation across directions. Subsequently, we generated a matrix with normalized mean responses per muscle (8 muscles × 36 time points = 12 directions × 3 PRs). Note that recordings from the stance leg of high-intensity perturbations were included in the normalization procedure to consider the maximum responses that could be elicited in a nonstepping limb.
Normalized data of each individual leg were decomposed as the product of 2 matrices: a matrix

To test the robustness of the dimensionality results (ie, between-group comparison of number of motor modules), we repeated this procedure with principal component analysis (PCA) instead of NMF. Both factorization algorithms yielded similar conclusions when comparing number of motor modules between legs (ie, control vs paretic vs nonparetic legs; P = .55 for NMF and P = .24 for PCA using the Kruskal-Wallis test). We used Pearson correlations between
Kinematic Data Analysis
The postural instability induced by the perturbation was characterized with the maximum angular displacement of the body relative to the preperturbation body position. The body was modeled as a single link-inverted pendulum, where the height of the pendulum was the seventh cervical vertebra (C7) and the base was the projection of C7 onto the floor at the start of the perturbation. We computed the body’s maximum angular displacement during the first second after perturbation onset per trial and averaged these values across trials of the same perturbation direction.
Statistical Analysis
We tested for differences in motor module recruitment between stroke survivors and controls using a general linear model for repeated measures for each motor module and PR separately. The
If activation of a motor module was different between groups, we used Pearson correlations to determine the association between activation coefficients of this module (
If motor modules were missing between groups, we compared body sway values and clinical scores between participants with and without this motor module using a Mann-Whitney test. We only considered body sway values for the preferred direction of the motor module. Note that muscle responses and body sway were averaged across legs in the control group prior to performing these association analyses (regression and Mann-Whitney tests). P values <.05 were considered statistically significant.
Results
Stroke Survivors Exhibited Normal Complexity of PRs but Atypical Paretic Motor Modules
We found that stroke survivors with mild to moderate motor impairments did not have fewer modules than controls; however, some of their paretic motor modules were atypical. Specifically, stroke and control groups both exhibited 3 to 5 motor modules accounting for >77% and >78% of the variability in PRs for every muscle and perturbation direction, respectively (Supplementary Table 1). This similarity shows that the complexity in postural responses was unaffected by stroke. In contrast, some paretic motor modules differed from those of controls. More specifically, we consistently observed 3 motor modules (W1-W3; Supplementary Figure 2) in controls, but only W1 (formed by SOL and GLUT) and W2 (formed by TA, PER, and RFEM) were identified in all paretic legs (Figure 1). The presence of W2 in stroke individuals indicates that TA exhibited PRs even if not all stroke survivors could voluntarily activate this muscle during the Fugl-Meyer assessment. On the other hand, W3 primarily formed by hamstring muscles was atypical in 4 out of 10 paretic legs. This motor module was replaced by either a subject-specific motor module (WSS) or a motor module formed by ERSP (W4). Unlike the paretic side, motor modules W1 to W3 were intact in 9 out of 10 nonparetic legs. In sum, the complexity of PRs was not affected by stroke, but the structure (ie, muscles activated simultaneously) of paretic motor modules was abnormal because of low hamstring postural responses.
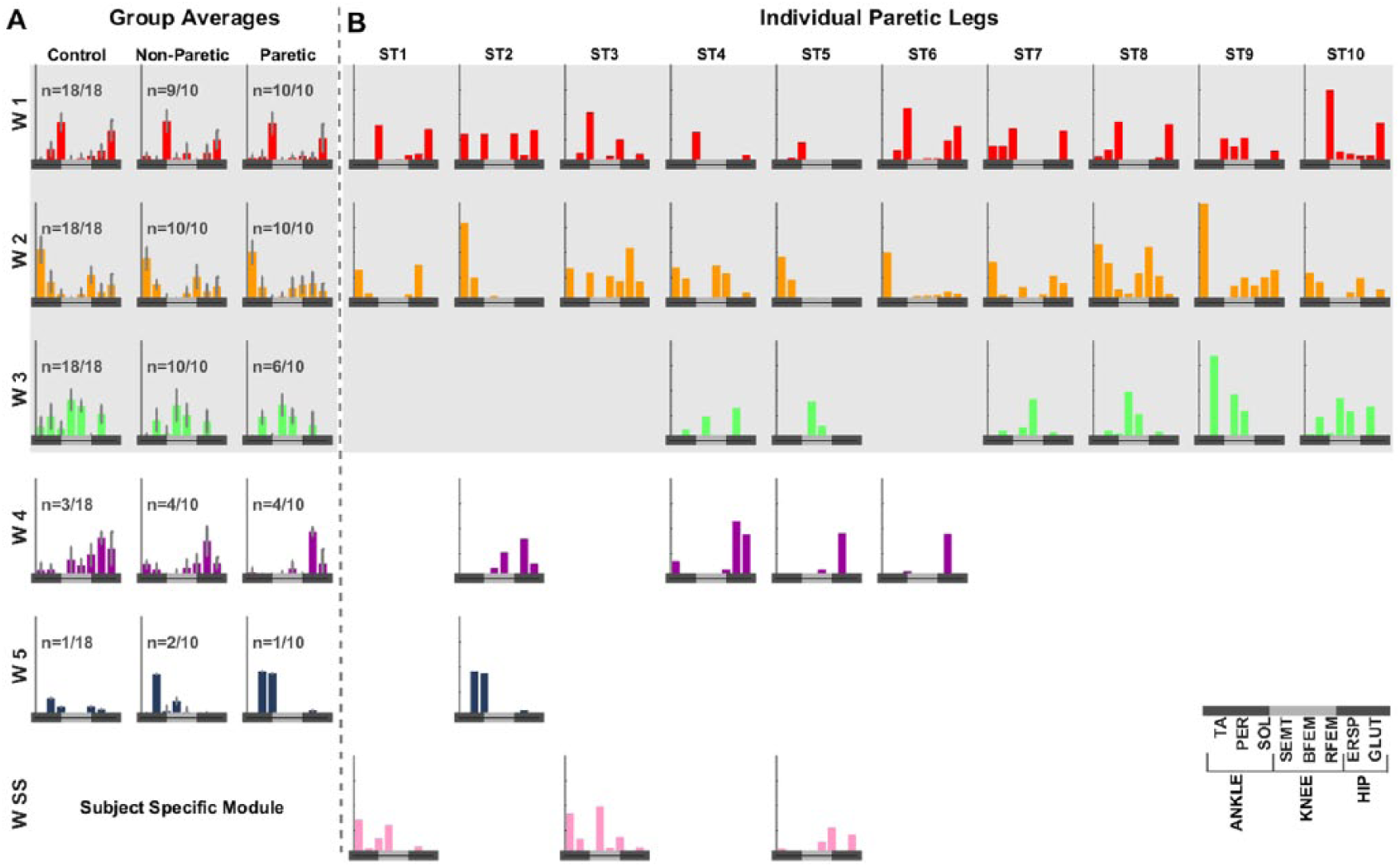
Motor modules in controls and people with stroke: A. Group averages of motor modules identified in controls, nonparetic legs, and paretic legs are shown. Every motor module is formed by a subset of muscles that are consistently activated simultaneously. The structure of each motor module is indicated by the relative activity across muscles within a module, which is represented by the height of the bars. Error bars represent interquartile ranges for each subject group. Modules W1 to W3 were consistently present in controls and nonparetic legs. W1 was mainly formed by soleus and gluteus medius (SOL and GLUT), W2 was composed of anterior leg muscles (tibialis anterior[TA], peroneus longus [PER], rectus femoris[RFEM]) and to a lesser extent GLUT, and W3 was formed by hamstrings (biceps femoris [BFEM], and semitendinosis [SEMT]) and erector spinae (ERSP). In addition to W1 to W3, we identified motor modules W4 composed of GLUT and ERSP and W5 composed of PER and SOL. Although these 2 motor modules were not consistently found in our cohort of participants (ie, W4 and W5 were only identified in only 3 and 1 out of 18 legs, respectively), both modules have previously been identified when participants experienced large balance perturbations.7,32 B. We also show individual paretic motor modules to appreciate differences across participants. On the paretic side, W3 was missing in 4 out of 10 people with stroke. The missing motor module W3 was replaced by either subject-specific modules (WSS) or motor module W4 primarily formed by ERSP.
Deficiencies in Recruitment of Motor Modules for Balance After Stroke
Although motor modules in both groups were activated for the same perturbation directions, the magnitude of activation was different, particularly for perturbations loading the paretic leg (Figure 2; colored [stroke] vs gray lines [controls]). Note that the paretic leg was loaded when the platform moved medially with respect to the paretic side (120°-240° in the paretic leg’s reference frame, Figure 2A) and laterally with respect to the nonparetic side (0°-60° and 300°-330° directions in the nonparetic leg’s reference frame, Figure 2B). Differences between groups were most pronounced in the activation of paretic and nonparetic W2 when stroke individuals were falling backward and toward the paretic side. In these directions, we specifically observed reduced activity of paretic W2 in PR1 [LEG effect: F(2, 35) = 3.48, P = .042; LEG × DIR effect: F(22, 385) = 1.78, P = .019; post hoc: P < .05 for 90°-180°] and to a lesser extent in PR2 [LEG × DIR effect: F(22, 385) = 1.93, P = .01; post hoc: P < .05 for 150°-210°] and PR3 [LEG × DIR effect: F(22, 385) = 1.99, P = .01; P = .031 for 210°]. In PR3, there was also a slight increase of paretic W2’s activity for directions 0° (P = .049) and 60° (P = .01) that were unrelated to its preferred recruitment direction. On the other hand, activity of nonparetic W2 was larger than in controls when stroke individuals fell backward and loaded their paretic side, particularly for the later PRs (P < .05 for PR2 60° and PR3 0° and 60°-120°). Differences in activity of W3 between groups were mostly observed in the nonparetic leg, but also on perturbations that loaded the paretic side [F(22, 341) = 2.71, P < .01 for LEG × DIR in PR2]. Specifically, nonparetic W3 had increased responses to backward platform translations (300°-330°) and reduced responses to forward ones (30°-90°). However, these latter ones were inconsistently observed in controls, suggesting that this may not represent an actual deficit in stroke survivors. Finally, activity of paretic and nonparetic W1 increased compared with controls in response to lateral translations (P < .05) in PR3 only. Taken together, stroke-related changes in activation of motor modules were most pronounced in paretic and nonparetic muscles when the body was displaced toward the paretic side.
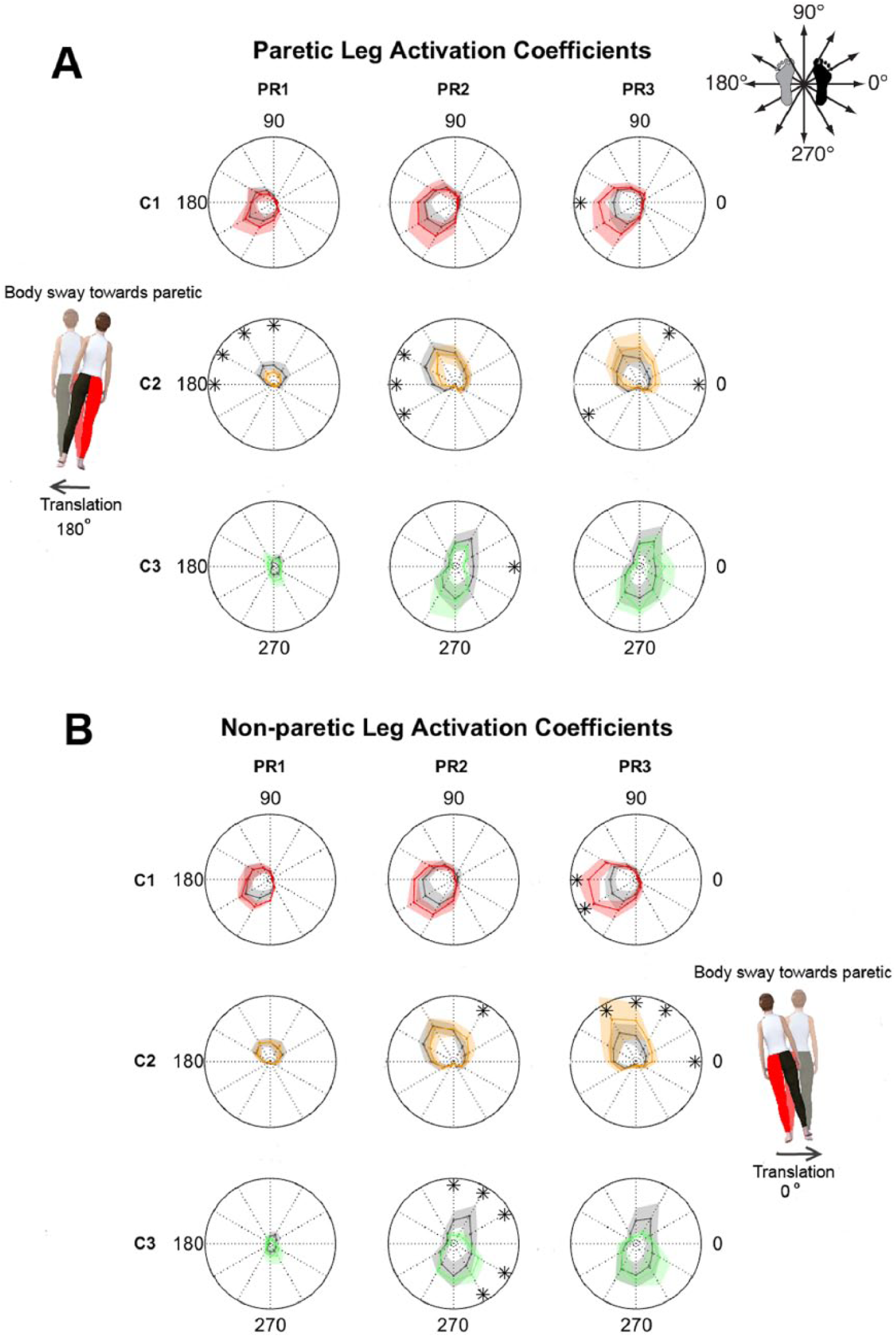
Activation coefficients for control, paretic, and nonparetic motor modules: A. Activation coefficients C1 to C3 for paretic and control motor modules W1 to W3. B. Activation coefficients C1 to C3 in nonparetic legs compared with controls. Activation coefficients C1 to C3 are displayed for the different postural responses (PRs) and translation directions. Polar plots indicate the mean activation magnitude (C) as a function of perturbation direction for stroke individuals (colored lines) and controls (gray lines). Shaded areas represent the interquartile range. *P < .05 for post hoc comparison between stroke patients and controls. Directions of the platform movements with respect to participants’ reference leg (black foot) are shown in the top-right schematic and the avatars. The red leg in the avatars corresponds to the paretic leg. Stroke-related changes in activity of motor modules were most pronounced in C2 and C3, which indicate the recruitment of W2 and W3, respectively. Note that the low initial activity C2 of paretic W2 when the body is displaced backward and toward the paretic side is followed by large activity of this motor module in both legs.
Stroke-Related Deficits in Paretic Motor Modules Were Associated With Body Sway Following Balance Perturbations
The atypical recruitment and structure of paretic motor modules, which were the 2 prominent deficits poststroke, were associated with increased body sway in specific directions (Figure 3). For example, individuals with reduced early activation of W2 (yellow line) exhibited larger body sway when the platform moved in directions 90° to 180°, which typically elicited large PRs in this motor module and displaced the body posterolaterally. A strong relation between body sway and recruitment of W2 was further evidenced by the univariate regression analysis including all participants (Figure 3B). This regression shows that individuals who recruited W2 less (ie, small C2 activation values) experienced larger body sway on balance perturbations in the preferred activation directions of this module (R2 = 0.68; P < .001). Similarly, individuals who replaced the paretic W3 with another motor module demonstrated larger forward body sway compared with stroke patients with intact W3 (P = .02) and controls (P = .05) following platform displacements in the preferred activation directions of W3 (240°-300°) displacing the body forward. Taken together, our findings suggest that deficits in paretic muscle coordination are disruptive because they lead to larger body sway on balance perturbations, typically eliciting PRs in the defective modules.
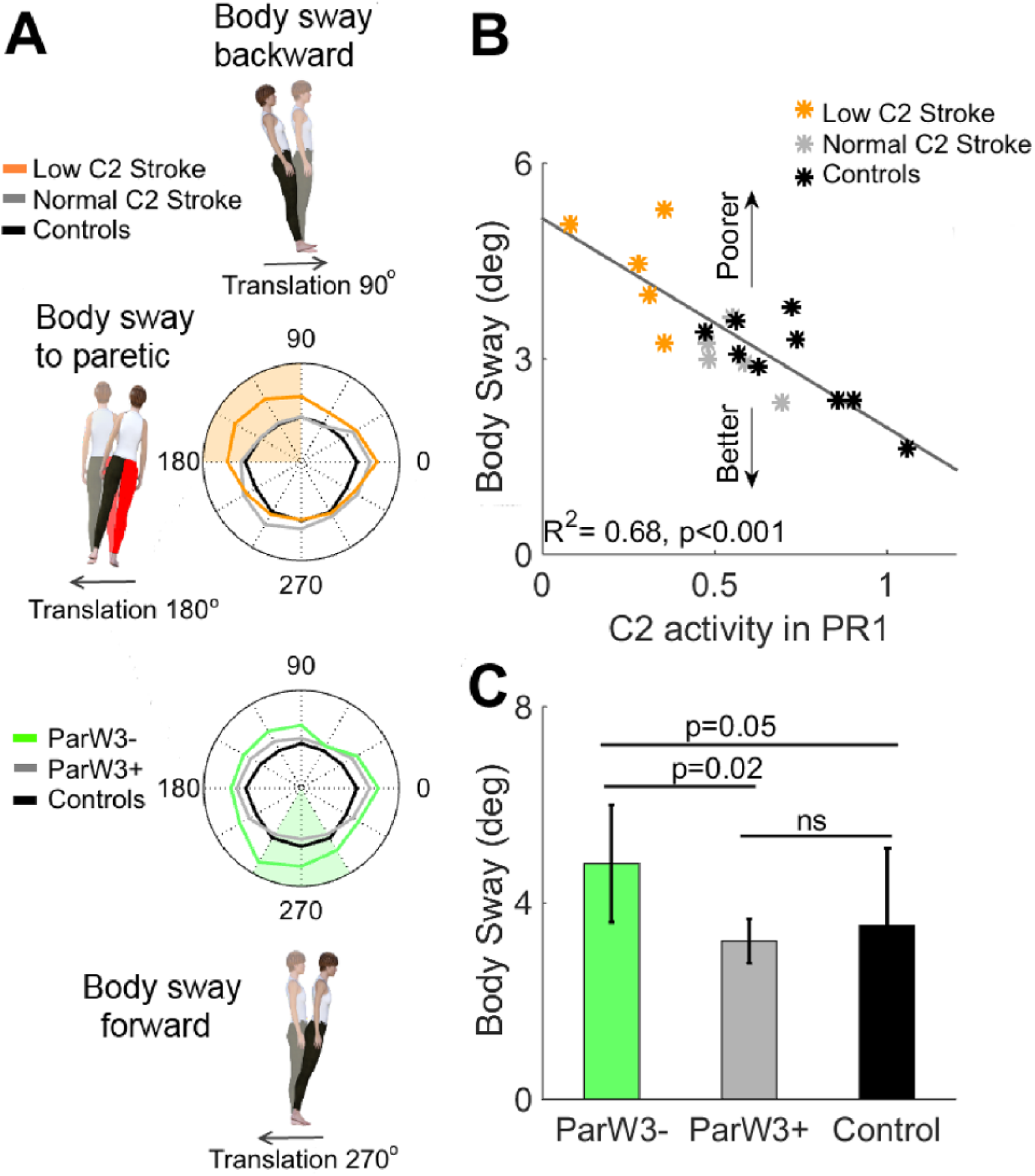
Paretic leg deficits in motor modules related to body sway: A. Maximum body sway as a function of perturbation direction for controls (black line), stroke patients with either low activation coefficient C2 (yellow line) in PR1 or missing W3 (green line), and stroke patients without the respective deficit (gray line). The avatar’s red leg corresponds to the paretic leg. The colored shaded areas indicate the preferred activation directions of the impaired motor module W2 (yellow patch) or W3 (green patch). Maximum body sway values were averaged for these perturbation directions for the statistical analyses shown in panels B and C. B. Regression characterizing the relation between mean activation coefficient C2 in PR1 and mean body sway across preferred activation directions of motor module W2. These 2 variables exhibited a strong negative association, indicating that those with lower initial activation of W2 also had larger body sway. C. Mean body sway for preferred activation directions of W3 across participants without W3 (ParW3−), stroke survivors with intact W3 (ParW3+), and controls. Stroke survivors without W3 demonstrated significantly larger body sway compared with stroke patients with intact W3 (P = .02) or controls (P = .05). In sum, stroke-related deficits in paretic motor modules were strongly associated with increased body sway following perturbations in the preferred activation directions of the defective modules.
Stroke-Related Paretic Deficits in Motor Modules Were Poorly Associated With Clinical Outcome Measures
Although deficits in paretic PRs were related to the extent of body sway following balance perturbations, we did not find clear associations between paretic muscle coordination and clinical scores (Figure 4A). Remarkably, neither of the paretic deficits in muscle responses to recover balance was related to individuals’ performance on the Berg Balance Scale, which suggests that this test is not sensitive enough to aberrant muscle activity presumably underlying poor balance control poststroke. The deficits in paretic motor modules were also not associated with performance on conventional tests assessing motor function of the leg (Fugl-Meyer leg motor score vs activity of W2 in PR1: P = .27; R = 0.39). The same results were observed if the activation of W2 was regressed with Fugl-Meyer subscales specifically assessing motor function of ankle dorsiflexion in sitting (P = .28; R = −0.38) or standing (P = .30; R = −0.36). On the other hand, the trunk motor score (Trunk Impairment Scale) was related to PR deficits in stroke survivors. Specifically, we found that stroke patients who replaced W3 with other motor modules had significantly poorer Trunk Impairment Scale scores than those with intact W3 (17.0 ± 2.2 vs 22.2 ± 1.2, P < .01). We also observed that individuals who scored lower on the Trunk Impairment Scale exhibited larger forward body sway on preferred perturbation directions of W3 (240°-300°; R2 = 0.63, P < .001; results not shown). In sum, the performance during clinical tests often used in stroke survivors did not reflect aberrant muscle activity to maintain balance, except for the Trunk Impairment Scale. Thus, this is the only clinical measure tested in this study, which is indicative of poor muscle control for balance.
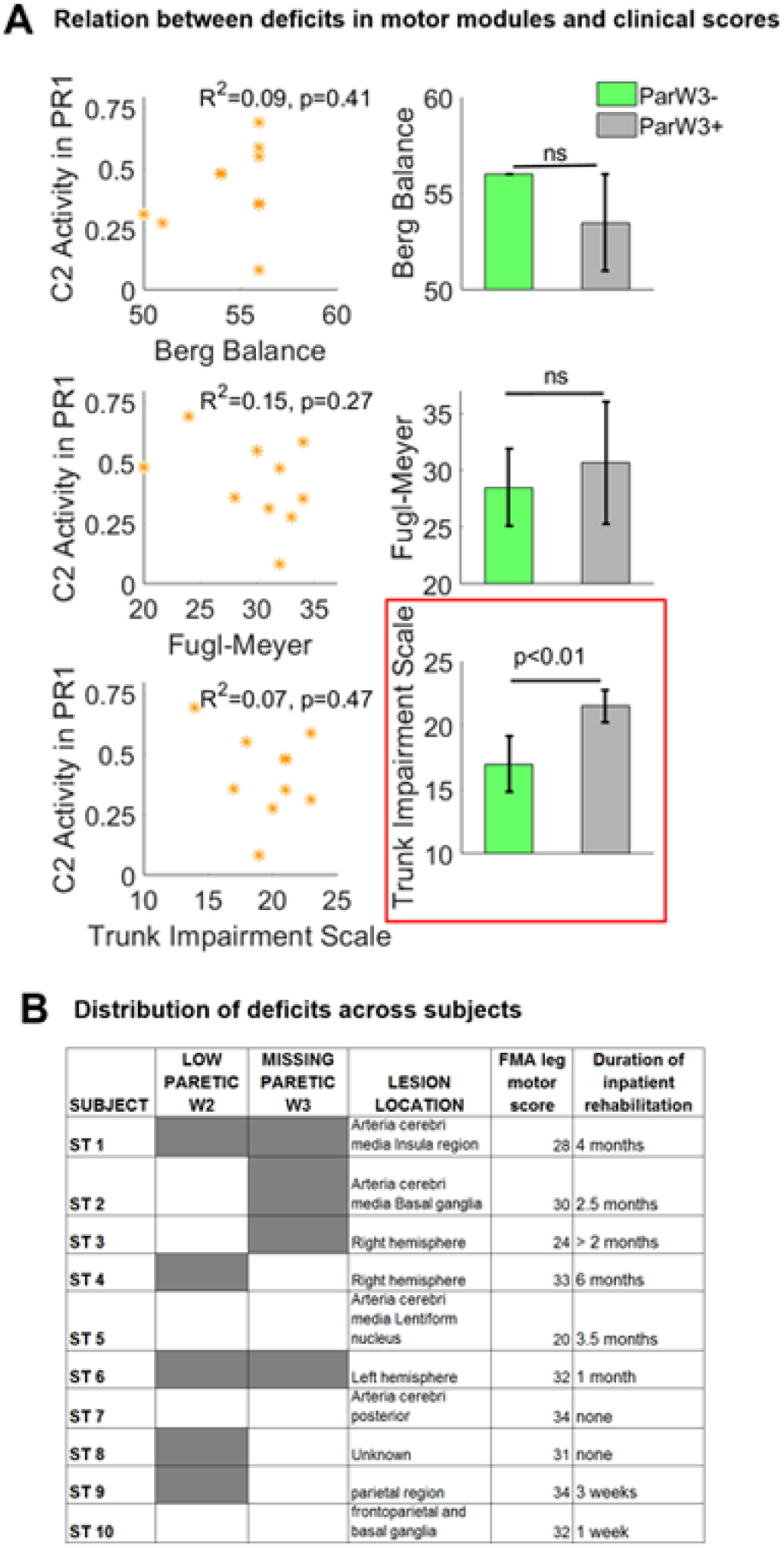
Relation between paretic leg deficits in motor modules and clinical assessment: A. Relation between muscle coordination deficits in the paretic side and clinical scores. The mean initial low activation of W2 (low activation coefficient C2 in PR1) across preferred activation directions of this motor module (y-axis) was not significantly correlated to either Berg Balance Score (top left), Fugl-Meyer assessment (FMA) leg motor score (middle left), or Trunk Impairment Scale (bottom left). Similarly, mean Berg Balance Scores (top right) and mean Fugl-Meyer leg motor scores (middle right) across stroke survivors with (ParW3+) and without W3 (ParW3−) did not significantly differ. On the other hand, stroke survivors with missing W3 had significantly lower Trunk Impairment scores (lower right). This indicates that lack of activity in paretic hamstrings forming W3 was associated with poorer trunk control. B. Muscle coordination deficits in the paretic side and lesion locations are shown for individual participants. Presence of either low activity in paretic W2 or missing W3 is illustrated in gray. The other columns include clinical characteristics of each stroke survivor. Specifically, we present lesion locations as determined by computed tomography scans. Note that more detailed information about lesion location was not available because magnetic resonance imaging scans are not a standard clinical assessment following a stroke in the Netherlands, where the data were collected. Nevertheless, these results demonstrated that muscle coordination deficits and lesion locations were heterogeneously distributed across stroke participants. In addition, subject-specific Fugl-Meyer leg motor score and self-reported duration of inpatient rehabilitation are presented in the right 2 columns.
Heterogeneous Distribution of Balance Deficits and Brain Lesions Across Stroke Survivors
Stroke survivors included in this study differed with regard to the type of muscle response deficiency and the location of their brain lesion (Figure 4B). Note that the 2 deficits that we identified in stroke survivors (ie, atypical recruitment and structure of paretic motor modules) were not consistently paired. Therefore, deficits in muscle coordination for postural control poststroke were heterogeneously distributed across our clinical cohort with mild to moderate impairments. In addition, the locations of brain lesions in the participants with stroke were also heterogeneous. These results suggest that distinct brain areas differently contribute to muscle coordination for human balance control, but further studies are needed to explicitly test this hypothesis.
Discussion
Our results show that automatic postural responses poststroke can be reproduced by a similar number of motor modules as in controls. However, the paretic motor modules were either abnormally activated or replaced by atypical motor modules. Importantly, these deficiencies compromised balance in stroke survivors because they were strongly associated with increased body sway following perturbations in the preferred activation directions of the defective modules. Finally, these deficits were heterogeneously distributed within the group of stroke survivors suffering from cerebral lesions in distinct locations, suggesting that different neural structures might regulate motor modules for balance control.
Neural Mechanisms for Balance Control Based on Paretic Postural Responses
The similar number of motor modules in stroke survivors with mild to moderate motor impairments and controls indicates that the complexity of muscle coordination for balance control is comparable across these populations. This finding suggests that an intact cerebrum is not necessary for multidirectional tuning of postural responses which is consistent with previous reports of intact direction-specific postural responses in decerebrate cats. 9 In contrast, this tuning is severely compromised after spinal cord transection. 36 Taken together, these results suggest that sophisticated tuning of postural responses is housed in the brainstem, rather than in the spinal cord or cerebrum. Our dimensionality results are in contrast to previous observations of fewer motor modules in stroke survivors than in unimpaired individuals during reaching and walking.25,37,38 These discrepancies may result from differences in muscle coordination requirements across behaviors (ie, arm dexterity vs balance), differences in recorded muscles or impairment level of participants across studies, or our normalization procedure, which contrary to prior work,25,37 reduced the magnitude of paretic tonic activity strongly influencing data dimensionality. 39 Finally, it is worth pointing out that we identified fewer motor modules than in previous balance studies—likely because of the smaller number of recorded muscles and lower intensity of balance perturbations that we used. Nevertheless, the muscles forming motor modules in our study match those from prior reports.7,31 In summary, we found that stroke survivors with mild to moderate motor deficits and controls have a similar number of motor modules for balance, suggesting that intact cerebral structures are not needed for multidirectional tuning of postural responses.
Although the complexity of postural responses is not affected in stroke individuals with mild to moderate impairments, paretic motor modules are atypically structured or abnormally recruited, supporting the idea of direct or indirect cerebral involvement in human balance control. Direct cerebral involvement 40 is supported by the atypical structure of the paretic motor module formed by hamstrings (ie, W3), which exhibited long latency responses. This finding is consistent with the proposed direct cerebral pathway for long-latency feedback responses in the arm41,42 and leg 13 that may be enabled by direct projections from the cortex to individual muscles.43-45 On the other hand, indirect cerebral involvement in postural responses is supported by the abnormal recruitment of motor modules formed by anterior muscles (ie, W2), suggesting that cerebral structures may regulate the gain of motor modules that are harbored in the brainstem. 14 Interestingly, not all motor modules had deficits poststroke. Particularly, the responses of calf muscles when falling forward were intact after stroke, indicating differences in the extent of cortical involvement in postural responses across muscles. Other studies have also shown more supraspinal control when falling backward than forward,11,12,46 which are directions with distinct postural threat. Thus, larger risks of falling may require more cortical involvement to maintain balance. 47 Our results raise the question of what cerebral areas may contribute to direct and indirect cortical control of balance. We speculate that the supplementary motor area and the prefrontal cortex are likely candidates because the recovery of activity in these regions on postural perturbations has been associated with better balance control in stroke survivors.48-50 Although further studies are needed to pinpoint the specific cerebral regions contributing to human postural control, we believe that distinct anatomical structures mediate the direct and indirect cerebral control of balance because stroke survivors with heterogeneous cerebral lesions exhibited different muscle coordination deficits. In summary, the abnormal structure and recruitment of paretic motor modules suggest direct and indirect involvement of cerebral structures in human balance, and distinct neural regions might underlie these processes.
Compensatory Nature of Nonparetic Balance Responses
Although nonparetic motor modules were formed by the same muscles as the ones identified in unimpaired individuals,7,32 the recruitment of most of the nonparetic motor modules was abnormally large. There are 2 possible reasons explaining this higher recruitment. First, it could be a compensation strategy for low paretic postural responses because increased nonparetic responses were only observed for later time bins that followed the initially low paretic activity. This compensation is consistent with the suggested cortical shaping of later phases of postural muscle activity based on contextual demands. 14 Second, an increased magnitude in postural reflexes resulting from asymmetric weight bearing commonly observed after stroke could also explain the larger nonparetic responses. 12 Taken together, our findings suggest that motor modules for balance control are intact on the nonparetic side, but they are activated more than in controls to possibly compensate for paretic postural deficits.
Deficits in Paretic Postural Responses Compromise Balance Poststroke
A key finding of our study was that we elucidated the functional consequences of aberrant patterns of muscle activity poststroke. Specifically, low activation of the motor module formed by anterior muscles (W2) was strongly associated with increased body sway toward the paretic side in stroke survivors, which may explain their propensity to fall in this direction. 51 Also, the relation between atypical motor module W3 and limited trunk control suggests that the poor activation of proximal muscles in stroke survivors may contribute to their reduced capacity for sustaining large balance perturbations. 26 Thus, the observed associations between defective paretic motor modules and direction-specific instability confirms the distinct biomechanical functions of motor modules as it has been observed in gait, 24 upper-extremity movements, 52 and balance control in standing and dynamic conditions. 53 Particularly, motor modules in balance are associated with the control of the center-of-mass displacement, 31 which is consistent with our results. In sum, our findings support the idea that motor modules have a functional biomechanical output, and as such, defective motor modules negatively affect behavior.
Study Limitations
There are a few limitations inherent to behavioral studies in stroke survivors that could affect the interpretation of our results. First, the limited number of trials that we could collect in stroke survivors precluded us from doing trial-to-trial analysis in the covariation of muscle activity. Thus, although our results characterize well individuals’ impairments in the balance task, the motor modules that we identified do not necessarily represent muscles receiving a common neural drive. Another limitation of our study is that we accounted for the delayed onset of postural responses in stroke survivors 26 by shifting the start of the time period that we used to characterize automatic postural responses. However, we kept the same duration of this time window for unimpaired and poststroke groups without considering that the activity of some muscles poststroke could be delayed beyond the fixed time period that we used. Thus, the lack of paretic responses in the hamstrings, and therefore atypical motor modules including these muscles, might have resulted from delayed, rather than missing, hamstring activity. In addition, our small sample size did not allow us to identify plausible associations between atypical recruitment or structure of paretic motor modules and subsequent compensatory muscle activity on either leg. Future studies with larger number of participants are needed to explore this possibility. Finally, most of our participants were mildly impaired (ie, 29 < Fugl-Meyer Assessment leg motor score ≤ 34), which limits the generalizability of our results to more severely affected individuals. 54 Thus, further studies are needed to identify balance coordination deficits in more severely affected stroke survivors.
Clinical Implications
Our findings have 3 noteworthy clinical implications for people with mild to moderate impairments poststroke. First, we identified deficient muscle coordination that was mostly pronounced when falling backward and toward the paretic side. These deficits may underlie stroke survivors’ propensity to fall toward the paretic side. 51 However, it is yet to be determined if similar muscle coordination impairments are observed during more dynamic tasks such as walking or reactive stepping. Second, deficient paretic responses were not associated with the Berg Balance Score (BBS). This could be explained either by the fact that the BBS mostly evaluates aspects of balance control not characterized in this study, such as anticipatory corrections to self-initiated movements, or because of a ceiling effect of this metric in individuals with mild to moderate impairments. Yet the fact that we observed muscle coordination deficits in these participants with near-maximum BBS supports that this test is not sensitive to poor reactive balance control, 55 which is associated with greater fall risk in people with stroke. 56 Thus, we believe that tests evaluating this key aspect of balance, such as the MiniBESTest, 57 may be preferred for assessing balance performance poststroke in clinical settings. Third, we identified distinct balance deficits across individuals, highlighting the need for personalized rehabilitation of balance in stroke survivors. In conclusion, our findings provide insights that may contribute to reducing fall risk in individuals mildly to moderately affected by stroke, for which effective interventions are currently lacking. 58
Supplemental Material
Supplementary_Figure1_MethodsFinal – Supplemental material for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control
Supplemental material, Supplementary_Figure1_MethodsFinal for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control by Digna de Kam, Alexander C. Geurts, Vivian Weerdesteyn, and Gelsy Torres-Oviedo in Neurorehabilitation and Neural Repair
Supplemental Material
Supplementary_Figure_2_ControlSyn – Supplemental material for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control
Supplemental material, Supplementary_Figure_2_ControlSyn for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control by Digna de Kam, Alexander C. Geurts, Vivian Weerdesteyn, and Gelsy Torres-Oviedo in Neurorehabilitation and Neural Repair
Supplemental Material
Supplementary_MaterialR1CleanVersion – Supplemental material for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control
Supplemental material, Supplementary_MaterialR1CleanVersion for Direction-Specific Instability Poststroke Is Associated With Deficient Motor Modules for Balance Control by Digna de Kam, Alexander C. Geurts, Vivian Weerdesteyn, and Gelsy Torres-Oviedo in Neurorehabilitation and Neural Repair
Footnotes
Supplementary material for this article is available on the Neurorehabilitation & Neural Repair website along with the online version of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was financially supported by a Netherlands Organization for Scientific Research Veni Research Grant (916.10.106) to Vivian Weerdesteyn (![]() ) and the National Science Foundation (NSF 1342183) to Gelsy Torres-Oviedo. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
) and the National Science Foundation (NSF 1342183) to Gelsy Torres-Oviedo. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.