
Abstract
Background. Following traumatic brain injury (TBI), clinical cognitive training paradigms harness implicit and explicit learning and memory systems to improve function; however, these systems are differentially affected by TBI, highlighting the need for an experimental TBI model that can test efficacy of cognitive training approaches. Objectives. To develop a clinically relevant experimental cognitive training model using the Morris water maze (MWM) wherein training on implicitly learned task components was provided to improve behavioral performance post-TBI. Methods. Eighty-one adult male rats were divided by injury status (controlled cortical impact [CCI]/Sham), non-spatial cognitive training (CogTrained/No-CogTrained), and extra-maze cues (Cued/Non-Cued) during MWM testing. Platform latencies, thigmotaxis, and search strategies were assessed during MWM trials. Results. Cognitive training was associated with improved platform latencies, reduced thigmotaxis, and more effective search strategy use for Sham and CCI rats. In the Cued and Non-Cued MWM paradigm, there were no differences between CCI/CogTrained and Sham/No-CogTrained groups. During novel testing conditions, CogTrained groups applied implicitly learned knowledge/skills; however, sham-cued CogTrained/rats better incorporated extramaze cues into their search strategy than the CCI-Cued group. Cognitive training had no effects on contusion size or hippocampal cell survival. Conclusions. The results provide evidence that CCI-CogTrained rats that learned the nonspatial components of the MWM task applied these skills during multiple conditions of the place-learning task, thereby mitigating cognitive deficits typically associated with this injury model. The results show that a systematic application of clinically relevant constructs associated with cognitive training paradigms can be used with experimental TBI to affect place learning.
Keywords
Introduction
After traumatic brain injury (TBI), many cognitive rehabilitation treatments can improve outcomes. 1 However, evidence-based clinical practice guidelines recommend treatments tailored to address residual cognitive deficits after TBI. 1 Memory remediation strategies include errorless learning 2 and compensatory strategy training, 3 each demonstrating that process-oriented strategy training can be adapted to situations with different memory requirements for improved functional performance. 3 Furthermore, some cognitive rehabilitation paradigms demonstrate carryover effects, even after treatment discontinuation. 4 These findings indicate that patients with TBI can learn and apply skills in novel situations, despite limitations within other cognitive domains.
This learning may occur through separate but interactive implicit and explicit learning and memory networks that facilitate information acquisition, storage, and retrieval. 5 Explicit learning/memory represents the conscious acquisition/recall of information, including spatial mapping and directionality. Implicit learning/memory represents the subconscious acquisition/recall of procedural skills (including search strategy formulation), stimulus–response associations, perceptual priming, habituation, and sensitization. 6 Implicit networks are fundamental to functional skill training and are relatively preserved after injury, while explicit memory networks are relatively impaired. Many cognitive treatment paradigms leverage relatively intact implicit learning/memory systems to remediate other impaired systems. However, explicit feedback can impair the use of implicitly learned skills.7,8 Our recent work used implicit pretraining in the Morris water maze (MWM) to dissect the cognitive components of behavioral deficits observed with place learning after experimental TBI 9 ; implicit learning/memory were relatively intact after experimental TBI, while explicit learning/memory were impaired. The data also suggested that implicit training after injury may be a viable method for improving new learning/memory after experimental TBI.
Presently, there is no well-characterized, clinically relevant model of implicit cognitive training (rehabilitation) after experimental TBI. However, such a model could facilitate our understanding of learning/memory impairment mechanisms after TBI and how pharmacotherapies work in the context of clinically relevant cognitive training constructs. The MWM is a commonly used test of learning and memory10,11 that engages both implicit and explicit learning/memory networks to solve a spatial navigation task.9,12 When contextualized to the MWM, explicit learning/memory includes spatial map acquisition (ie, escape platform location relative to extra-maze cues). Implicit learning/memory includes the ability to suppress thigmotaxis, search the pool, and recognize the escape platform as the solution to the task. 12
We developed a postinjury cognitive training model that is relevant to clinical cognitive rehabilitation paradigms, that introduces rats post-injury to the implicit elements of the MWM, and that examines therapeutic effects on both implicit and explicit aspects of place learning with this task. We hypothesized that our cognitive training paradigm would alleviate the implicit learning burden associated with place learning. Based on the clinical cognitive rehabilitation literature, and previous work with implicit training in the MWM prior to injury, 9 we hypothesized that (a) implicit networks would be comparatively intact and responsive to training, (b) postinjury cognitive training would alleviate the implicit learning burden associated with place learning in the MWM task, and (c) non-spatial strategy use acquired with cognitive training after controlled cortical impact (CCI) could be applied effectively to multiple testing conditions. We also explored whether cognitive training would facilitate the incorporation of extra-maze cues to solve the task across various testing conditions.
Methods
Experimental Groups and Controlled Cortical Impact Surgery
Animal procedures were conducted with approval from the University of Pittsburgh Institutional Animal Care and Use Committee. Eighty-one young adult male Sprague–Dawley rats were housed with ad libitum access to food/water, an ambient temperature (21°C ± 1°C), and a 12-hour light:dark cycle. Behavioral testing was conducted between 7:00 and 11:00
The CCI injury device (Pittsburgh Precision Instruments Inc, Pittsburgh PA) is described by Dixon et al 13 and has been used previously by our group.9,14 The device consists of a small-bore double-acting stroke-constrained pneumatic cylinder with a 5-cm stroke. The lower rod end has an impact tip with a diameter of 6 mm, and the impactor tip is pneumatically driven at a preprogrammed velocity, depth, and duration of tissue deformation. Under anesthesia, animals were secured in a stereotactic frame and underwent unilateral parasagittal CCI or sham surgery. For CCI, the impact was delivered to a depth of 2.8 mm at 4.0 m/s. Craniotomy (6 mm diameter) was performed between bregma and lambda. The tissue deformation was ~2.8 mm below the dural surface. Shams underwent the same procedures, including craniotomy, except the CCI injury. Core body temperature during surgery was maintained at 37.0°C ± 0.5°C. The behavioral deficits observed in this study/injury model are consistent with a moderately severe TBI. Initial injury severity was assessed post-operatively by monitoring righting reflex time. A schematic representation of research procedures is provided (see Supplemental Table 1).
Motor Testing
The beam balance and walking tasks were completed to assess motor function. 9 Animals were pretrained on both tasks 1 day prior to surgery, and baseline performance was established on the day of surgery. Rats completed 3 trials per day for each task on postoperative days 1 to 6. A maximum of 60 seconds per trial was allotted. Average time across daily trials was calculated.
Morris Water Maze Testing
The MWM apparatus and place learning task has been previously described. 9 The pool was located in a room (2.5 × 2.5m) with prominent visual cues fixed to the walls. For each trial, rats were introduced into the water (26°C ± 1°C) hindquarters first, facing and with forelimbs on the pool wall. On all testing days, there were 4 trials and a 4-minute intertrial interval. All trials had a maximum duration of 120 seconds, and 30-second platform habituation. If rats did not locate the platform in <120 seconds, they were guided there by hand for 30 seconds of habituation. The platform was submerged 2 cm below the surface of the water, unless otherwise noted.
Cognitive training was provided to a subset of both CCI and Sham rats on postoperative days 8 to 13 and included 4 trials per day. The goal of training was to learn the implicit components of the MWM task (eg, thigmotaxis suppression, swimming strategy formulation, recognizing the platform as the solution to the task, climbing on to the platform) and is analogous to (implicit) procedural training in clinical populations.12,15 Rats began each trial at a static entry point (north), and the submerged platform was randomly rotated to a different quadrant between each trial. Training was conducted in the same testing room as all other conditions of the MWM task, except distal cues were covered, creating an essentially homogeneous environment. Cue covers matched the color of the walls of the testing facility, and the covers were flush with the walls to eliminate possible shadows or other variations that could provide incidental spatial cues. To minimize treatment differences between CogTrained and No-CogTrained groups, the No-CogTrained rats were handled daily and exposed to the warming box when collecting daily weights.
Cognitive training status and inclusion criteria for MWM analysis was conducted to ensure that only rats successfully acquiring implicitly learned non-spatial navigational skills during cognitive training were included in subsequent MWM trial analysis as outlined. We conducted a 2-part post hoc analysis where we assumed that rats having ≥3 trials with 120-second latencies on the final day of training did not learn the essential non-spatial skills (n = 9 rats). We then determined the average peripheral-zone time allocation (TA) as 70% for rats with more than or equal to three 120-second trials on postoperative day 13, and eliminated from MWM analysis 7 of these rats, whose average peripheral-zone TA was ≥70% on postoperative day 13 (n = 3 CCI/CogTrained/Cued; n = 3 CCI/CogTrained/Non-Cued; n = 1 Sham/CogTrained/Non-Cued).
Quantitative Morris Water Maze Analysis
All MWM trials were recorded using the SMART video tracking system from which multiple measures were extracted and compiled as outlined previously. 9 TA in the pool periphery (peripheral-zone; 29% of total pool area) was recorded to quantify thigmotaxis. No portion of the escape platform was located in the peripheral-zone (Figure 1). The presence of extra-maze cues tests (explicit) spatial learning and memory for the location of a hidden platform.
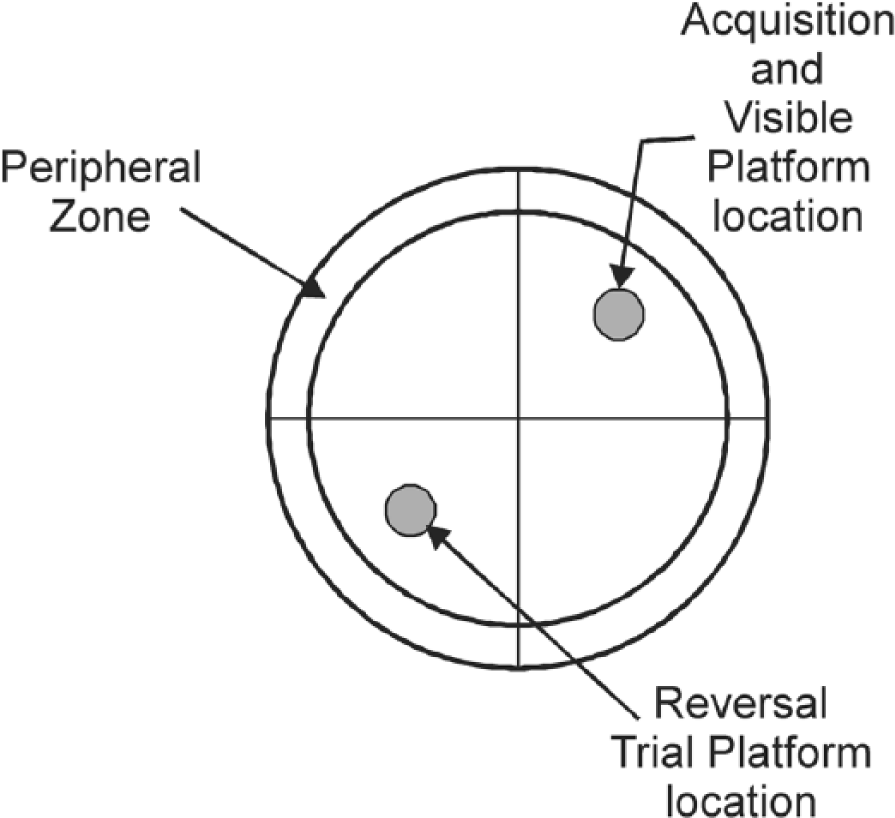
Schematic diagram of Morris water maze pool and zone definitions.
Acquisition trials (4 trials per day, pseudo-randomized dynamic entry point [North–South–East–West] and static platform location [South–East quadrant]) occurred on postoperative days 14 to 18. Rats were tested with or without extra-maze cues. Platform latencies for each trial were averaged daily. Also, average swimming speeds (cm/s) were recorded on postoperative day 14 for group comparison.
Visible platform (VP) trials (4 trials, pseudo-randomized dynamic entry point, static platform location [South–East]) raised 2 cm above the water, and made visible with white tape, were completed on postoperative day 19. While VP trials can be used as a control condition to test for sensorimotor and motivational deficits, the VP also is an egocentric cue from which to formulate spatial search strategies.
On postoperative days 20 and 21, reversal platform (R1-R2) trials were completed as acquisition trials with the platform moved to the North–East quadrant to measure flexibility (ie, the ability to learn a novel platform location).
Qualitative Swim Strategy Analysis
Qualitative swim strategy analyses were completed for postoperative days 14 and 18 acquisition, VP, and both reversal trials (1319 total trials) using a static image of the swim path for each trial. An investigator blinded to treatment group categorized the predominant searching strategy exhibited during each trial based on criteria adapted from previously used classification schemes16-18 as described by Wagner et al. 9 Swim strategies were characterized as spatial, non-spatial, or thigmotaxic. These broad categories were subdivided into 7 more specific search strategies, 9 and representative strategies are provided (Figure 2). The percentage of runs for each trial type and primary strategy (Spatial/Non-Spatial/Thigmotaxis) was calculated and analyzed. All assessments conducted by a single experimenter blinded to animal treatment group used preset guidelines to categorize a static image of the swim path taken by the animal within the testing pool. Ambiguous runs were assessed by a second experimenter blinded to treatment group and previous categorization.
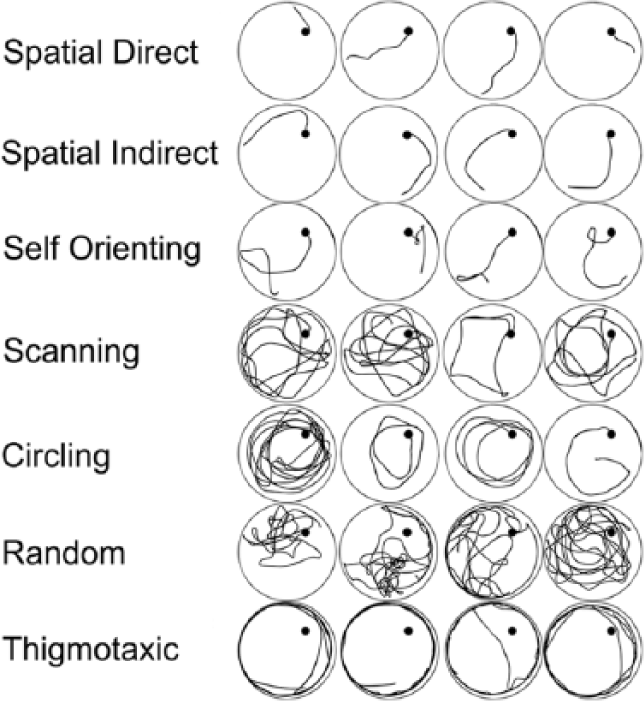
Swim strategies: Example swimming strategies from representative rats evaluated in our cohort. The spatial strategy category includes spatial direct, spatial indirect, and self-orienting. The non-spatial strategy category includes scanning, circling, and random strategies. Thigmotaxic swimming was qualified as its own category, as it represents a reflex response. Each run was classified according to the predominant behavior observed during the course of the trial.
Histological Characterization
Twenty-one days after CCI/sham injury, rats were perfused transcardially under deep anesthesia with pentobarbital (100 mg/kg, intraperitoneally). Brains were extracted, placed in formalin, and paraffin embedded. Seven-micrometer thick coronal sections were cut at 300-µm intervals through the contusion using a rotary microtome, with 1 mm allowed between slice sections. Sections were later deparaffinized in xylenes, rehydrated, and stained with cresyl violet. One coronal section underlying the area of contusion (~3.5 mm posterior to bregma) from each injured rat in all groups was analyzed to determine injury and cognitive training effects on hippocampal CA1, CA3, and dentate gyrus (DG) neuron survival. Morphologically intact neurons (ie, those with a clearly defined cell body and nucleus) were counted using a Nikon Eclipse E600 microscope (40× objective). Data were reported as a percentage of total neurons in ipsilateral (injured) regions relative to each contralateral (uninjured) hippocampal region. 14
To further evaluate tissue loss/sparing, we assessed contusion volume and volume of tissue remaining using MCID Imaging Research Systems (St Catharines, Ontario, Canada). Lesion area (mm2) was calculated by outlining missing cortical tissue for each section taken at 1-mm intervals, and lesion volume (mm3) was determined by multiplying the sum of the contused areas obtained from each section by the distance between sections (1 mm). Area of tissue remaining (mm2) was determined by drawing a midline and measuring the ipsilateral and contralateral hemispheres. Volume of tissue remaining (mm3) was determined by summing remaining tissue areas from each section and multiplying by the distance between sections (1 mm). Percentage of tissue remaining was calculated by dividing the volume of the remaining ipsilateral tissue volume by the remaining contralateral tissue volume.
Statistical Analysis
Statistical analyses were performed using Microsoft Excel and SPSS 20.0. Numerical summaries were computed for all quantitative data. Data are presented as mean ± standard error of the mean (mean ± SEM) and as percentages of trial TA. We examined injury effects, cognitive training status, and the presence/absence of extra-maze cues on platform latencies, peripheral-zone TA, and swim strategy selection. Injury and training status were assessed for motor testing and histology. Based on previous work with non-spatial training prior to injury, 9 we hypothesized that cognitive training effects would result in at least a 20% average reduction in platform latencies and peripheral-zone TA for CCI rats in the cued condition. As we allowed for the possibility of excluding some CogTrained rats due to minimal reductions in peripheral-zone TA on the last cognitive training day, we assumed a minimum group size of 8 rats, to achieve 81% power (95% CI = 0.773-0.844) in detecting the hypothesized difference between groups. Our criterion group sizes were n = 8-11 for CCI rats and Shams.
For quantitative MWM results, a summary table for all main effects can be found in Supplemental Table 2. Quantitative motor and cognitive performance data with repeated measures were assessed using repeated-measures analysis of variance (ANOVA) with Fisher’s least significant difference adjustment for primary pairwise comparisons. Sphericity was evaluated for all repeated measures ANOVA analyses using Mauchly’s W test. When sphericity was violated (W P value <.05), degrees of freedom were adjusted using the Huynh–Feldt estimate, and adjusted P values were used when reporting main effects (see Supplemental Table 2). Group differences in behavioral performance within each trial day were assessed using single-factor ANOVA, with Fisher’s least significant difference adjustment. Hippocampal cell survival and residual tissue volume comparisons for injured CogTrained/No-CogTrained groups were made using Student’s t test. Pearson’s correlations were used to relate histological endpoints with MWM performance across different testing conditions. Generalized estimating equations analyses were used to assess group differences in categorical data generated using a repeated measures study design 19 ; search strategy categorization was assigned for each trial on each testing day.
Results
Postsurgical Reflexes
Postsurgical righting reflexes were similar for CCI/CogTrained rats, compared with CCI/No-CogTrained rats (371.73 ± 3.29 vs 367.75 ± 2.72 seconds; P = .35).
Motor Testing
There was a main effect of group, time, and a significant group × time interaction for beam balance time, beam walk latencies, and beam score (P < .001 all comparisons). There were no significant differences in overall performance across days observed between the Sham/CogTrained and Sham/No-CogTrained on beam balance or beam walk latency/score trials (Supplemental Figure S1). There were no significant differences in overall performance across days for beam balance and beam walk latencies. CCI/CogTrained rats tended to have better beam walking latencies compared with CCI/No-CogTrained rats (P = .058), and latencies to traverse the beam were significantly faster on days 4, 5, and 6 for CCI/ CogTrained rats (P < .05 all comparisons).
Morris Water Maze Testing
Acquisition trials were conducted to establish cognitive training effects on place learning and cue utilization in the context of CCI. A summary of main effects for data collected in the MWM can be found in Supplemental Table 2. Also, swimming speeds on testing D14 were similar between CCI/CogTrained and CCI/no-CogTrained groups (35.43 ± 4.97 vs 34.61 ± 3.80; P = .900) suggesting equivalent motoric abilities during MWM testing.
Cued Condition
There was a significant group and day effect for peripheral-zone TA and latencies (P ≤ .001), and there was a group × time interaction for latencies (P = .016) indicating differences in place learning. CCI/CogTrained rats had shorter latencies than No-CogTrained rats (P = .013) but no difference in peripheral-zone TA, though thigmotaxis was prevalent on D14 for CCI/No-CogTrained rats (Figure 4A). Sham/CogTrained rats performed better across days than Sham/No-CogTrained rats in both latency (P < .001) and peripheral-zone TA (P = .047). While CCI/CogTrained rats did not perform as well as Sham/CogTrained rats in latency (P < .001) or peripheral-zone TA (P = .019), there were no differences versus Sham/No-CogTrained rats, indicating that cognitive training ameliorated CCI induced place learning deficits (Figure 3A and B). CCI/CogTrained rats tended to exhibit less thigmotaxis (P = .064) on postoperative day 14 than Sham/No-CogTrained rats. By postoperative day 18, thigmotaxis was eliminated from all groups. Non-Spatial strategies were most prominent in CCI/CogTrained rats (P < .001) versus Sham/CogTrained rats, and spatial strategies were dominant for Sham/CogTrained, (P < .001) versus CCI/CogTrained groups (Figure 4 A and B), Together, the results suggest that cognitive training reduces thigmotaxis, and improves platform latencies after CCI, primarily through the use of non-spatial strategies learned with cognitive training.
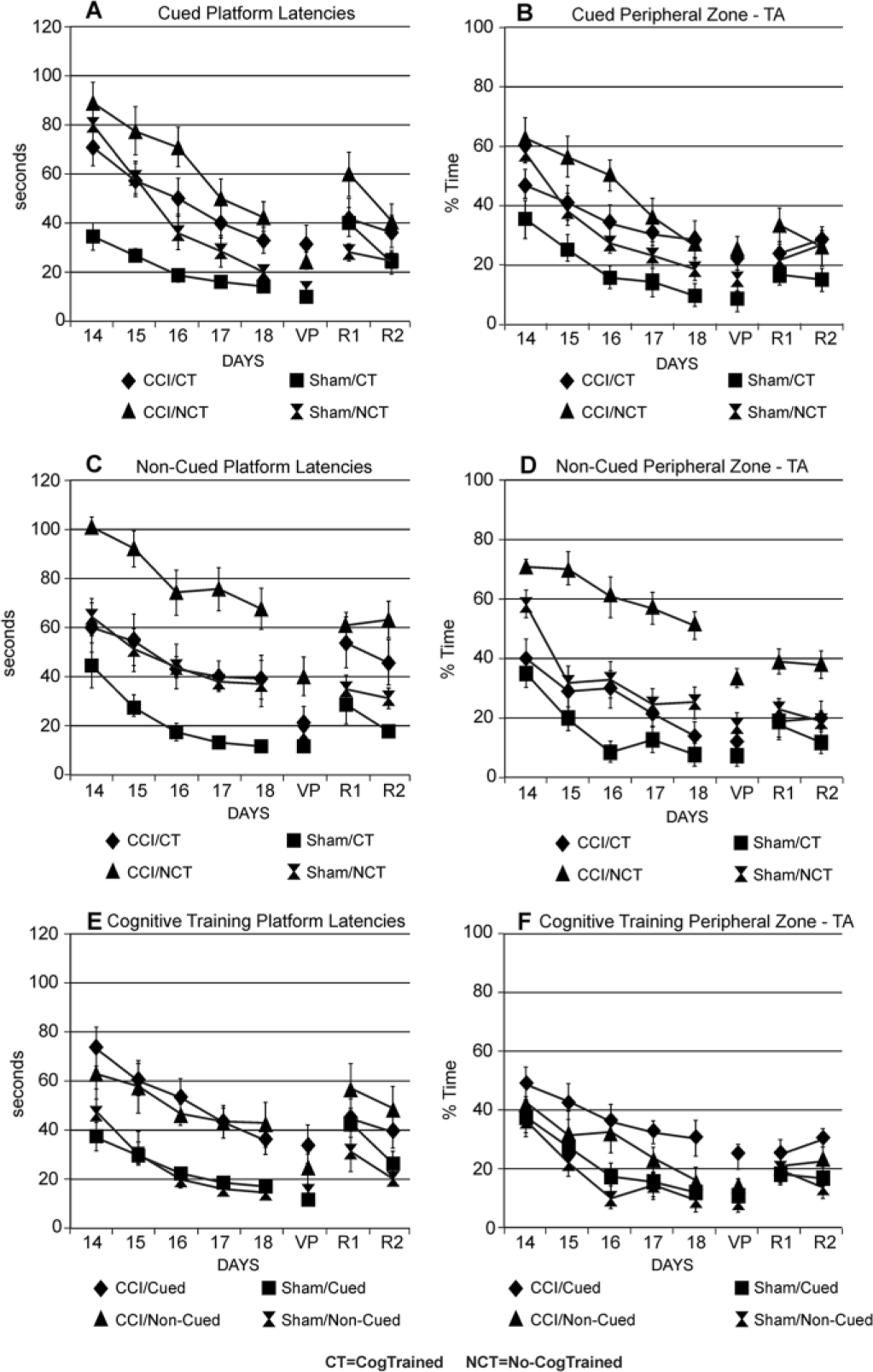
(A) Cued platform latencies—CCI/NCT rats had longer latencies than both Sham groups across acquisition days (CCI/NCT vs Sham/CT, acquisition, P < .001). CCI/CT rats had shorter latencies than NCT rats (P = .013) CCI/NCT versus CCI/CT P < .05 for D15-16 comparisons. CCI/NCT rats had longer latencies than both Sham groups across reversal days (CCI/NCT vs Sham/CT, reversal trial days (CCI/NCT vs Sham/CT-R1 P = .021, R2, P = .018; CCI/No-CT vs Sham/NCT, R1, P = .002, R2, P = .036). (B) Cued peripheral-zone TA—CCI/NCT rats had higher peripheral-zone TA than Sham groups across acquisition trials (CCI/NCT vs Sham/CT, acquisition, P < .001). CCI/CT rats tended to have lower peripheral-zone TA on D14 than Sham NCT rats,. CCI/NCT rats had higher peripheral-zone TA than Sham/CT rats on reversal trial R1 P = .01. (C) Non-Cued platform latencies—CCI/NCT rats had longer latencies than both sham groups across acquisition and VP trials (CCI/NCT vs Sham/CT, P ≤ .001 both comparisons; CCI/NCT vs Sham/NCT, P ≤ .005 both comparisons). CCI/NCT versus Shams P < .01 for daily acquisition (D14-D18) comparisons. CCI/No-CT rats also had longer reversal latencies (CCI/NCT vs Sham/CT, R1 P = .004, R2 P < .001; CCI/NCT vs Sham/NCT, R1 P = .014, R2 P = .001) trial days. (D) Non-Cued peripheral-zone TA—CCI/NCT rats had higher peripheral-zone TA than both sham groups across acquisition trials (CCI/NCT vs Sham/CT P < .001; CCI/NCT vs Sham/NCT, P < .001). CCI/NCT versus Shams P ≤ .001 for daily acquisition (D15-D18) comparisons. CCI/NCT rats also had higher VP peripheral-zone TA (CCI/NCT vs Sham/CT P < .001; CCI/NCT vs Sham/NCT, P = .003), and Reversal Trial peripheral-zone TA (CCI/NCT vs Sham/CT R1 P = .003, R2 P < .001; CCI/NCT vs Sham/NCT, R1 P = .015, R2, P = .003). (E) Cognitive training platform latencies—CCI groups had longer latencies than sham groups (P < .001). Daily acquisition trial comparisons revealed lower latencies for sham groups on D15-D18 (P < .05 all comparisons). There were no differences among CCI groups or among sham groups across acquisition trial days, VP, or reversal trials. (F) Cognitive Training peripheral-zone TA—CCI/CT/Cued rats had significantly higher peripheral-zone TA than shams across acquisition trials (P ≤ .017 both comparisons). Acquisition trial comparisons revealed higher peripheral-zone TA for CCI/CT/Cued rats than either sham group on days 16-17 P ≤ .02, all comparisons). On D18, CCI/CT/Cued rats had higher peripheral-zone TA than CCI/CT/Non-Cued (P = .041) and Shams (P ≤ .012; all comparisons). For VP, CCI/CT/Cued had higher peripheral-zone TA than Shams (CCI/CT/Cued vs Sham/CT/Cued, P = .018; CCI/CT/Cued vs Sham/CT/Non-Cued, P = .016). CCI, controlled cortical impact injury; CT, CogTrained; NCT, No-CogTrained; TA, Time Allocation; VP, Visible Platform.
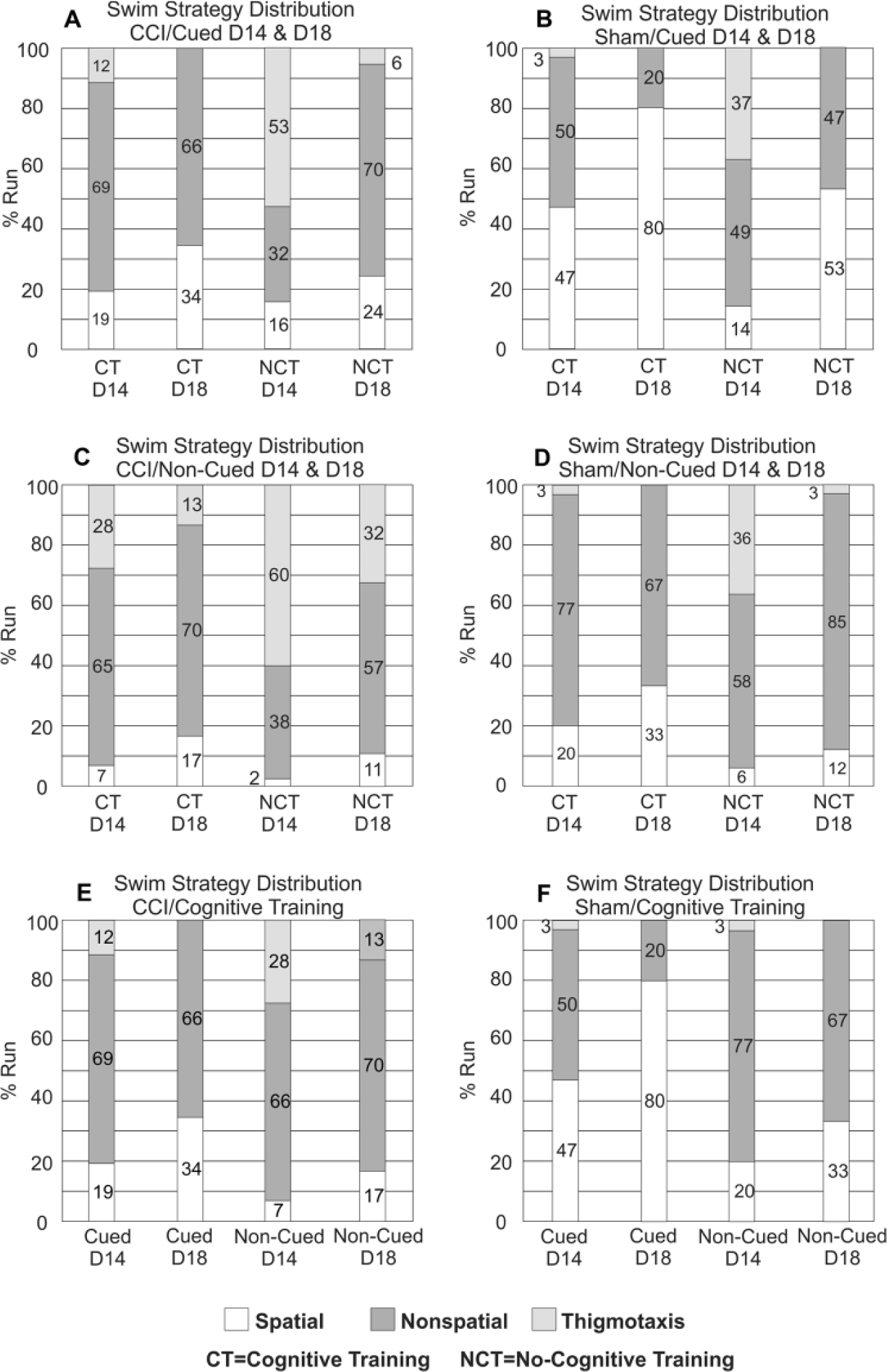
(A) D14 and D18 swim strategy distributions—CCI/Cued rats: CCI/NCT rats exhibited primarily thigmotaxic search strategies on D14, but by D18 non-spatial strategies were most prevalent (P < .001). CCI-CT groups had a nonsignificant increase in spatial strategies by D18 compared with D14 (P = .226). Neither group exhibited a predominantly spatial swim strategy distribution. (B) D14 and D18 swim strategy distributions—Sham/Cued rats: Sham NCT rats had the most thigmotaxis among groups on D14 (P = .005). Thigmotaxis was completely suppressed by D18 for the NCT group. By D18 Sham rats had a higher spatial search strategy frequency compared with their D14 performance (P < .001 both comparisons). (C) D14 and D18 swim strategy distributions—CCI/Non-Cued rats: While non-spatial strategies are most prominent by D18 of testing among non-cued groups, thigmotaxis was more prevalent compared to CCI/Cued counterparts (CCI/CT cued vs non-cued P = .112
Non-Cued Condition
There was a significant group and day effect for latency and peripheral-zone TA (P < .001). CCI/CogTrained rats exhibited lower peripheral-zone TA and shorter platform latencies than CCI/No-CogTrained rats (P < .001). CCI/CogTrained rats had longer platform latencies (P = .005; but not peripheral-zone TA) than Sham/CogTrained rats. Also, CCI/CogTrained rats performed similarly to Sham/No-CogTrained rats in both latency and peripheral-zone TA. Sham/CogTrained rats had shorter latencies (P = .005) and lower peripheral-zone TA (P = .004) than No-CogTrained shams (Figure 3C and D). CCI/CogTrained and Sham/No-CogTrained rats exhibit similar swim strategy distributions on postoperative days 14 and 18, with both groups preferring non-spatial strategies. CogTrained rats exhibited fewer thigotaxic runs on postoperative day 14 of testing than their no-CogTrained controls (P < .014 both comparisons), an effect that was lost by postoperative day 18. All groups used predominantly non-spatial strategies by postoperative day 18 (Figure 4C and D). Sham/CogTrained rats frequently used what appeared to be spatial search strategies on postoperative day 18, which could reflect increased efficiency with pool navigation and result in shorter platform latencies and patterns that appear to be spatial.
CogTrained Condition
There was a group and day effect for platform latencies (P < .001) and peripheral-zone TA (P ≤ .021). Primary differences were due to injury effects for latencies (CCI/Cued vs Sham/Cued, P < .001; CCI/Non-Cued vs Sham/Non-Cued, P < .001) and peripheral-zone TA (CCI/Cued vs Sham/Cued, P = .017), with trends noted for CCI/Non-Cued versus Sham/Non-Cued; P = .097 (Figure 3E and F). There were no differences between injury or sham groups based on cue status. CCI groups favored non-spatial search strategies, despite the presence of distal spatial cues. Both sham groups used primarily non-spatial strategies on postoperative day 14, but by postoperative day 18, spatial strategies dominated for Sham/Cued rats (P = .006), showing increased pliancy and/or increase cue utilization for CogTrained-Shams when provided extra-maze cues (Figure 4E and F).
Visible Platform
We conducted VP trials to assess the use of a proximal cue (VP) and extra-maze cues in place learning. A summary of the main effects on VP performance are provided in Supplemental Table 2. There was a group effect on platform latency (P = .002) and peripheral-zone TA (P = .039) among cued rats. Neither Sham/CogTrained nor CCI/CogTrained rats differed from their No-CogTrained counterparts on any measure. Sham/CogTrained rats had shorter latencies and lower peripheral-zone TA, than both CCI groups (P < .05, all comparisons). Sham/No-CogTrained rats had shorter latencies than CCI/CogTrained rats (P = .005), and tended to have shorter latencies than CCI/No-CogTrained rats (P = .057; Figure 3A and B). These data indicate that CCI rats suppress thigmotaxis, but suggest that CCI rats had difficulty recognizing the VP as a goal.
There was a group effect on platform latency (P = .005) and peripheral-zone TA (P < .001) among Non-Cued rats. In this condition, CCI/CogTrained rats had lower peripheral-zone TA (P < .001) than CCI/No-CogTrained rats. However, platform latencies were similar, regardless of CogTrained status (P = .095; Figure 3C and D). While CCI/No-CogTrained rats performed worse on all measures than Shams, CCI/CogTrained rats did not differ from either Sham group, indicating that cognitive training has some effect on CCI rats VP trial performance. There was a group effect (P = .007) on platform latencies among CogTrained rats. Both CCI/Cued and CCI/Non-Cued rats exhibited longer latencies than Sham counterparts (P < .05). CCI/Cued rats had higher peripheral-zone TA than Sham/Cued rats (P = .018), though there were no differences among non-cued groups. There were no latency or peripheral-zone TA differences between cued/non-cued CCI or sham rats (Figure 3E and F).
Despite no previous exposure to explicit feedback, CCI/CogTrained and both Sham non-cued rats used primarily spatial strategies (P < .045 all comparisons) versus CCI/No-CogTrained/Non-Cued. CCI/No-CogTrai-ned/Non-Cued rats, however, used non-spatial strategies, and tended to have increased thigmotaxis versus other non-cued groups (P < .065, all comparisons). CCI/Cued rats similarly used spatial and non-spatial search strategies during VP trials. Notably, CCI/CogTrained rats showed less spatial strategies versus their respective Sham/Cog-Trained rats (P < .008 both comparisons; Figure 5C), suggesting relative impairments in cue integration for CCI/CogTrained rats despite effective thigmotaxis sup-pression.

(A) VP swim strategy distributions—Cued rats: Sham/CT rats tended to use more spatial search strategies than CCI/CT rats (P = .087). (B) VP swim strategy distributions—Non-Cued rats: CCI-CT and Sham-CT rats use spatial strategies more frequently than NCT Control groups (P < .043 both comparisons). Sham-CT rats had significantly more spatial strategy trials than CCI-CT rats (P = .002). (C) VP swim strategy distributions—CogTrained rats. There were no differences in spatial strategy frequency based on cue status. Sham-CT groups used spatial frequencies more than CCI-CT groups (P < .007 both comparisons). CCI, controlled cortical impact injury; CT, CogTrained; NCT, No-CogTrained; VP, Visible Platform.
Reversal Trials
A summary of the main effects on reversal trial performance are provided in Supplemental Table 2. There was a significant group effect on platform latency on R1 (P = .018) among Cued rats and a significant group effect on peripheral-zone TA among Non-Cued rats on R1 (P = .008) and also R2 (P = .001). There were no differences observed between CCI/CogTrained/Cued rats and any other group. Also, Sham/CogTrained rats had lower peripheral-zone TA than Sham/No-CogTrained rats on R2 (P = .049). CCI/CogTrained rats did not differ from Sham/No-CogTrained rats in either latency or peripheral-zone TA. Sham/CogTrained rats did not differ from Sham/No-CogTrained rats, and only outperformed CCI/CogTrained rats on platform latencies (R1, P = .026; R2, P = .006). There were no differences between CogTrained or Sham groups in latencies or peripheral-zone TA during reversal trials (Figure 3). Though R1 was somewhat similar with spatial strategy use for CCI versus Sham CogTrained/Cued rats, Sham/CogTrained/Cued rats showed more spatial search strategies on R2 than the CCI-cued group, (P < .03) with similar trends noted with reduced spatial strategy use for Sham/No-CogTrained/Cued rats (Figure 6). These results indicate that Sham/CogTrained/Cued rats most effectively learned the novel platform location and incorporated extra maze cues into their search strategy compared with CCI, while CCI groups effectively used non-spatial strategies with limited incorporation of extra-maze cues.
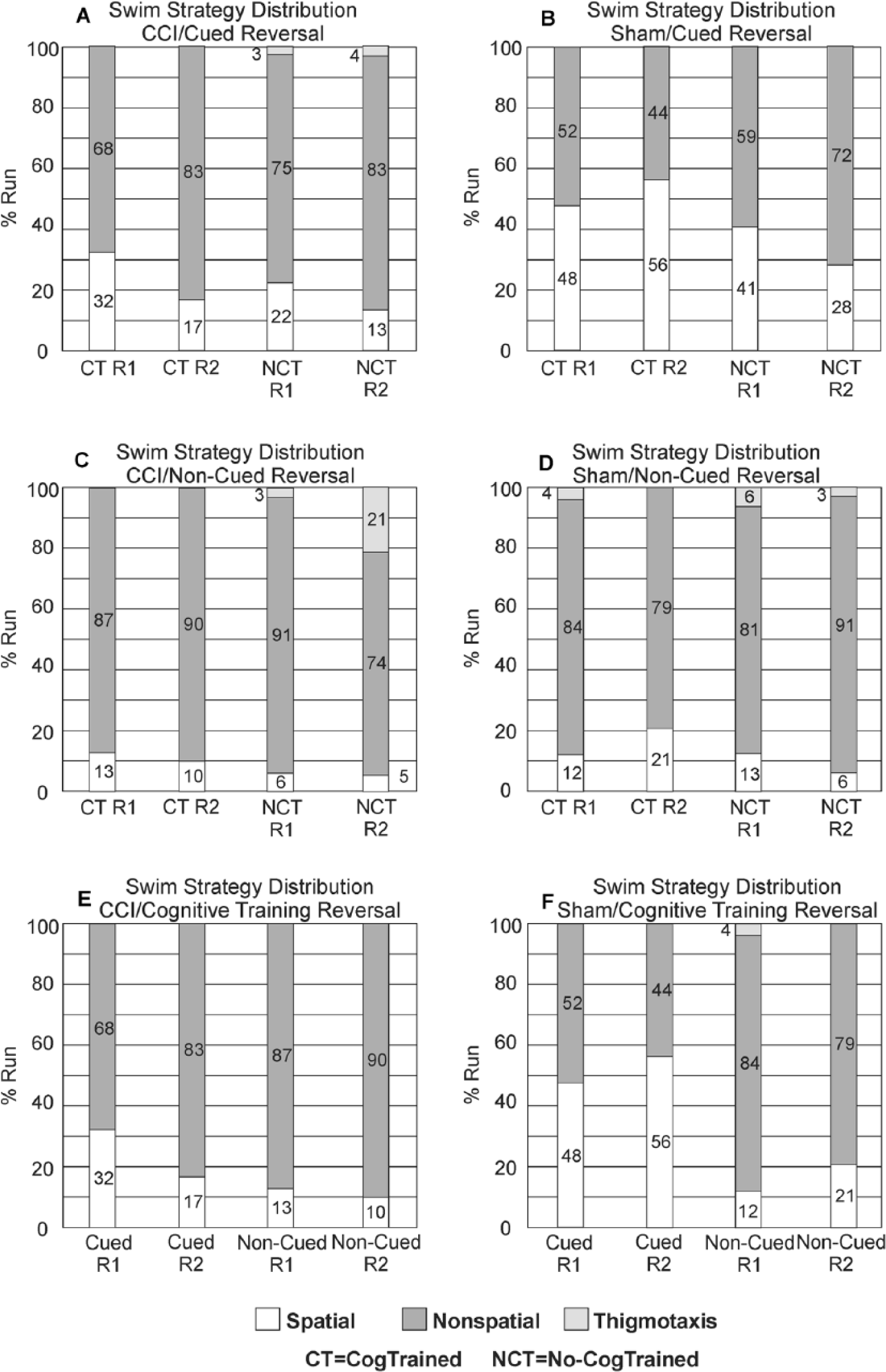
(A) R1 and R2 swim strategy distributions—CCI/Cued rats: There were no significant differences in strategy utilization frequencies noted between groups. (B) R1 and R2 swim strategy distributions—Sham/Cued rats: By R2, there was a difference between the frequency of spatial strategies used for Sham-CT versus Sham NCT (P = .051). (C) R1 and R2 swim strategy distributions—CCI/Non-Cued Rats: CCI/NCT rats exhibit more thigmotaxis on R2 than any other CCI group (P < .05, all comparisons). (D) R1 and R2 swim strategy distributions—Sham/Non-Cued rats: There were no significant differences among sham/non-cued groups in search strategy selection. (E) R1 and R2 swim strategy distributions—CCI/CT rats: No significant differences noted between cued or non-cued groups. Sham/CT use significantly more spatial strategies on R2 than CCI/CT rats (P = .008). (F) R1 and R2 swim strategy distributions—Sham/CT rats: No differences noted between cued or between no-cued groups. CCI, controlled cortical impact injury; CT, CogTrained; NCT, No-CogTrained.
Histology
The mean cortical contusion volume for CCI/CogTrained was 49.60 ± 6.27 mm3, and the mean cortical contusion volume was similar for CCI/No-CogTrained was 54.14 ± 4.24mm3 (n = 9 per group). Similarly, residual tissue analyses showed that the percent of residual tissue remaining for CCI/CogTrained was 79.73% ± 2.23% and CCI/No-CogTrained was 76.28% ±1.41%. All CCI rats had a significant loss of morphologically intact hippocampal neurons in the hemisphere ipsilateral to the injury site compared with the contralateral hemisphere. However, there were no cognitive training effects on regional hippocampal cell survival (CA-1, CCI/CogTrained 20.08% ± 2.40%, CCI No-CogTrained 18.78% ± 1.84%; CA-3, CCI/CogTrained 18.29% ± 3.69%, CCI/No-CogTrained 22.60% ± 4.55%; DG, CCI/CogTrained 52.955% ± 4.13%, CCI/No-CogTrained 48.80% ± 1.26%). Also, CCI groups showed no correlations between latencies or between peripheral-zone TA and contusion volume, percent of ipsilateral tissue remaining or percent hippocampal cell survival.
Discussion
Following TBI, evidence-based cognitive rehabilitation paradigms exist that are tailored to address residual cognitive deficits. 1 However, similar relevant preclinical treatment paradigms do not exist. Based on previous work from this lab with implicit training pre-CCI, 9 we developed a cognitive training model for experimental TBI relevant to clinical cognitive rehabilitation paradigms. The goal for this initial study was to build an effective implicit cognitive training model as an exemplar from which to develop combination therapies, and to test specific learning, memory, and training constructs in relationship to histological and molecular analyses. Our data are novel in that they indicate that postinjury implicit training improved place learning using standard MWM performance metrics such as platform latencies. Cognitive training also helped suppress thigmotaxic behavior across the MWM testing conditions used, particularly within the non-cued condition, as demonstrated with peripheral-zone TA and swim strategy analysis. The data also suggest that implicit learning and memory are relatively intact after CCI and that implicit (non-spatial) training may be harnessed as a therapeutic tool.
While there are some articles published in other injury models20,21 suggesting that training and/or previous acquisition trials can improve platform latencies when later evaluating the same task, we have shown that the MWM can be used to dissociate and evaluate implicit versus explicit learning and memory systems after experimental TBI. 9 The incorporation (vs lack) of extra-maze cues within the context of acquisition trial testing also is novel to experimental TBI behavioral studies and further shows that when rats are tested on the implicitly learned task components, there are minimal impairments. Our data show that the provision of a specific training protocol postinjury can be anchored to implicitly learned task components that are used successfully with and without spatial mapping trials. The novelty of this study compared with our previous work 9 is that postinjury, rats can newly learn the implicit components of the task and apply this training during acquisition trials. As with our previous work with non-spatial pretraining, 9 the 3 group categorization structure (spatial, non-spatial, and thigmotaxis) was sensitive to injury effects and treatment, as well as incorporation of extra-maze and VP as cues to solve the task. Similar to clinical experience with compensatory strategy utilization, the implicitly learned skills acquired may be leveraged in the MWM to improve functional performance (ie, platform latencies), despite modest (nonsignificant) spatial strategy selection improvement corresponding with residual, injury-induced learning impairments.
Controlled cortical impact/CogTrained rats that reached criterion during cognitive training trials performed better than their No-CogTrained counterparts such that they did not differ from Sham/No-CogTrained controls in either the cued or non-cued conditions. These data demonstrate that CCI rats can implicitly learn the non-spatial requirements of the task and apply those skills in novel testing, including situations where explicit (spatial) learning/memory is advantageous. Latency and peripheral-zone TA data indicate that CCI and Sham rats learn the appropriate behavioral strategies during cognitive training. However, swim strategy data demonstrates that CCI/CogTrained/Cued rats exhibit comparable frequencies of non-spatial swimming patterns on postoperative days 14 and 18, indicating that they perseverate on using non-spatial strategy selection, despite the advantage of having distal spatial cues. Strategy perseveration may be due to the immediacy with which testing began after cognitive training and reflect pliancy deficits that limit cognitive flexibility to adjust from preferred strategies initially learned during training. Other work shows that rats with hippocampal damage exhibit pliancy deficits.22,23 Though CCI/No-CogTrained rats do not perform like shams, daily improvements in latency and peripheral-zone TA suggest increasingly efficient non-spatial search strategies. Conversely, Sham/No-CogTrained rats learned the cued task and exhibited a large latency and peripheral-zone TA reduction that matched Sham/CogTrained rat performance during later testing days.
Some CCI rats did exhibit spatial swim strategies in the cued condition. However, these apparent spatial strategies could result from a directional response, instead of precise spatial awareness, as has been demonstrated after hippocampal and fimbria fornix lesions.24,25 Some Shams use apparent spatial strategies in the non-cued condition. These findings may result from efficient non-spatial search strategy utilization that appears to be spatial when graded by a blinded experimenter.
Visible platform results suggest that successful navigation to the novel proximal cue may either be a product of cognitive training, or previous exposure to distal spatial cues. CCI/CogTrained/Cued rats did not differ from Sham/No-CogTrained/Cued rats in VP peripheral-zone TA, but latency differences were observed, indicating that CCI animals used appropriate navigational behavior strategies, but as a group, did not recognize fully the VP as a goal. CCI/CogTrained/Cued rats showed some preference for spatial search strategies (nonsignificant), suggesting that they were able to navigate near the VP, but did not mount the escape platform. In the non-cued condition, cognitive training enabled CCI rats to incorporate an egocentric cue (VP) into searching strategies, in the absence of previous explicit feedback. There were, however, swim strategy differences during the VP trial. Many CCI/CogTrained rats navigated to the platform using spatial strategies and treated the platform as a goal, while CCI/No-CogTrained rats still used predominantly non-spatial strategies or exhibited thigmotaxis, suggesting that CCI rats can use cognitive training to navigate to the novel cue. CCI/CogTrained/no-cued rats did not differ from sham counterparts with VP peripheral-zone TA or latency, while CCI/CogTrained/Cued rats performed worse than their sham counterparts, suggesting that CCI/CogTrained/Cued rats exhibited relative strategy perseverance and decreased performance with the new VP condition.
Few behavioral differences occurred during cued reversal trials. However, CCI/No-CogTrained/Cued rat performance deteriorates, reverting to thigmotaxic behavior, while CCI/CogTrained rats suppressed thigmotaxis. While CCI/CogTrained/Cued and Sham/No-CogTrained/Cued rats behaved similarly in other measures, groups diverged in their swim strategy selection, suggesting that Shams rely on extra-maze cues to guide behavior during reversal trials. CCI rats relied more on the non-spatial behaviors acquired during cognitive training or maze exposure during acquisition trials. Among CogTrained rats there were few quantitative differences based on cue status, indicating that both CCI and Sham/Non-Cued rats developed different, but effective, strategies to solve the MWM task over the different testing conditions.
Spatial mapping theory and localized lesion studies show the hippocampus as central to formulating and retaining spatial maps.10,11,26 However rats with hippocampal injury can learn the spatial version of the MWM with appropriate training23-25,27 suggesting that postinjury, MWM-based cognitive training paradigm can affect functional performance with place learning task conditions. Similarly, clinical studies show patients with TBI perform better with some implicit learning and memory remediation protocols, and they can utilize intact implicit memory to improve explicit functionality,8,28-30 Notably, our data suggest that the implicitly learned information, including search strategy formulation, was applied successfully, particularly for rats in the non-cued condition, across MWM testing conditions, albeit with only modest benefits with explicit learning (ie, extra-maze cue integration) occurring along with reductions in platform latencies where applicable. In this sense, the carryover effects may be considered similar to those observed in clinical studies involving strategy training. 31 However, it is difficult to speculate how implicit training and strategy development in the MWM might affect performance with a very different memory task that also has exploratory task components (eg, radial arm maze). Nonetheless, we believe that the carryover effects of implicitly learned information across MWM conditions are translationally relevant
Anatomical correlates with implicit learning are demonstrated in several dry tasks,32-37 but these relationships have not been fully elucidated in the MWM; also these tasks have not been operationalized from a cognitive training perspective. Our behavioral data suggest that implicit associations intrinsic to striatal systems may be functional in our model of TBI. While the neuroanatomical distinction between implicitly versus explicitly learning is not yet clear with MWM and CCI, these dichotomies may be useful in further developing our experimental cognitive training model using targeted cognitive rehabilitative constructs.
When considering study limitations, a post hoc performance criterion is not representative of clinical rehabilitative practices. Furthermore, No-CogTrained rats may have variable intrinsic abilities with implicit learning that may have resulted in analysis removal had they been exposed to cognitive training. However, the experimental procedures were set a priori, and did not use a training-to-criterion model, in order to allow for exposure and time-specific comparisons across studies and study groups. The training model described here is relatively simple and focuses exclusively on implicit learning and memory. However, this training paradigm provides a clinically relevant construct representing postinjury cognitive training therapies in the CCI model of TBI 15 and also demonstrates the flexibility that rats can use to solve the task, even when platform latencies are similar. Probe trial inclusion in future studies, may provide insight into the dynamic nature of search strategy selection during trials in which the swim time is fixed by assessing how much latencies reflect strategy selection and or a change in strategy selection once it is evident to rats that the platform location has moved. Finally, while No-CogTrained rats were handled and exposed to the warming boxes used with training, additional work assessing water exposure (without swimming) could be informative in discerning an environmental exposure (water) from an implicitly learned task component.
Future work will investigate how other cognitive training concepts within this basic model can further enhance spatial learning and incorporation of extra-maze cues, like priming. In addition to work examining dosing, timing, and duration of cognitive training on place learning, other work might evaluate how well extended acquisition trial training improves performance, relative to cognitive training, to determine the importance of only exposing rats to implicit components of the place learning task prior to beginning acquisition trials. One might consider how rats receiving cognitive training would perform in another unrelated exploratory or memory tasks. Additional work will examine cognitive training effects within different injury models, like blast TBI, which results in significant subcortical/axonal injury. 38 Sex and age differences in cognitive effects also will be explored as will training + drug combination therapies.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by National Institutes of Health Grant 1R21HD071728.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.