
Abstract
Background. Mirrored feedback has potential as a therapeutic intervention to restore hand function after stroke. However, the functional (effective) connectivity of neural networks involved in processing mirrored feedback after stroke is not known. Objective. To determine if regions recruited by mirrored feedback topographically overlap with those involved in control of the paretic hand and to identify the effective connectivity of activated nodes within the mirrored feedback network. Methods. Fifteen patients with chronic stroke performed a finger flexion task with their unaffected hand during event-related functional magnetic resonance imaging (fMRI). Real-time hand kinematics was recorded during fMRI and used to actuate hand models presented in virtual reality (VR). Visual feedback of the unaffected hand motion was manipulated pseudorandomly by either actuating the VR hand corresponding to the moving unaffected side (veridical feedback) or the affected side (mirrored feedback). In 2 control conditions, the VR hands were replaced with moving nonanthropomorphic shapes. Results. Mirrored feedback was associated with significant activation of regions within and outside the ipsilesional sensorimotor cortex, overlapping with areas engaged when patients performed the task with their affected hand. Effective connectivity analysis showed a significantly interconnected ipsilesional somatosensory and motor cortex in the mirrored feedback condition. Conclusions. Mirrored feedback recruits ipsilesional brain areas relevant for control of the affected hand. These data provide a neurophysiological basis by which mirrored feedback may be beneficial as a therapy for restoring function after stroke.
The excitability of the ipsilesional corticospinal system after stroke is a marker of long-term functional recovery, 1 which may explain why therapies that foster activation of the ipsilesional motor cortex have therapeutic value.2,3 We suggest that providing mirrored feedback during movement, by visually mirroring the actual limb movement in real time, can facilitate activity of the ipsilesional motor cortex via rich intrahemispheric4-6 and interhemispheric7-10 connections among frontoparietal areas. Mirror therapy was originally introduced by Ramachandran and Rogers-Ramachandran 11 to reduce phantom limb pain in amputees and may benefit the recovery of hand function after stroke.12-14 The core feature of mirror therapy is the incongruence between actual performance and mirrored feedback, which can be delivered by means of an actual mirror or through virtual reality (VR).
To our knowledge, only one group has so far investigated the neural correlates of mirror therapy in populations with chronic stroke. Michielsen and coworkers 15 showed that mirror therapy may activate the precuneus and posterior cingulate cortex and that 6 weeks of bimanual exercises with mirrored feedback leads to significant increases in the blood oxygen level–dependent signal and its relateralization toward the ipsilesional side (compared to similar training without mirrored feedback). 12 A number of investigations have revealed similar effects in healthy individuals,16,17 along with increases in corticospinal excitability (measured with motor evoked potentials).18-21 Together, these findings suggest that mirrored feedback can modify the activation of sensorimotor brain areas in healthy populations and those with chronic stroke. However, the effective connectivity for mirrored feedback within the sensorimotor network, that is, how nodes in the sensorimotor network functionally interact in response to mirrored feedback, is still unknown. Studies by our group22-24 and others25,26 have shown that feedback of hand motion presented through high-fidelity VR can facilitate classic visuomotor networks in healthy and patient populations. Therefore, the aims of this study in patients with chronic stroke were the following: (1) To identify areas of the visuomotor system where activation is facilitated by mirrored feedback of unaffected hand motion. We hypothesized that mirrored feedback would activate the ipsilesional motor cortex. (2) To identify the functionally interconnected brain networks during mirrored feedback. We hypothesized that regions in the ipsilesional posterior parietal cortex and motor cortex would show increased functional connectivity elicited by mirrored feedback. (3) To test the relationship between changes in mirrored feedback–induced brain activation and the level of hand function. We hypothesized that the degree of activation elicited by mirrored feedback would depend on the level of hand function. (4) To test if regions with activation elicited by mirrored feedback overlap with those that are involved in volitional control of the affected hand. We hypothesized that mirrored feedback would facilitate brain regions that are relevant for control of the affected hand.
For this, patients with chronic stroke underwent event-related functional magnetic resonance imaging (fMRI). Participants performed simple finger movements during scanning, with their finger motion recorded using instrumented data gloves. Kinematic data from the gloves were used to actuate, in real time, the motion of virtual hand models. In any given trial, the VR hand motion corresponded either to the moving unaffected hand (veridical feedback) or to the affected hand (mirrored feedback). The use of a VR mirror setup with simultaneous performance of fMRI and hand kinematics allowed us to (1) ensure that patients focused exclusively on mirrored feedback without being distracted by seeing motion of the other hand, as might occur when using a physical mirror setup; (2) use an event-related design to pseudorandomize the conditions (which would be impossible if using a physical mirror); (3) identify and exclude from analysis any trials confounded by inadvertent motion of the fingers; and (4) deliver a set of control conditions to rule out confounders in activation that may be attributed to motor output, visual field, or arbitrary visual motion effects.
Methods
Participants
Fifteen right-handed patients with hemiparesis due to stroke (5 right hemiplegics and 5 females; mean age, 54 ± 12 years) (Table 1) participated after providing institutionally approved informed consent. One patient had excessive head movement during fMRI, and those data were excluded. Patients’ clinical status was assessed with the Chedoke-McMaster Rating Scale 27 and the Wolf Motor Function Test, 28 including its distal subtest (dWMFT). 29 Lesion location and volume (Figure 1A) were characterized from high-resolution T1-weighted MRI scans using MRIcron software . 30 Inclusion criteria for participation were the following: 30 to 80 years of age, a diagnosis of a first-time stroke at least 6 months before the study date, and normal or corrected-to-normal vision. Exclusion criteria included a history of any neurological pathological abnormality other than stroke, visuospatial deficits such as neglect or visual agnosia, impaired proprioception or sensation, any contraindications to being able to enter an MRI environment, any orthopedic trauma to the hand (not related to the stroke) that would hinder volitional control of movement, and receipt of Botox (Allergan Inc, Irvine, California) injections within the past 3 months.
Patients’ Clinical Information.
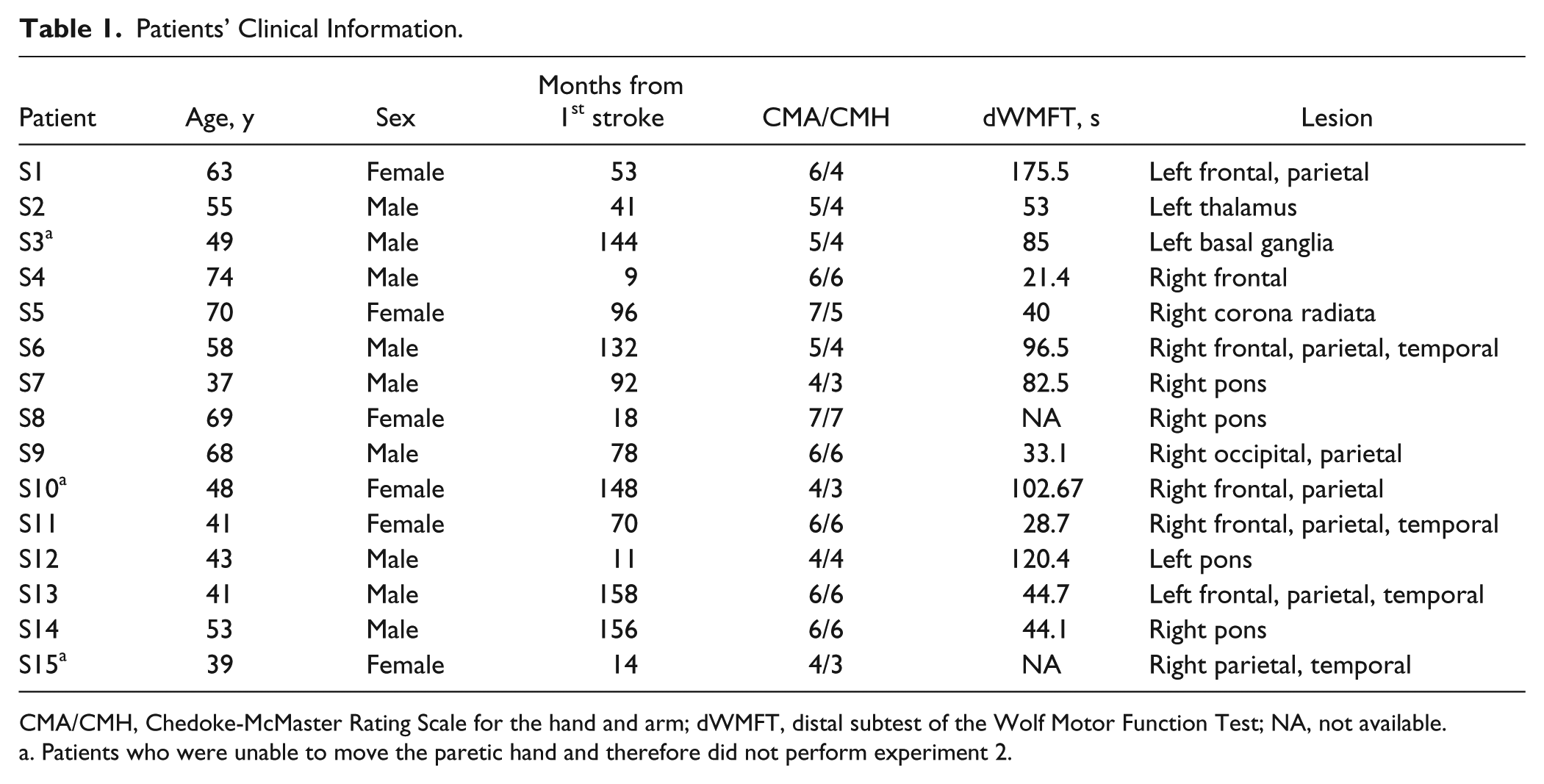
CMA/CMH, Chedoke-McMaster Rating Scale for the hand and arm; dWMFT, distal subtest of the Wolf Motor Function Test; NA, not available.
Patients who were unable to move the paretic hand and therefore did not perform experiment 2.
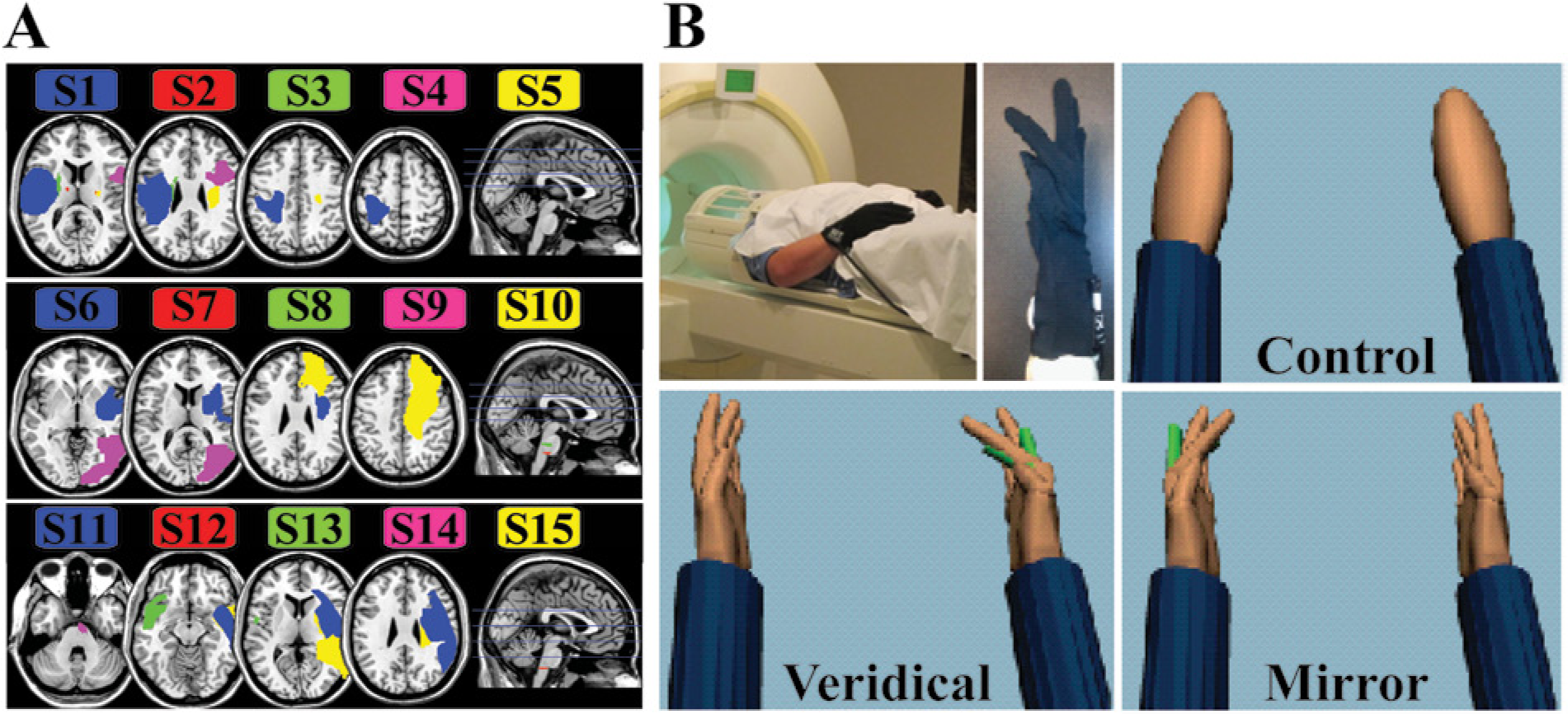
(A) Mapping of each patient’s lesion. Each row shows 5 different patients with lesions filled in with color. Blue horizontal lines in the midsagittal slice mark the levels of the axial slices to the left. (B) The experimental setup and feedback conditions in virtual reality (VR), viewed from the patient’s perspective. Motion of the right unaffected hand (upper left inset) would actuate the right (veridical) or left (mirrored) VR hand or a left/right nonanthropomorphic shape (control). All conditions were randomly interleaved within a functional magnetic resonance imaging run.
Setup
Patients wore left and right MRI-compatible recording gloves (Fifth Dimension Technologies Inc, Pretoria, South Africa).The gloves use fiber-optic sensors to measure metacarpophalangeal and proximal interphalangeal finger joint angles. The gloves were interfaced with a VR environment developed with Virtools software (Dassault Systemes, Vélizy-Villacoublay, France). 31 Patients viewed the VR environment in the scanner through a back-projected image on a semitransparent screen, which enabled them to see a first-person view of left and right virtual hand models in semipronated positions (thumb toward the viewer) (Figure 1). Joint angle data streaming from the recording gloves actuated (in real time) motion of the virtual hand models. For example, motion of the index finger produced a corresponding motion of the virtual index finger. All joint angle data were also saved for statistical analysis of the movement kinematics.
Task
Patients with stroke were required to perform whole-hand finger flexion-extension movements with their unaffected hand (experiment 1) or with their affected hand (experiment 2). During the task, data from the recording gloves were streamed to actuate the virtual hand models. Visual feedback was manipulated as described below. All patients were familiarized with the task and feedback conditions before imaging.
Visual Feedback Conditions for Experiment 1
Experiment 1 was designed to answer aims 1 to 3 (see introductory section). To identify brain activation elicited by mirrored feedback, patients performed the task with the unaffected hand. The event-related fMRI protocol consisted of 4 feedback conditions that were presented in a pseudorandom order (Figure 1). Each condition was repeated for 8 trials per fMRI run. Four fMRI runs were performed per patient. Trial duration was 5 seconds, separated by an intertrial rest period that randomly varied from 4 to 7 seconds. The visual feedback conditions were the following:
Veridical feedback condition: The actuated virtual hand corresponded to the same side as the moving hand (eg, moving the right hand actuated the right virtual hand).
Mirrored feedback condition: The actuated virtual hand was on the opposite side of the moving hand (eg, moving the right hand actuated the left virtual hand).
Control feedback conditions were added to subtract potentially confounding activation related to the position of virtual objects in the visual field, gaze direction, and observed motion. For this, the virtual hand models were replaced with nonanthropomorphic objects (ellipsoids that were similar in size and color to the virtual hands). One of the 2 objects rotated about an oblique axis at 1 Hz.
Veridical control condition: The moving nonanthropomorphic object was on the same side as the moving hand.
Mirrored control condition: The moving nonanthropomorphic object was on the opposite side to the moving hand.
Experiment 2 was used to answer aim 4 (see introductory section). To identify brain activation associated with volitional control of the affected hand, patients moved only the affected hand. Only veridical feedback was presented. Movement and rest periods were presented in alternating order (16 trials per functional run). Two functional imaging runs were conducted. Three patients could not move the fingers of the affected hand consistently and therefore did not perform experiment 2 (Table 1).
Movement Kinematics
Kinematic analysis was used to verify that movements were consistent across the feedback conditions. Trials with inadvertent motion of the stationary hand were not included in the analysis. For each trial, movement onset and offset were defined as the time at which the mean angular velocity of the 4 metacarpophalangeal joints exceeded and then fell below 5% of the mean peak angular velocity. Movement time was the interval between onset and offset. Movement onset and movement time were included in the fMRI general linear model on a trial-by-trial basis for convolution of the blood oxygen level–dependent signal of each event with the hemodynamic response function. To verify that movements remained consistent, the peak angle (angular excursion) and movement time were averaged across trials for each functional imaging run and each feedback condition and were submitted to a 3-way repeated-measures analysis of variance (rmANOVA). Factors (levels) were object viewed (virtual hands, control), feedback type (veridical, mirrored), and fMRI run (run 1, run 2, run 3, run 4). Statistical significance was set at .05 (Bonferroni corrected).
fMRI Acquisition and Analysis
The fMRI data were acquired using a 3-T Siemens Allegra (Munich, Germany) head-only scanner with a standard head coil. Parameters for the high-resolution structural images were the following: repetition time (TR), 2 seconds; echo time (TE), 4.38 milliseconds; voxel size, 0.938 × 0.938 × 1 mm; number of slices, 176; and slice thickness, 1 mm. Parameters for the functional images were the following: T2*-weighted echo planar imaging sequence; TR, 2 seconds; TE, 30 milliseconds; field of view, 100 mm; voxel size, 3 mm3; number of slices, 32; interslice time, 62 milliseconds; and number of volumes, 175. All fMRI data were preprocessed and analyzed in SPM8, (
Functional volumes were realigned to the first volume, slice time corrected, coregistered to the structural image, and normalized to a standardized brain in the Montreal Neurological Institute (MNI) space. A binary mask of each patient’s lesion was used to optimize brain normalization with the DARTEL 32 toolbox in SPM8, consistent with established approaches.32-36 Functional scans were smoothed using an 8-mm full width at half maximum Gaussian kernel. Separate general linear models were created in SPM8 for each experiment and patient.
Analysis was conducted at the single-patient (fixed-effects) and group level (random-effects). For group-level analysis, fMRI scans of patients with left hemisphere lesions were flipped to the right. This is consistent with previous approaches for creating group-level means in patient populations who have lesions in different hemispheres. 37 Statistical significance for all analyses was set at P < .05 (cluster-level family-wise error corrected [voxel extent k > 10]).
The following contrasts were created for experiment 1:
Contrast 1: Main effect of VR hand feedback. This contrast identified regions recruited by observing virtual hand motion versus nonanthropomorphic control (CTRL) objects.
(HAND[mirror] + HAND[veridical]) > (CTRL[mirror] + CTRL[veridical])
Contrast 2: Effect of mirrored feedback. This contrast identified regions recruited by mirrored feedback.
HAND[mirror] > (HAND[veridical] + CTRL[mirror] + CTRL[veridical])
The following contrasts were created for experiment 2:
Contrast 3: Effect of paretic hand motion. This contrast identified regions recruited for controlling movement of the affected hand.
HAND[veridical] > rest
Contrast 4: Conjunction analysis. This contrast identified regions that were recruited both by mirrored feedback (contrast 2) and by volitional control of the paretic hand (contrast 3). For the conjunction analysis, a binary mask of contrast 3 images was created (threshold of t > 1.7) and multiplied by contrast 2 images at a patient-by-patient level. To reduce the possibility of type II error in this sample size, a nonparametric pseudo–t test was computed for the overlap images using SnPM. 38 The SPM8 parameters were set to zero variance smoothing and the maximum number of iterations. Results were evaluated at a cluster-wise threshold of P < .05 (family-wise error corrected).
Effective Connectivity
Effective connectivity explains the task-specific changes in functional connectivity between a region of interest and the rest of the brain. Effective connectivity was quantified using the generalized psychophysiological interaction (PPI) toolbox for SPM8.39-41 Three a priori–defined regions of interest within the ipsilesional sensorimotor cortex42,43 were specified, based on their importance in sensorimotor control, for their predictive validity of stroke recovery 44 and their involvement in mirrored feedback paradigms in healthy populations (see introductory section) 45 and in contrast 2 of the current study. A fourth adjacent seed (supplementary motor area [SMA]) was added based on published reports that it has significantly altered connectivity with the motor cortex in populations with stroke. 46 The mean ± standard deviation of each seed location in the MNI space was the following:
Precentral gyrus corresponding to the motor cortex (Brodmann area 4 [BA4]): 40 ± 4.2, −13 ± 4.5, 47 ± 2.1
Primary somatosensory cortex (BA1): 62 ± 1.7, −18 ± 1.4, 40 ± 5.3
Secondary somatosensory cortex (BA2): 41 ± 7, −30 ± 4, 50 ± 3
SMA: 0 ± 8, −4 ± 16, 67 ± 9
The procedure for PPI analysis is well published.40,41 Briefly, the time series of each seed is extracted, deconvolved with the boxcar function of the task events for each condition, and entered as a regressor in the general linear model. The strength of the regression between the seed and the remaining voxels in the brain represents the effective connectivity map for that seed (referred to as a contribution map). Contribution maps were generated for each condition and contrasted at the single-patient level using the following contrast:
Contrast 5
HAND[mirror] > (HAND[veridical] + CTRL[mirror] + CTRL[veridical])
These contrast images were statistically tested at the group level using a 1-sample t test.
Relationship Between Brain Activation and Hand Function
To ascertain if the magnitude of the mirrored feedback–based activation was related to the degree of hand function, a regression analysis was performed on a patient-by-patient basis between the t values of the significantly activated clusters of contrast 2 and contrast 5 and the dWMFT clinical score.
Results
Consistency of Hand Kinematics Across Conditions
The mirror experiment was performed with the unaffected hand. Hand kinematics was inspected to identify missed trials and inadvertent movements. Such trials comprised only 2% of all trials and were excluded from analyses. A χ2 test revealed no significant difference in the number of trials removed from each condition (P > .05).
Hand kinematics was also analyzed to verify if movements were consistent across conditions. A 3-way rmANOVA for mean angular velocity and movement duration did not show significant effects of object viewed (virtual hands, CTRL), feedback type (veridical, mirrored), or fMRI run (run 1, run 2, run 3, run 4) (P > .05). Finger excursion (movement amplitude) tended to increase from 0.7 ± 0.39 rad in the first run to 0.89 ± 0.41 rad in the last run of the CTRL condition (fMRI run × object viewed interaction: F3,39 = 4.67; P = .01), although this did not occur when patients performed the task with real-time feedback of the VR hands. These analyses suggested that patients performed slightly larger amplitude movements in the control condition but that, otherwise, movements were generally comparable across veridical and mirrored feedback conditions, making any differences in subsequent fMRI results unlikely to be confounded by movement kinematics.
Main Effect of Movement on Brain Activation
Motion of the unaffected hand (across all feedback types) was associated with activation in a distributed network predominantly involving the contralateral frontoparietal network that is traditionally recruited during visually guided movement. Motion of the affected hand was associated with significant activation of the bilateral sensorimotor cortex, parietal lobe, ipsilateral occipitotemporal cortex, and contralateral insula (Table 2).
fMRI Results for All Contrasts.
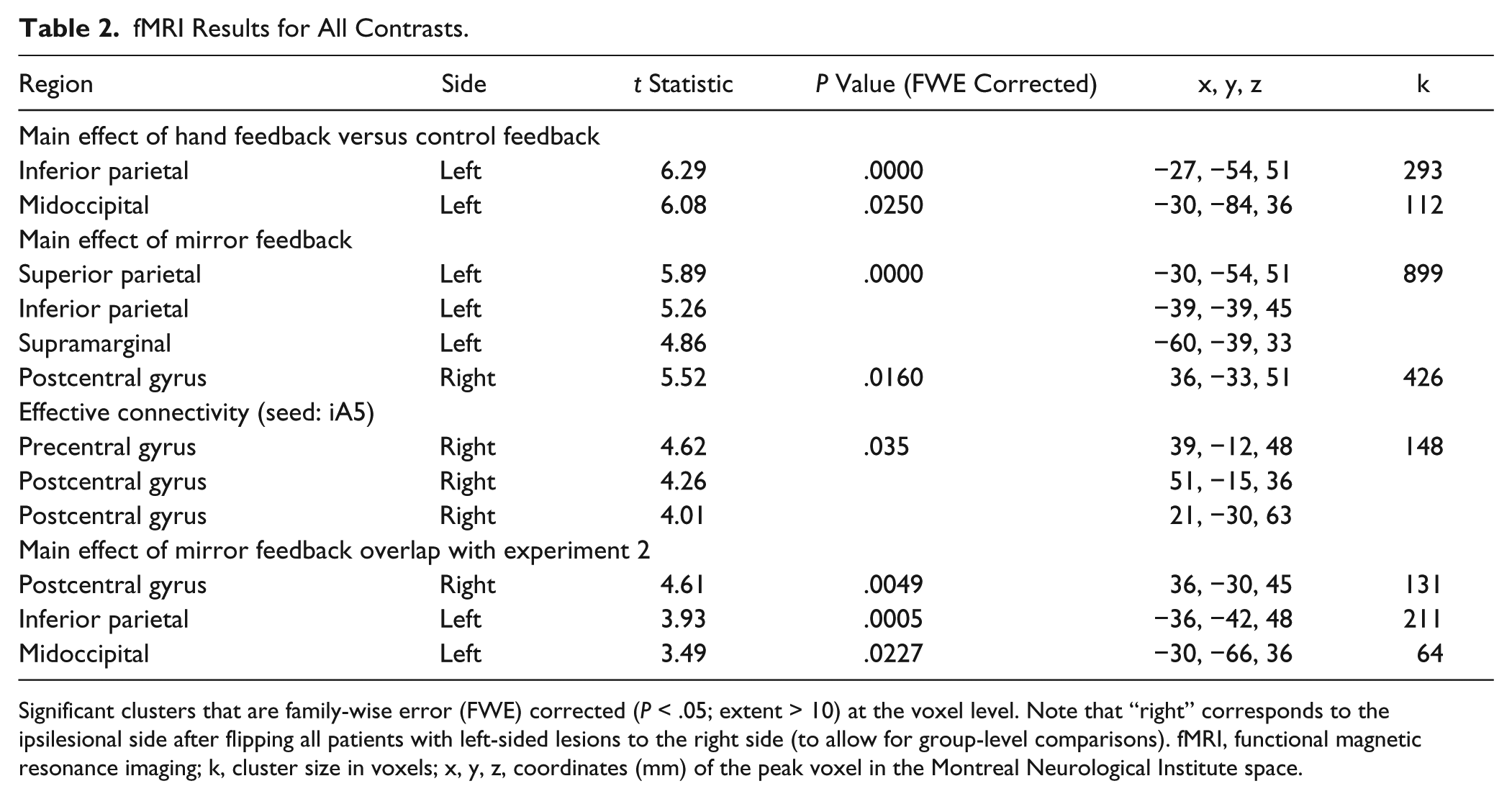
Significant clusters that are family-wise error (FWE) corrected (P < .05; extent > 10) at the voxel level. Note that “right” corresponds to the ipsilesional side after flipping all patients with left-sided lesions to the right side (to allow for group-level comparisons). fMRI, functional magnetic resonance imaging; k, cluster size in voxels; x, y, z, coordinates (mm) of the peak voxel in the Montreal Neurological Institute space.
Main Effect of Viewed Object Type on Brain Activation
The type of object viewed (virtual hands > CTRL) was associated with significant activation in the contralesional posterior parietal cortex, extending to the occipitoparietal area, middle occipital cortex, and ipsilesional somatosensory cortex (Table 2).
Brain Activation Related to Mirrored Feedback
Mirrored feedback was associated with significant activation of a large cluster in the ipsilesional postcentral gyrus, corresponding to BA1 (Figure 2 and Table 2). The cluster extended rostrally to the anterior bank of the central sulcus (BA4) (eg, primary motor cortex) and caudally to the precuneus, adjacent to a region identified by Michielsen et al 15 in their mirror paradigm. Figure 2 (also Table 2) shows that mirrored feedback also elicited significant activation in the contralesional cortex, including the postcentral gyrus (BA1-BA3), superior bank of the intraparietal sulcus and precuneus (BA7), and supramarginal gyrus (BA40). A conjunction analysis (Figure 3) revealed that activation for mirrored feedback (during motion of the unaffected hand) was topographically similar to regions activated by volitional motion of the affected hand under veridical feedback conditions. The overlapping regions included the ipsilesional postcentral gyrus (corresponding to the junction of BA3 and BA5) and contralesional superior parietal lobule (BA7) (Table 2).
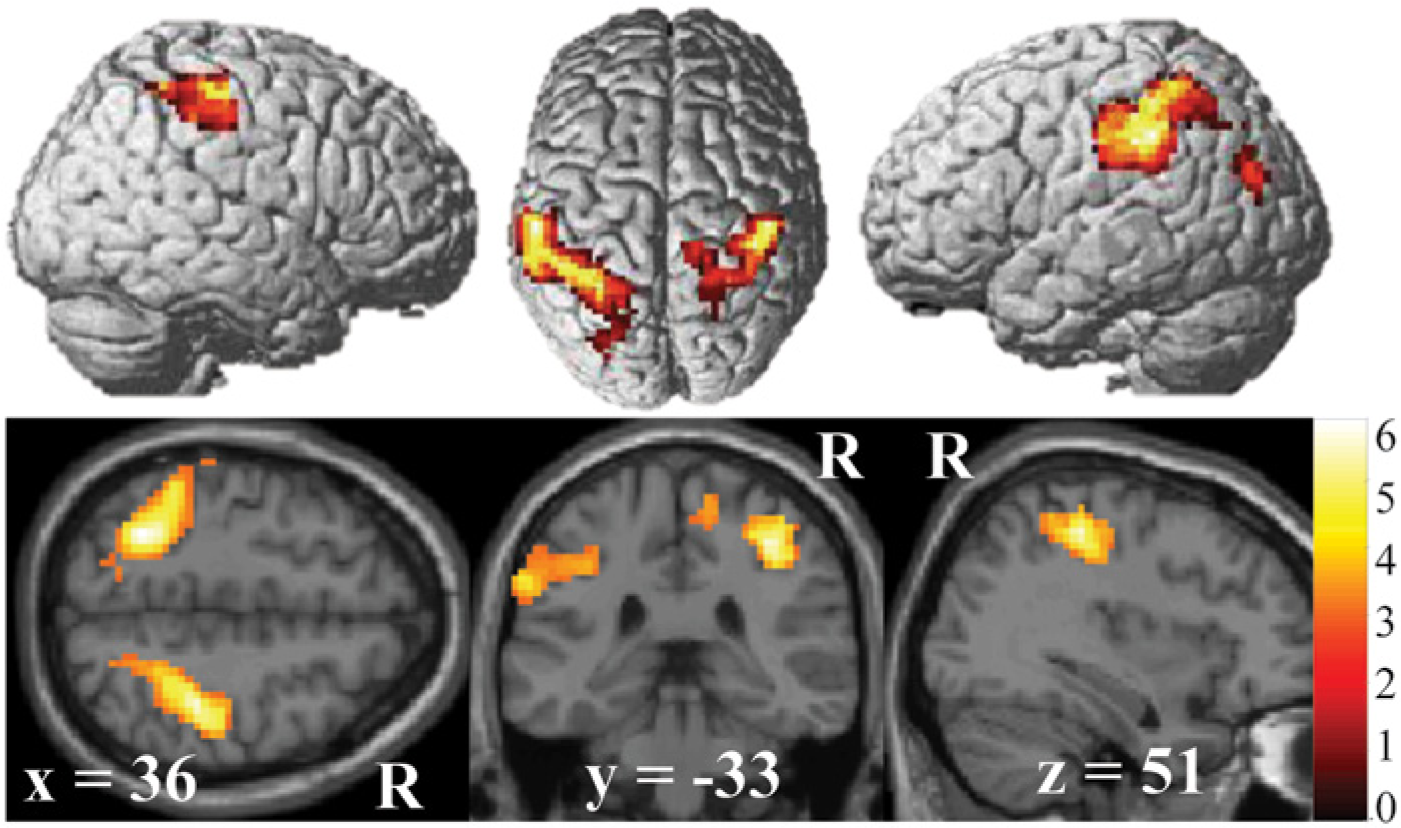
Group mean of the main effect of mirrored feedback (contrast 2) (family-wise error corrected P < .05; minimal cluster size k = 10). For averaging purposes, left-sided lesions were flipped to the right (see Methods) such that the right hemisphere (R) represents the ipsilesional hemisphere. Surface plots were generated using render and canonical templates in SPM8. Note that mirrored feedback activation was observed in the ipsilesional precentral gyrus, corresponding to the hand area of the motor cortex, along with other distributed frontal and parietal areas.
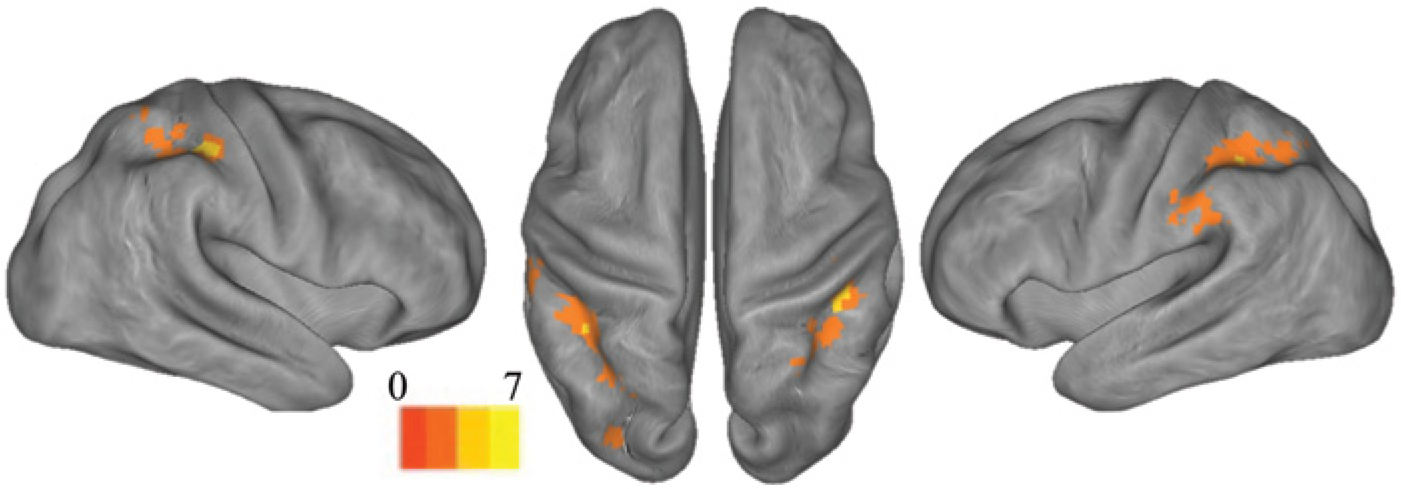
Group mean of the conjunction analysis between contrast 2 (experiment 1) and contrast 3 (experiment 2) to identify topographic overlap between brain regions recruited during motion of the unaffected hand with mirrored feedback and those controlling motion of the affected hand.
Effective Connectivity During Mirrored Feedback
Effective connectivity was used to quantify task-based network interactions between 4 seed regions (BA4, BA1, BA2, SMA) and the rest of the brain (see Methods: Effective Connectivity for details). Figure 4A and 4B show that the connectivity between the BA1 seed and the primary motor cortex (BA4) and between the BA1 seed and the primary sensory cortex (BA3) were significantly stronger for mirrored feedback, relative to the other conditions. Effective connectivity between the other seed regions and the whole brain did not differ between conditions.
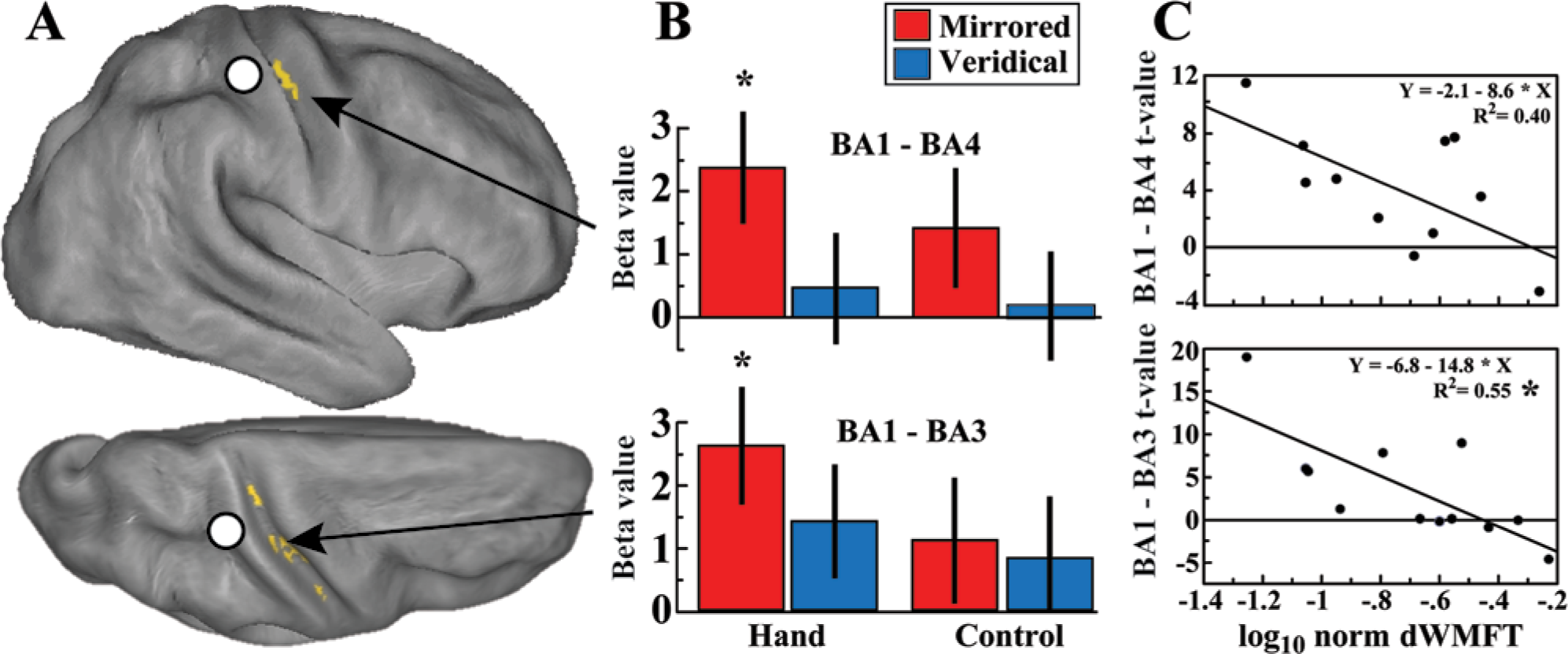
(A) Effective connectivity of the psychophysiological interaction between the Brodmann area (BA1) seed region (white circle) (Montreal Neurological Institute space: 63, −18, 39) and the rest of the brain (family-wise error corrected P < .05; minimal cluster size k = 10) overlaid on a partially inflated cortical surface (Caret software, http://www.nitrc.org/projects/caret/, St. Louis, MO) from the right lateral and top view. (B) Bar plots showing the strength of effective connectivity between the BA1 seed and BA4 (39, −12, 48) and between the BA1 seed and BA3 (51, −15, 36) in each of the 4 conditions. (C) Linear regression between the t values corresponding to the strength of effective connectivity (y-axis) and level of hand function (x-axis). Hand function is plotted as the log-normed distal subtest of the Wolf Motor Function Test (dWMFT) score (1-log10 dWMFT), with larger values indicating better hand function.
Relationship Between Brain Activity and Affected Hand Function
No significant correlation was noted between the level of hand function (log-normed dWMFT) and the degree of activation elicited by mirrored feedback, relative to the other conditions. However, Figure 4C (bottom) demonstrates that a significant correlation between BA1-BA3 effective connectivity and the log-normed dWMFT score (F1,11 = 14.14; P = .0037, Bonferroni corrected) was present, suggesting that stronger connectivity was elicited in persons with greater impairment. The correlation between BA1-BA4 effective connectivity and the dWMFT score (Figure 4C, top) showed a similar trend but did not reach statistical significance after correction (F1,10 = 5.94; P = .0375).
Discussion
Our data show that mirrored feedback during movement of the unaffected hand is associated with significant activation of the ipsilesional sensorimotor cortex. The observation of virtual hands versus nonanthropomorphic shapes recruited higher order visual networks in the lateral occipital cortex similar to other fMRI studies.22,26 The fMRI contrast, with subtraction of the control conditions, as well as the analysis of finger kinematics acquired concomitantly with the fMRI data, suggests that this activation is unlikely to be confounded by visual motion, gaze effects, 47 location of objects in a particular hemifield, any potential differences in movement kinematics between conditions, or inadvertent mirror movements. 48 Finally, the presence of significant activation even after subtraction of activation in the control conditions (in which patients performed the same movements) suggests that the cortical response was most likely elicited by feedback processes rather efferent signals from the contralesional motor cortex.
Previous work from independent laboratories suggests that the sensorimotor system of the hemisphere contralateral to the mirrored feedback hand can be facilitated in healthy individuals. While this facilitation seems to include the corticospinal system itself,18,20,49,50 work on fMRI, electroencephalography, and magnetoencephalography also suggests that it may extend to higher order visuomotor areas in the parietal, posterior temporal, and occipital areas as well.15-17,45,51 Further substantiating this view is a recent elegant study in which healthy individuals trained on fine motor tasks over 4 days using their right hand with either veridical or mirrored feedback. 52 The mirrored feedback group demonstrated superior performance on the task when tested with the untrained left hand and an increase in activity in the right dorsal premotor cortex, left ventral premotor cortex, and left SMA. Together, these data in healthy individuals lend support to the idea that the mirrored feedback responses observed in our study may be similar, although not identical, to those noted in the above-mentioned studies.
Given the therapeutic potential of mirror therapy, the question therefore arises whether the brains of patients with stroke have the capacity to respond to mirrored feedback similar to what has been noted in healthy individuals. Interestingly, studies investigating the effectiveness of mirror therapy have yielded inconsistent results12,13,53-57 (for review, see Thieme et al 14 and Ezendam et al 58 ), suggesting that the responsiveness of the lesioned nervous system, particularly the sensorimotor system, may be questionable. Indeed, the only study, to date, that we are aware of testing for activation associated with mirrored feedback in patients with stroke found significant activation induced by mirrored feedback in the posterior cingulate cortex and precuneus but not the sensorimotor cortex. 15 Differences between our design and those of Michielsen et al 15 (such as using a blocked vs event-related design and a real vs virtual mirror setup) may have led to the distinctions in the observed results. Indeed, some have shown that mirrored feedback presented through VR may potentiate corticospinal excitability more strongly than when presented through a real mirror, 25 although the reason for this still remains unclear. In this regard, a limitation of our design was the absence of a comparison group with a real mirror, which was logistically impossible in an event-related fMRI design, precluding us from drawing such comparisons.
Effective Connectivity of Networks Processing Mirrored Feedback
A central aim of this study was to characterize the effective connectivity of the network interactions invoked by mirrored feedback. Our data show significant effective connectivity interactions between the ipsilesional somatosensory cortex (BA1) and the sensorimotor cortex in the caudal and rostral banks of the central sulcus (BA3 and BA4). Notably, we found that BA1-BA3 effective connectivity was correlated with the level of hand function, suggesting a testable prediction that mirror therapy may have the greatest yield for more impaired individuals, at least for individuals in the chronic stage. If true, then this may provide a basis for testing the potential of mirror therapy in patients with more acute stroke who are generally more impaired and have the greatest potential for recovery. It is interesting to note that no change in effective connectivity was observed between an adjacent region, the SMA, and the sensorimotor cortex, highlighting the specificity of mirrored feedback–induced interactions. However, these speculations need to be directly tested in empirical studies. Moreover, it remains unclear at this point how lesions to nodes in this network may affect the mirrored feedback–induced changes in activation.
Mirrored Feedback Recruits Regions Involved in Volitional Control of the Affected Hand
A notable finding in our study was that regions activated by mirrored feedback share a degree of topographical overlap with regions involved in volitional control of the affected hand. These data suggest that mirrored feedback may facilitate motorically relevant regions of the sensorimotor system in patients with stroke with moderate hemiparesis. Because movement-based interventions for lower functioning individuals are limited by the amount of remaining volitional motion after stroke, activation of the sensorimotor system through mirrored feedback may be particularly suitable for severely hemiplegic patients or for those in the early stages after stroke. This finding is consistent with recent fMRI work in amputee patients, showing that mirrored feedback paired with mental imagery of moving the amputated limb can recruit sensorimotor territories that would represent the amputated extremity. 59
Conclusion
Our study demonstrates that mirrored feedback can invoke activity and strengthen functional interactions within topographically relevant sensorimotor areas of the lesioned hemisphere. Further study of the neural mechanisms and causal interactions that underlie mirrored feedback is necessary to develop therapeutic interventions that boost activity in select brain networks to potentially enhance brain reorganization.
Footnotes
Acknowledgements
The authors thank Michal Kafri and Thushini Manuweera for review and feedback on this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by National Institutes of Health grants K01-HD059983 (E.T.) and R01-HD58301 (S.V.A.) and by National Institute on Disability and Rehabilitation (Research Rehabilitation Engineering Research Center) grant H133E050011 (S.V.A.).