
Abstract
Although global movement abnormalities in the lower extremity poststroke have been studied, the expression of specific motor impairments such as weakness and abnormal muscle and joint torque coupling patterns have received less attention. We characterized changes in strength, muscle coactivation and associated joint torque couples in the paretic and nonparetic extremity of 15 participants with chronic poststroke hemiparesis (age 59.6 ± 15.2 years) compared with 8 age-matched controls. Participants performed isometric maximum torques in hip abduction, adduction, flexion and extension, knee flexion and extension, ankle dorsi- and plantarflexion and submaximal torques in hip extension and ankle plantarflexion. Surface electromyograms (EMGs) of 10 lower extremity muscles were measured. Relative weakness (paretic extremity compared with the nonparetic extremity) was measured in poststroke participants. Differences in EMGs and joint torques associated with maximum voluntary torques were tested using linear mixed effects models. Results indicate significant poststroke torque weakness in all degrees of freedom except hip extension and adduction, adductor coactivation during extensor tasks, in addition to synergistic muscle coactivation patterns. This was more pronounced in the paretic extremity compared with the nonparetic extremity and with controls. Results also indicated significant interjoint torque couples during maximum and submaximal hip extension in both extremities of poststroke participants and in controls only during maximal hip extension. Additionally, significant interjoint torque couples were identified only in the paretic extremity during ankle plantarflexion. A better understanding of these motor impairments is expected to lead to more effective interventions for poststroke gait and posture.
Introduction
Stroke is the leading cause of long-term adult disability in the United States with 800 000 new and recurring strokes each year. 1 After a stroke, 80% of survivors can ambulate independently 1 ; however, poststroke stance, balance, and gait differ significantly from normal,2-6 in part because of motor impairments in the voluntary control of the paretic lower extremity. After a stroke, cortical and subcortical lesions in the brain interrupt descending and ascending projections that carry motor and sensory information. Loss of corticofugal projections results in weakness7,8 and may increase reliance on brainstem motor pathways9,10 that project to ventral motor neuron pools of axial, proximal and to a lesser extent distal limb muscles11-14 and innervate neurons over many spinal segments.
Previous studies quantifying paretic weakness and joint torque couples in the lower extremity have reported differing results15-21 from weakness only in hip flexion, abduction, adduction, and ankle dorsiflexion16,17,21 to decreased torque strength of all joints and in all directions (flexion and extension, abduction and adduction19,20). These investigations have reported results ranging from the existence of poststroke abnormal joint torque coupling in the form of a dominant extensor synergy16,18 to absence of stereotypical joint torque coupling patterns in the paretic extremity.19,20 Contrasting results may be explained by the differences in (1) experimental posture and associated muscle length and force generating capabilities,18-20,22,23 based on the force-length curve 24 and (2) weightbearing conditions—motor output will vary if weight is supported on the nontested lower extremity19,20 or when both lower extremities are unloaded. 16 In this study, we examined lower extremity weakness and joint torque coupling patterns in a posture with muscle lengths that resemble those during gait loading response, 25 while providing maximum stabilization of the lower extremity segments. In addition, our experimental configuration allowed the nontested extremity to be fully unloaded and free to move, to reduce interlimb coupling driven by activity in the nontested extremity. By controlling for previous confounding factors we hope to better characterize poststroke impairments in the voluntary control of the lower extremity.
The goal of this study was to characterize poststroke weakness and joint torque coupling patterns by quantifying strength and associated inter (between 1 or more joints) and intra (between degrees of freedom in the same joint) joint coupling. We hypothesized the presence of an extension bias and limited flexion-generating ability in the paretic lower extremity, in agreement with the clinically described lower limb extensor synergy (coupling of hip extension, hip adduction, knee extension and ankle plantarflexion26,27), but not in the nonparetic extremity of poststroke participants or in the lower extremity of control participants, during the generation of maximal and submaximal extension torques. Our findings indicate relative preservation of hip extension/adduction strength, a prevalence of extension synergy muscle coactivation patterns and a bias toward extension/adduction torque coupling in the paretic lower limb.
Methods
Participants
Poststroke participants were recruited for this study from the Clinical Neuroscience Research Registry, and the study was approved by Northwestern University’s institutional review board. Motor impairment was measured using the lower extremity portion of the Fugl-Meyer (F-M) Motor Assessment. 28 The Berg Balance Test 29 and the 10-meter walk test 30 were used to test balance and walking speed, respectively.
Poststroke participants should have had a unilateral brain lesion from a single stroke at least 12 months prior to the experiment. Selection criteria included the following: ability to provide informed consent, paresis confined to one side, cortical, or subcortical lesions not involving the brainstem or cerebellum, as determined from available magnetic resonance imaging scans or reported in clinical records. We also required absence of severe sensory impairments, wasting or contracture, and cognitive or affective dysfunction. Medications known to suppress central nervous system activity, including alcohol were not allowed.
Data from one participant were rejected due to severe edema, which affected sensation and added significant noise to EMG measurements. Data for another participant was rejected due to failure to understand the tasks. The final sample of poststroke participants consisted of 13 community ambulators and 2 household ambulators. Eight age-matched control participants were recruited from the community and took part in the study. Demographic information is shown in Table 1.
Participant Demographics.
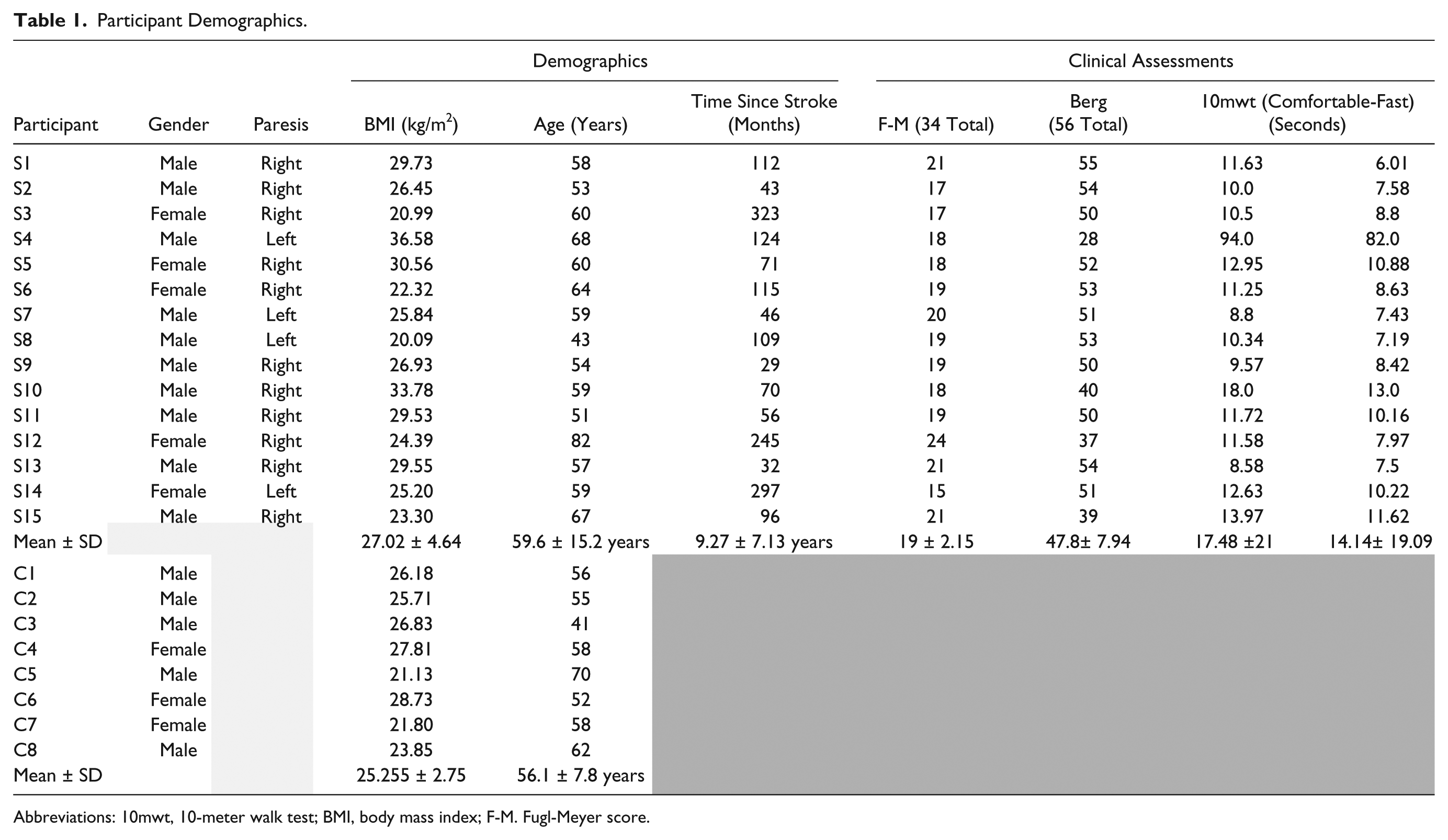
Abbreviations: 10mwt, 10-meter walk test; BMI, body mass index; F-M. Fugl-Meyer score.
Experimental Setup
Participants were fitted into the MultiLEIT 15 (Figure 1). The weight of the upper body was supported by a bicycle saddle and a harness, 15 which also minimized movement. Joint angles for the tested extremity were set to 20° hip and knee flexion, 0° ankle flexion, and 10° hip abduction as those occurring during the unimpaired, healthy loading response phase, 25 and during the loading response phase in poststroke participants defined as slow walkers and reported in a previous study. 31 Each lower extremity of poststroke participants and each task (maximum voluntary torque [MVT] and submaximal task) was tested on separate visits to the lab. All MVTs were consistent between sessions. The right lower extremity of control participants was tested in a single visit to the lab.
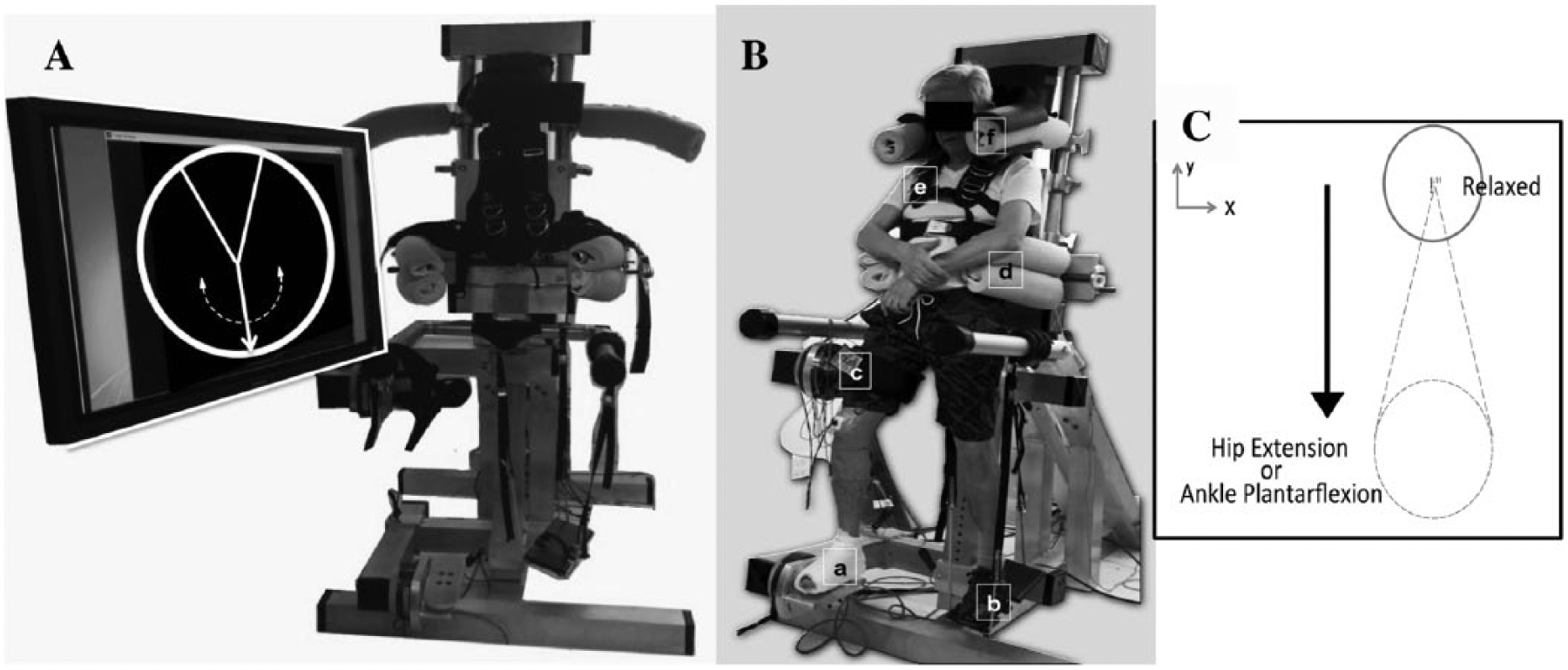
Photograph of the MultiLEIT experimental setup and visual feedback showing the speedometer provided to the participant for the maximum voluntary torque (MVT) task. (A) The vertical arrow rotates clockwise or counterclockwise depending on torque direction. Arrow rotation is mapped to the magnitude of the torque generated. (B) (a) Rigid foot attachment and bottom sensor. (b) Adjustable foot-sole support for the nontested extremity consisting of a bungee cord attached to a placing surface. (c) Adjustable thigh cuff and bottom sensor. (d) Pelvic clamps. (e) Trunk harness (f) Shoulder clamps. (C) Visual feedback for the submaximal task. Vertical displacement and location of the target were mapped to the percentage of hip extension or ankle plantarflexion. The cursor only moved in the vertical direction, all other degrees of freedom were locked.
Torques and forces were measured using two 6–degree-of-freedom (DOF) load cells (Model 75E20A4, JR3 Inc, Woodland, CA) and transformed into joint torques using static Jacobian transformation matrices.15,32 Single differential surface Ag EMG sensors (Delsys Bagnoli, Delsys Inc, Boston MA) were placed on the muscle belly of the gluteus maximus (GMax), gluteus medius (GMed), adductor longus (AddL), the lateral portion of the quadriceps complex (rectus femoris/vastus lateralis [RF/VL] 33 ), vastus medialis (VM), biceps femoris (BF), semimembranosus (SM), medial head of the gastrocnemius (Gastroc), soleus (Sol), and tibialis anterior (TA). The reference electrode was placed on the patella. EMG data were sampled at 1000 Hz.
Experimental Protocol
First, participants practiced the generation of each joint torque outside the setup to understand each of the tasks. Once fitted in the setup, participants were asked to perform MVTs in hip abduction, adduction, flexion, extension, knee flexion, extension, ankle dorsiflexion, and ankle plantarflexion in randomly ordered blocks to eliminate the effects of learning and fatigue. Visual feedback (Figure 1A) was provided only for the primary DOF being maximized. Participants were aided by tactile feedback from the researcher to help with joint torque generation. Consistent instructions were provided for all participants. Multiple trials were performed until torque magnitudes for each DOF plateaued and were sustained for 2 seconds. Voluntary (primary) torques and coupled (secondary) torques generated at the other joints were computed online based on the raw forces and torques measured from the two 6-DOF load cells. Two trials with MVT values within 10% of each other, with the second trial being of smaller magnitude than the first trial were required to ensure MVTs were achieved.
We tested submaximal levels of hip extension and ankle plantarflexion to determine whether the same coupling patterns occurred at submaximal levels. We intended to test submaximal knee extension in order to characterize all extension DOFs, but participants were unable to accurately control and sustain submaximal knee torques. Participants were instructed to generate 25%, 50%, and 75% hip extension and ankle plantarflexion MVTs. Visual feedback was provided only for the submaximal DOF. Participants were asked to start in a relaxed state, ramp up to the desired torque level, and sustain it for 2 seconds within ±5% of the target torque (Figure 1C).
Signal Processing and Data Analysis
Data acquisition was performed using a National Instruments DAQ (PCI 6031E, National Instruments, Austin, Texas). Signal processing and data transformations were performed in a custom Matlab graphical user interface (Mathworks Inc., Natick, MA). Data were acquired for 8s using a 1000 Hz sampling frequency and a 500 Hz anti-alias filter.
Force and torque data were smoothed using a 250ms moving average window. The MVT for each task was obtained from a 250ms interval where the torque magnitude was held constant. Mean secondary torques were extracted for the same time interval. Torques were normalized to the maximum torque in each DOF obtained during the entire experiment, either primary or secondary.
All statistical analyses were run in SPSS (SPSS Inc., Chicago, IL). Data were tested for normality using Q-Q plots. Homogeneity of variance was tested using Levene’s Statistic. Normalized coupled torques were compared between groups using a nested linear mixed effects model for each DOF and for each task, to test significant main effects of group (Stroke, Control), lower extremity (Control, Nonparetic, Paretic) and interactions. Participants (C1 to C8 and S1 to S15) were the random factors. Post-hoc comparisons were run using least significant difference (LSD) analyses on the estimated marginal means for each fixed effect factor. Analyses were run for a sample including all participants and a sample without participant S4 given the differences in clinical metrics observed for this participant. Results did not differ when S4 was excluded, therefore participant S4 was included in all analyses.
A linear mixed effects model was also run for the submaximal data, with the levels of torque as a repeated-measures fixed factor to test for significant effects of group, lower extremity, and level on secondary torque magnitudes.
The relative joint torque weakness (RTW) ratio was calculated for each participant as
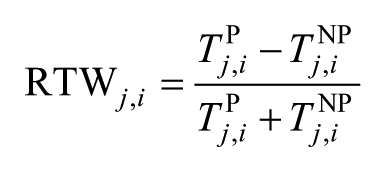
for each joint j, in each DOF i. NP and P stands for nonparetic and paretic, respectively. A RTW ratio equal to 0 indicates equal strength, a negative RTW ratio indicates loss in paretic strength and a positive RTW indicates a stronger paretic extremity. One-sample t tests were used to determine significant differences from 0 for the RTW.
EMG data were baseline corrected, full wave rectified and smoothed using a 250-ms moving average window. The mean EMG amplitude corresponding to the MVT was identified and averaged for the 50 ms leading to the time-point of MVT for each task. We tested the repeatability of muscle activation measurements based on raw EMG in the paretic extremity by computing the difference in EMG magnitude between the 2 trials that qualified as MVTs. Variability within participants was identified and then averaged across participants to test for repeatability of EMG measurements.
EMGs were normalized to the peak maximum activation obtained for the task in which the muscle was expected to provide the greatest joint torque contribution based on anatomical action and moment arm 34 and expressed as %MVT-EMG. EMG normalization is necessary to account for within and across individual variability due to electrode placement on the muscle, the amount of subcutaneous fat, skin impedance, and differences in voluntary muscle activation between participants. Normalizing EMG by the muscle primary action across all participants resulted in a homogeneous baseline, such that deviations from 100% are indicative of modulation in voluntary drive to the muscle when used as a primary or secondary task effector. A %EMG-MVT value greater than 100% indicates that the muscle was more active as a secondary task effector. The tasks used to normalize the EMGs were hip abduction for GMed, hip adduction for AddL, knee extension for RF/VL and VM, knee flexion for BF, SM, Gastroc, hip extension for GMax, ankle plantarflexion for Sol and ankle dorsiflexion for TA. Statistical analyses were run as in the MVT task.
Results
Torque Weakness
Joint torques normalized to body mass index (BMI) and associated relative torque weakness (RTW) results are summarized in Table 2. Significant differences in both the paretic and nonparetic hip extension torque compared with controls were observed and the RTW in hip extension did not differ significantly from zero. Overall, maintenance of proximal strength, relative to the nonparetic extremity was seen in this cohort of ambulators with hemiparetic stroke.
Torque Magnitudes Normalized by Body Mass Index (BMI) and Relative Joint Torque Weakness (RTW), Expressed as Mean ± Standard Deviations. a
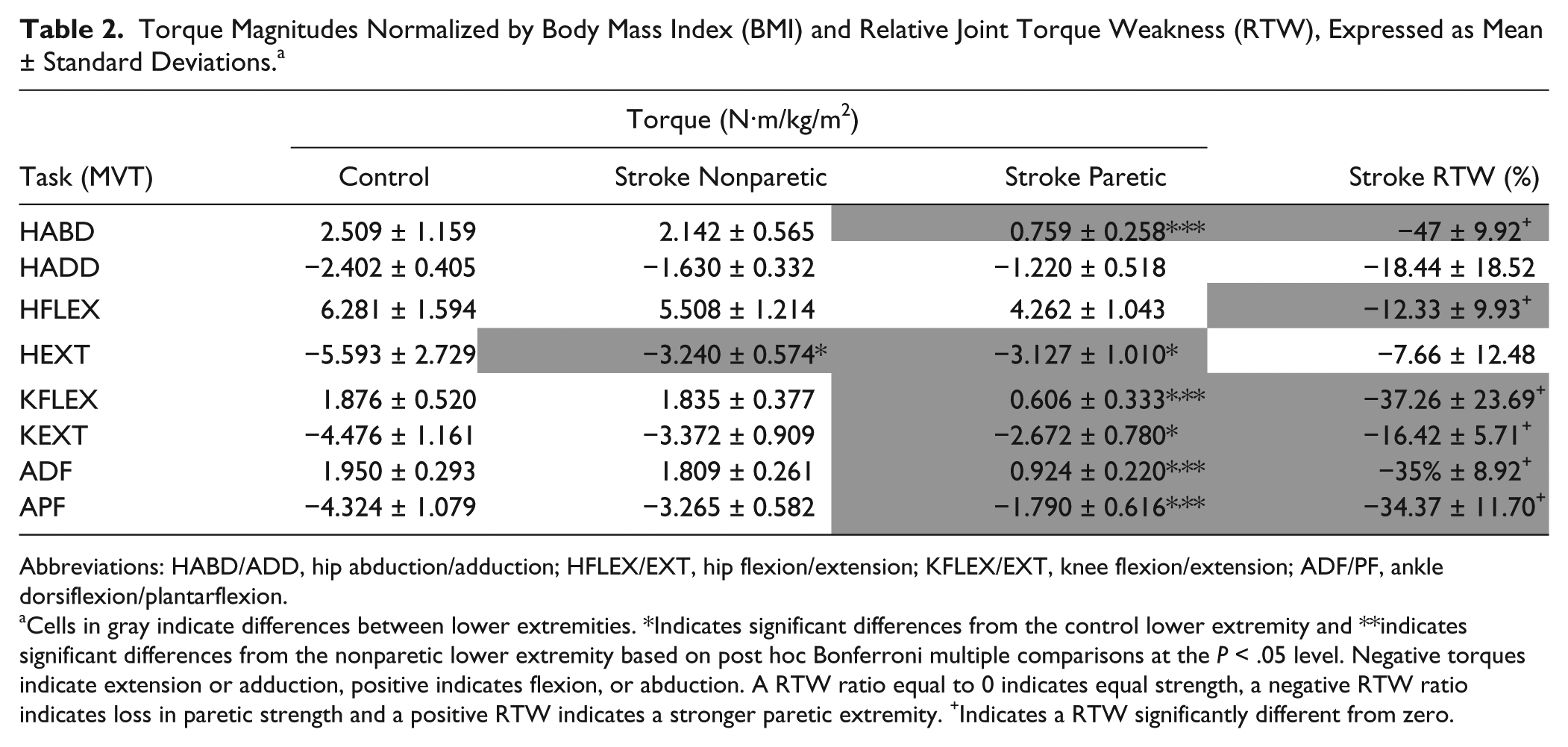
Abbreviations: HABD/ADD, hip abduction/adduction; HFLEX/EXT, hip flexion/extension; KFLEX/EXT, knee flexion/extension; ADF/PF, ankle dorsiflexion/plantarflexion.
Cells in gray indicate differences between lower extremities. *Indicates significant differences from the control lower extremity and **indicates significant differences from the nonparetic lower extremity based on post hoc Bonferroni multiple comparisons at the P < .05 level. Negative torques indicate extension or adduction, positive indicates flexion, or abduction. A RTW ratio equal to 0 indicates equal strength, a negative RTW ratio indicates loss in paretic strength and a positive RTW indicates a stronger paretic extremity. +Indicates a RTW significantly different from zero.
Joint Coupling and Muscle Coactivation During MVTs
None of the MVT tasks in the abduction or flexion directions elicited the full flexor synergy (coupling between hip flexion, abduction, knee flexion, and ankle dorsiflexion26,27). In contrast, we identified torque coupling as that described in the extensor synergy 26 during hip extension and ankle plantarflexion. However, joint torque coupling patterns during hip extension did not differ between controls and the paretic and nonparetic extremity of poststroke participants. Significant adduction torques measured during maximal hip extension and significant hip extension and adduction torques during ankle plantarflexion in poststroke participants are consistent with the 2 DOFs with preserved strength reported in our weakness analyses. All findings for the MVT task and multiple comparisons results are listed in Table 3.
Joint Torque Coupling for All Tasks and Muscle Activation for the Paretic Extremity. a
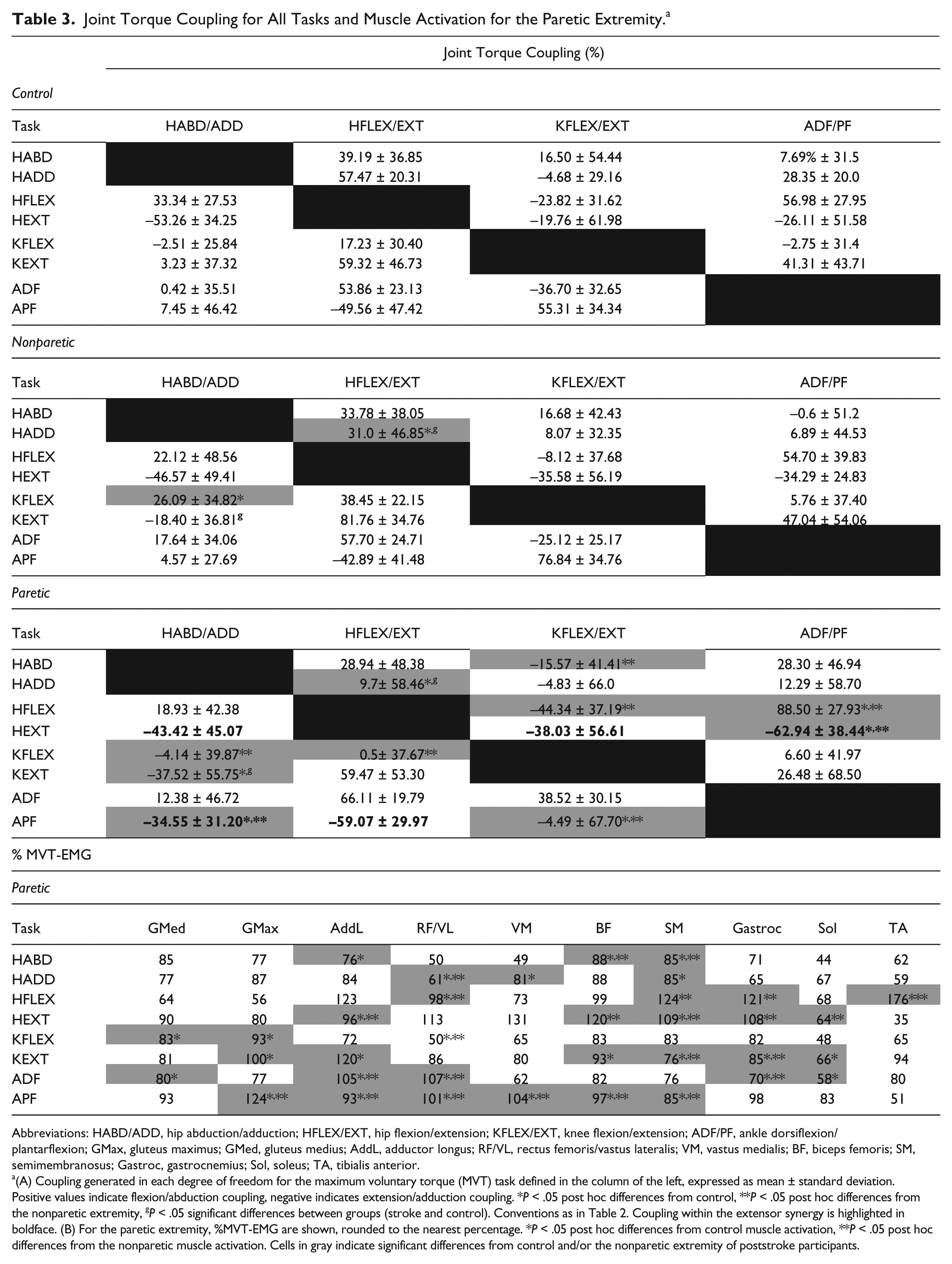
Abbreviations: HABD/ADD, hip abduction/adduction; HFLEX/EXT, hip flexion/extension; KFLEX/EXT, knee flexion/extension; ADF/PF, ankle dorsiflexion/plantarflexion; GMax, gluteus maximus; GMed, gluteus medius; AddL, adductor longus; RF/VL, rectus femoris/vastus lateralis; VM, vastus medialis; BF, biceps femoris; SM, semimembranosus; Gastroc, gastrocnemius; Sol, soleus; TA, tibialis anterior.
(A) Coupling generated in each degree of freedom for the maximum voluntary torque (MVT) task defined in the column of the left, expressed as mean ± standard deviation. Positive values indicate flexion/abduction coupling, negative indicates extension/adduction coupling. *P < .05 post hoc differences from control, **P < .05 post hoc differences from the nonparetic extremity, gP < .05 significant differences between groups (stroke and control). Conventions as in Table 2. Coupling within the extensor synergy is highlighted in boldface. (B) For the paretic extremity, %MVT-EMG are shown, rounded to the nearest percentage. *P < .05 post hoc differences from control muscle activation, **P < .05 post hoc differences from the nonparetic muscle activation. Cells in gray indicate significant differences from control and/or the nonparetic extremity of poststroke participants.
During MVT, greater coactivation of paretic AddL was measured during all extension tasks and surprisingly, during hip abduction and ankle dorsiflexion. Greater co-contraction of extensor musculature was measured during all extension tasks, that is, hip extension, knee extension, and ankle plantarflexion. As shown in Table 3, coactivation was higher as the joint torque maximized became more distal. Finally, in the paretic extremity, coactivation of musculature was greater in magnitude than that during volitional activation. EMG activations measured in the paretic extremity, and expressed as %MVT-EMG are summarized in Table 3.
Of particular interest are the hip extension and ankle plantarflexion MVT results. Both these tasks elicited the full extensor synergy, with coupling of hip extension, adduction, knee extension and ankle plantarflexion and significant coactivation of all extensor musculature with AddL (Figure 2). For hip extension however, coupling patterns did not differ between groups, despite a stronger ankle plantarflexion coupling measured in the paretic extremity (F = 4.343, P = .045). During plantarflexion, hip adduction torques were significantly greater in the paretic extremity compared with both the nonparetic extremity and controls (F = 9.793, P = .004). In controls and in the nonparetic extremity, ankle plantarflexion generated knee flexion coupling, whereas in the paretic extremity, mean coupling was biased toward knee extension (F = 19.204, P = .001).
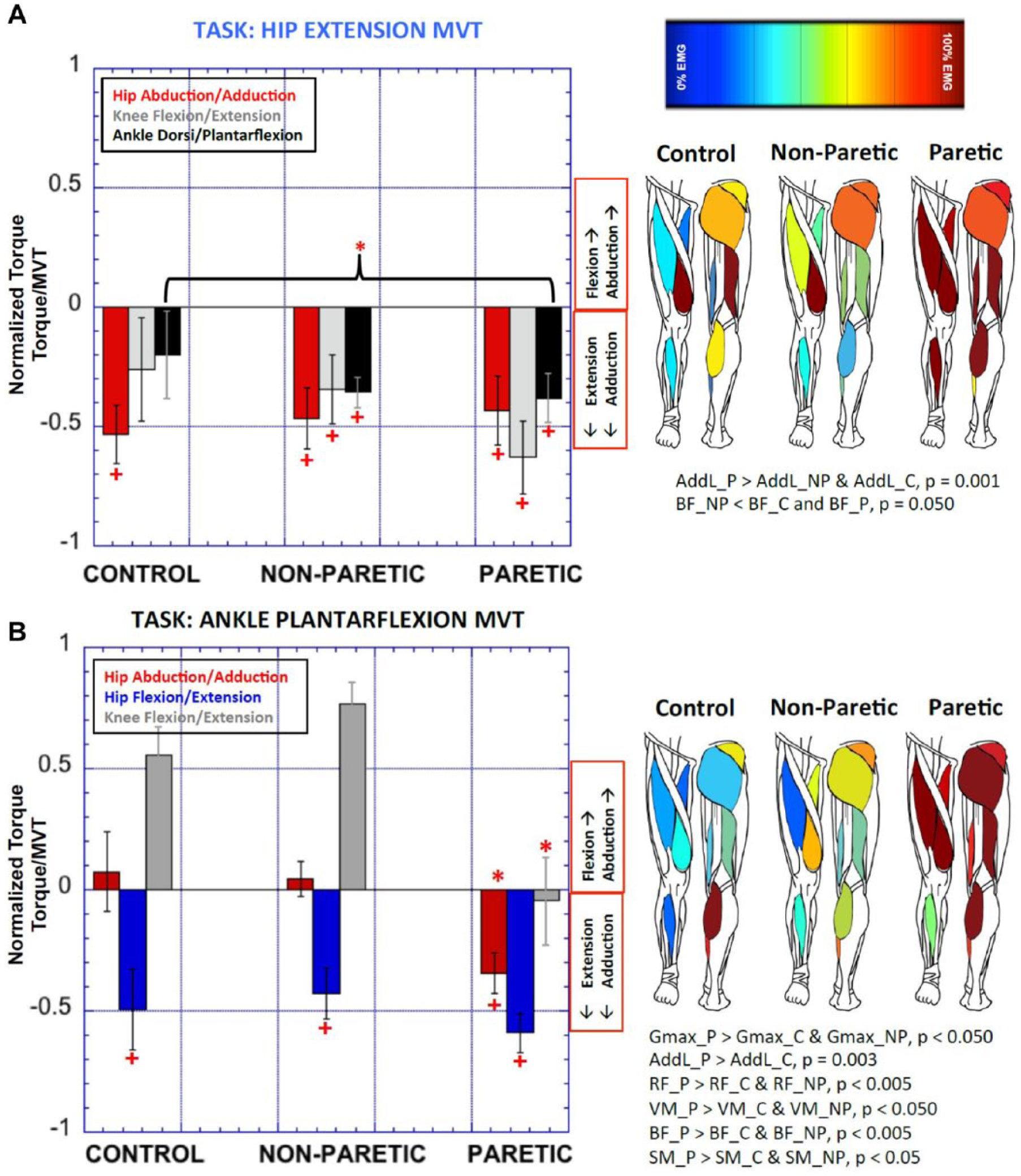
Spontaneous joint torque coupling patterns during hip extension (A) and ankle plantarflexion maximum voluntary torque (MVT) (B) and underlying electromyography (EMG) activation. All torques, both voluntary and spontaneous are shown as a percentage of the global maximum generated during the entire experiment. The column on the right shows the EMG activity generating the torque patterns. %EMG activation are coded from 0% EMG (blue) to 100% EMG (red). Significant differences between groups are indicated by *P < .05. Group average secondary torques significantly different from zero are indicated by the +. P, paretic; NP, nonparetic; C, control.
Attempting to maximize distal dorsiflexion torque, which is severely impaired in the paretic extremity, generated coactivation of all paretic musculature. GMed activation was 80% MVT-EMG compared with controls 29% MVT-EMG, P = .059. AddL activation accounted for 105% MVT-EMG, (compared with 54% MVT-EMG in controls, P = .024), RF/VL activation was 107% MVT-EMG (compared with 18% MVT-EMG in controls, P = .006). Antagonist Gastroc activation was 70.5% MVT-EMG, compared with 27% MVT-EMG in controls, P = .011 and 29% MVT-EMG in the nonparetic extremity, P = .004 and SOL activation was 58%MVT-EMG compared with 11% in controls (P = .028). However, despite the significant coactivation, no differences in joint torque coupling patterns were seen between lower extremities during maximal dorsiflexion.
Repeatability measures of raw EMG signal magnitude (in mV), indicated variability within trials smaller than 15% for all muscles and for all tasks except hip extension and knee flexion. The intertrial variability for the GMax during hip extension was 28.7% (±26.86%). During knee flexion, intertrial variability for the BF was 40.57% (±70.3%) and for the SM 45.49% (±101.3%).
Coupling During Submaximal Tasks
Submaximal Hip Extension
Mixed model analyses indicated a significant main effect of hip extension level on hip adduction torque (F = 11.413, P < .001). The proportional increase in hip adduction with hip extension did not differ between groups (F = 0.689, P = .408) but tended to be greater in the paretic extremity (F = 2.742, P = .069) compared with the nonparetic extremity (LSD P = .031) (Figure 3A). For the ankle joint, significant differences were observed between groups (F = 20.256, P < .001), with the control lower extremity generating significantly smaller coupling (F = 18.330, P < .001) at different levels (F = 13.602, P < .001) (Figure 3A).
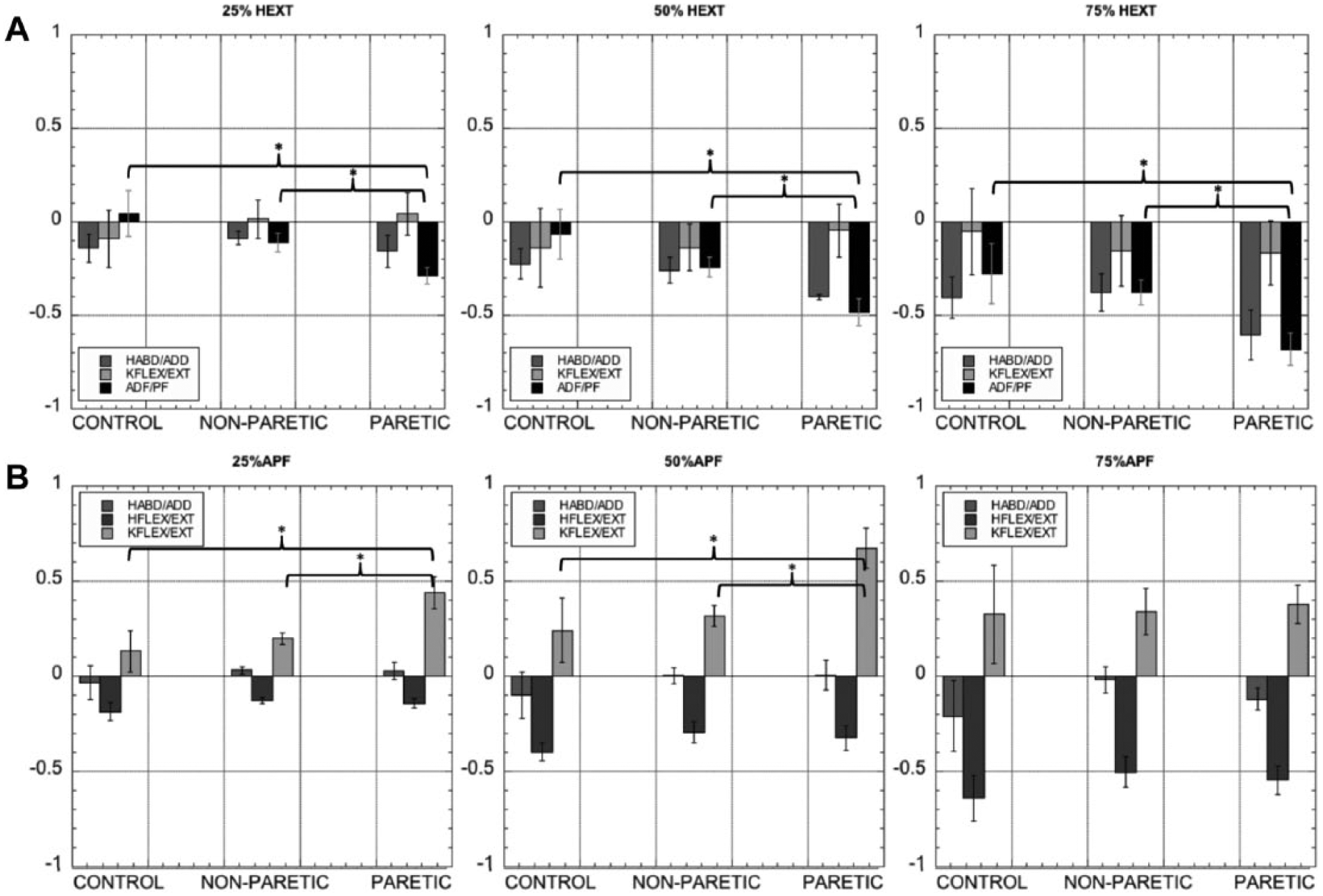
Single degree of freedom submaximal hip extension (A) and plantarflexion (B) tasks. Each level is plotted in a different figure. Torques are presented as a percentage of maximum voluntary torque (MVT) for each group. y-axes express spontaneous torques as a %MVT. *Indicates significant differences between lower extremities at a P = .05 level. HABD/ADD, hip abduction/adduction; HFLEX/EXT, hip flexion/extension; KFLEX/EXT, knee flexion/extension; ADF/PF, ankle dorsiflexion/plantarflexion.
EMG activation during submaximal hip extension
GMed, GMax, RF, VM, and BF activation increased as hip extension increased (P < .005) with no significant differences between control, paretic, and nonparetic extremities. The greater GMed activation may have been an effect of crosstalk due to a greater GMax activation compared with GMed. AddL EMG was higher for greater levels of hip extension (F = 22.401, P < .001). This activation was different across all groups (F = 6.252, P = .004), with a greater activity of paretic AddL compared with control and nonparetic (F = 5.012, P = .014 for group).
Submaximal Ankle Plantarflexion
No main effects of group or lower extremity was seen for the coupling between plantarflexion and hip abduction/adduction and hip flexion/extension. Coupling with the knee was significantly different between lower extremities, with the paretic lower extremity generating significantly greater knee flexion torques compared with controls (P = .016) and the nonparetic extremity (P = .016) (Figure 3B).
EMG activation during submaximal ankle plantarflexion
GMed activation increased as the plantarflexion level increased (F = 4.285, P = .043) for the control, paretic, and nonparetic muscle (F = 1.096, P = .368). AddL was more active in the paretic extremity at 50% and 75% MVT (F = 5.080, P = .006) compared with controls (F = 5.372, P = .026). Greater paretic VM activation compared with its nonparetic counterpart was observed for 25%, 50%, and 75% plantarflexion (F = 26.402, P < .001 for level and F = 5.203, P = .028 for group).
Discussion
Here, we measured decreased paretic hip abduction and distal strength, and preserved paretic hip extension and hip adduction strength. Furthermore, abnormal joint torque coupling between hip extension and hip adduction was observed in the paretic extremity. Our findings also show that muscle coactivation patterns, especially in the paretic extremity and less so in the nonparetic extremity, combine muscles that are part of the extension synergy when generating torques in various degrees of freedom at the hip, knee, and ankle. EMG analyses indicate a persistent activation of paretic hip adductor musculature which supports the maintained strength of this DOF and may help explain the decreases in net hip abduction torques. During stance, significant joint torques are needed to maintain the body upright and stable while progressing the body forward over the supporting extremity and while counteracting perturbations. Coupling of extension with adduction and impaired abduction may affect stance requirements: our findings explain some aspects of the deficits seen during paretic stance, mainly reduced stance times 4 reduced ability to bear weight on their paretic extremity, 35 decreased forward progression, 36 and pelvic drop. 25
Weakness
Hip extension torques were decreased bilaterally in poststroke participants, potentially due to the overall decrease in physical activity in this population, therefore masking relative weakness, which does not account for nonparetic weakness. We measured reduced RTW in all degrees of freedom but hip extension/adduction. For all our ambulating stroke participants, whenever they use their hip extensors coactivation with hip adductors may potentially prevent disuse-induced muscle atrophy. 37 Also, despite the fact that hip flexion torque magnitude did not differ significantly from controls, the normalized RTW was significantly different from zero, potentially because of reductions in variability after normalizing the data.
Our results indicate that hip abduction was the most impaired DOF, possibly due to decreased drive to the paretic abductors and greater coactivation with adductors. Hip abduction strength is important during the stance phase of gait to counteract contralateral pelvic drop and maintain a semineutral pelvic orientation during stance. 25 Hip abduction weakness and hip adduction bias may help explain mediolateral balance impairments during stance.
Weakness was also significant for all flexion DOFs, possibly a consequence of an extension bias and the associated extensor muscle coactivation patterns. Based on previous work in nonhuman primates,38-40 where ablation of the corticospinal pathways generated increased hip extensor/adduction tone, we postulate that the overall flexion weakness is due to decreased flexion/abduction cortical drive and greater brainstem-mediated postural drive required to maintain the body in an upright position.
Weakness of extensor DOFs was observed only in the more distal joints. We postulate that distal weakness is a result of losses of more prevalent direct corticomotoneuronal connections to distal lower extremity muscles, such as those that control the knee 41 and ankle joints.42-44
Muscle Coactivation Patterns
Poststroke changes in muscle coactivation patterns were supported by maximum activation of paretic and nonparetic muscles that was produced during the generation of secondary torques and not primary torques, as shown by muscle activation of more than 100% MVT-EMG, particularly during the tasks involving the knee and ankle. Loss of corticospinal pathways that connect directly to motorneurons of distal muscles may decrease specific volitional drive to these muscles. To satisfy the strength requirements of the task, muscles may have been recruited by drive from diffuse bulbospinal pathways that branch across multiple spinal segments, therefore sacrificing specificity for strength. This was particularly the case during ankle plantar and dorsiflexion. Abnormal coactivation patterns with a bias in extension not only decrease specificity in motor control but also demand a higher energetic cost to achieve the required torques. 45
EMG results also indicated an abnormal persistent activation of the AddL during hip abduction, knee extension, ankle dorsiflexion, and during the synergistic tasks ankle plantarflexion and hip extension in the paretic lower limb. Results also show high crosstalk between hip musculature (GMed and GMax) during maximal torque conditions, as seen by the simultaneous increase of GMed activation even when no hip abduction torque was generated, as during dorsiflexion MVT. It is possible that muscle activation in the GMed was underestimated due to crosstalk from the adjacent GMax muscle. Future research should use intramuscular EMG to measure GMed activity and avoid crosstalk contamination.
Coupling During Flexion Tasks
The results from this study do not fully reflect what has been described clinically as the flexor synergy,26,27 but instead, the observed coupling was consistent with the action of biarticular muscles. Note that the flexor synergy has been described while the person is in a supine posture. The neuromechanical requirements during the upright posture used here (vestibular drive and the vertical reaction forces on the tested foot), may spontaneously drive the knee into an extension torque and inhibit the flexor muscles in the supporting limb 46 as a postural response to maintain an upright posture. We do not rule out the existence of the flexion synergy in other postures but instead affirm the absence of flexion/abduction coupling during upright stance.
Extension/Adduction Bias During Hip Extension
Muscle coactivation patterns reflected the joint torque coupling in accordance with the clinically described extensor synergy 26 during maximal and submaximal levels of hip extension; surprisingly, coupling patterns did not differ between control and poststroke participants. Group similarities in coupling patterns may be due to the moment arm contributions to each joint torque in the posture implemented in the experimental setup and the fact that our experimental posture could in fact promote hip adduction coupling and extension of the knee and ankle as an effective antigravity strategy. More specifically, the 10° of hip abduction augment the moment arm contributions of the hip adductor muscles to the hip extension moment, even in control participants. Future work must determine whether participants can generate torque couples outside of the hip extension/adduction pattern.
Significant differences in muscle coactivation in both the paretic and nonparetic extremities of poststroke participants support the hypothesis that altered neural drive to the lower extremity is a main contributor to the observed coupling. Higher muscle coactivation seen in stroke may be due to an increased reliance on brain stem pathways such as the lateral vestibulospinal tract that coordinates and adjusts the activity of the ipsilateral 47 and contralateral extensor muscles 48 and the reticulospinal tract, which elicits bilateral task-specific excitatory post-synaptic potentials of extensor motorneurons. 49 These findings are supported by the submaximal hip extension task; poststroke participants seem to break out of the full extensor pattern when the muscle activation requirements are less, allowing for the mechanical coupling from biarticular muscles to overcome the abnormal neural coupling in the paretic lower extremity. This is in fact supported by research indicating that the ability to coordinate relative muscle activity after stroke is preserved. 50 We would like to further expand this statement by specifying that this capacity is preserved as long as the strength requirements of the task are submaximal. As shown here, as the torque generating requirements increase, the neural drive to the paretic extremity musculature must also increase through any remaining resources, possibly via brainstem postural pathways,38,51 thus sacrificing independent joint control and coordination of muscle activity for strength.
Our results are in agreement with previous isometric and isokinetic studies that showed differences in the direction of endpoint forces in the paretic lower extremity compared with the nonparetic extremity 50 : these findings can be summarized as a force line-of-action shifted closer to the knee in the sagittal plane and closer to parallel to the midline in the frontal plane. These changes in the force line of action can be explained by the torque coupling observed here. With regard to the functional implications of our results, our findings may explain why more than 50% of falls in individuals poststroke result in fractures of the paretic side 52 : the hip extension torque required to maintain the body upright to counteract a fall, may in turn generate an associated adduction torque which could lead the body to collapse toward the paretic side (if the trunk fails to compensate to counteract the fall).
Extension/Adduction Bias During Ankle Plantarflexion
During paretic ankle plantarflexion, muscle coactivation was observed when attempting to maximize this distal joint torque. Note that the observed coupling followed the extensor synergy at maximal conditions but not during submaximal efforts, indicating that the coupling from distal to proximal joints is weaker compared with the coupling from proximal to distal joints, in agreement with previous upper extremity work. 53
One of the limitations of this study is the use of the experimental setup with multiple contact points, unusual task perception and discomfort during the experiment which may have increased variability across participants. However, the benefits of the setup, mainly stabilization of the trunk and pelvis during maximal exertion, and removing the need to support the body allowed for generation of true MVTs during upright posture. To mitigate task perception biases, participants first performed the task outside the setup so that full understanding of the task was ensured. Finally, between trials, participants rested inside the setup in a supine posture to reduce discomfort. We believe that these measures helped mitigate biases from the experimental setup in our results, as supported by the repeatability of measures obtained across multiple sessions.
Limitations of this study also include high variability in EMG magnitudes for the GMax, BF, and SM during tasks where these muscles were the primary effector. For the specific case of the GMax, the amount of subcutaneous fat as well as the experimental setup, which may have created motion artifacts at the electrode when muscle volume changed, could have reduced signal strength and added noise to the signal, respectively. In the case of the muscles for knee flexion, we expected high variability due to the inherent knee flexion impairment in this chronic sample. 54 Despite the presence of reduced signal strength and increased noise, our primary hypotheses are supported by abnormal activation of muscles where we obtained robust measurements.
Our results provide an insight into an altered neural drive to the lower extremity poststroke. The presence of flexion/abduction weakness, and a bias toward extension/adduction coupling, may be a consequence of an increased reliance on brainstem pathways across this cohort of ambulatory chronic poststroke participants. Results from this study suggest that the common deficits seen in this population can be addressed via rehabilitation interventions that focus on strengthening flexion and abduction degrees not in isolation but in combination with extension tasks.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Institute of Health Grant R01HD039343 to JPA Dewald and American Heart Association Pre-doctoral Fellowship 13PRE14690048 to N Sánchez.