
Abstract
Background. During recovery from a unilateral cortical stroke, spared cortical motor areas in the contralateral (intact) cerebral cortex are recruited. Preclinical studies have demonstrated that compensation with the less-impaired limb may have a detrimental inhibitory effect on the intact cortical hemisphere and could impede recovery of the more-impaired limb. However, evidence from detailed neurophysiological mapping studies in animal models is lacking. Objectives. The present study examines neurophysiological changes in the intact hemisphere of the rat following a unilateral ischemic infarct to cortical forelimb motor areas. Methods. A total of 8 rats were trained for 2 weeks on a reach and retrieval task prior to an ischemic infarct induced by the vasoconstrictor endothelin-1 injected into the cortical gray matter encompassing the 2 forelimb motor representations: the caudal forelimb area (CFA) and the rostral forelimb area (RFA). Animals were randomly assigned to an infarct/training group (n = 4) or an infarct/no-training group (ie, spontaneous recovery, n = 4). After a 5-week postinfarct period, motor areas of the intact hemisphere (CFA and RFA) were characterized using intracortical microstimulation techniques. The resulting maps of evoked movements were compared with maps derived from CFA and RFA in normal rats (normal, n = 5; normal/training, n = 4). Results. Compared with the normal/no-training group, CFA representations were significantly smaller in the infarct/training group but not in the infarct/no-training group. No significant differences were found in RFA. Conclusions. Repetitive training of the more-impaired forelimb during the postinfarct recovery period reduces the size of motor representations in the intact hemisphere.
Following a unilateral stroke, spared cortical motor areas of the intact cerebral cortex in the opposite hemisphere are recruited during movement of the more impaired limb. 1 In considering the mechanisms underlying such reorganization, preclinical rodent models of focal ischemic injury have found a significant increase in dendritic arborization, microtubule associated protein 2, N-methyl-D-aspartate subunit 1, and FOS 2 as well as crossed corticostriatal sprouting. 3 In the present study in adult rats, intracortical microstimulation (ICMS) techniques were used to assess functional representations of the forelimb within the intact motor cortex of the rat opposite an experimental ischemic infarct that included the rodent homolog of the primary motor cortex forelimb area (caudal forelimb area [CFA]) and premotor cortex (rostral forelimb area [RFA]). The results indicate that daily postinfarct training on a reach and retrieval task with the more impaired forelimb significantly reduces the overall size of the CFA but not the RFA in the intact, contralateral hemisphere.
A total of 17 adult Long Evans rats (weight, 470 g; age, 5-6 months) were randomly assigned to 1 of 4 groups prior to behavioral assessment on a reaching task: infarct group with no postinfarct training (infarct/no-training, n = 4); infarct group with 4 weeks postinfarct training, beginning 10 days after the infarct (infarct/training, n = 4); normal, naïve group (normal, n = 5); and normal group with 2 weeks training (normal/training, n = 4). The 2 normal groups were included as control groups for possible lesion and training effects. Because in this infarct model rats cannot consistently reach out of the chamber during the first 2 weeks postinfarct, the 2-week normal/training group was included to match the experiences of the postinfarct trained rats. Each rat was singly housed within a temperature-controlled vivarium on a 12:12 hour light-dark cycle. Food and water were provided ad libitum prior to behavioral training. During training, rats were placed on a feeding schedule while receiving ad libitum access to water. All procedures were in accordance with the NIH Guide for the Care and Use of Laboratory Animals and the Animal Care and Use Committee of the Kansas University Medical Center.
Ischemic infarcts were made within the CFA and RFA contralateral to the rat’s preferred forelimb for reaching by multiple intracortical injections of endothelin-1 (ET-1; 0.3 µg ET-1 dissolved in 1 µL sterile saline [~120 pmol]; Bachem Americas, USA), a potent vasoconstrictor. 2 Boreholes were made through the skull and ET-1 injected at 8 locations relative to bregma: A/P, M/L (2.5, 2.5) (2.5, 3.5) (1.5, 2.5) (1.5, 3.5) (0.5, 2.5) (0.5, 3.5) (−0.5, 2.5) (−0.5, 3.5). At each location, 0.33 µL ET-1 (3 nL/s) was injected at a depth of ~1.5 mm below the pial surface via a micropipette (160 µm outer diameter) attached to a 1-µL Hamilton syringe.
The adequacy of the ET-1 lesion on skilled reaching performance with the forelimb contralateral to the lesion was assessed with a repeated-measures analysis of variance (ANOVA) comparing performance 1 and 2 weeks prior to and 1 week after the infarct. There was no significant group effect F (1,6) = 0.75 (P = .40) nor was there a significant interaction F (2,12) = 0.96 (P = .41), indicating that there were no group differences before or after the infarct. A significant effect of time F (2,12) = 52.40 (P = .0001) indicates that the forelimb performance declined in both groups after the infarct. Both infarct groups showed limited recovery over the 5 weeks postinfarct, and there was no significant difference in performance between the infarct/training and infarct/no-training groups: F(1, 6) = 0.49; P = .49).
Standard ICMS procedures 4 were used to derive motor maps within the CFA and RFA ipsilateral to the preferred forelimb in all groups (ie, intact hemisphere in infarct groups). Analysis of ICMS data was performed by an ANOVA followed by Dunnett’s post hoc comparisons when appropriate. The mean stimulus threshold was similar for all 4 groups in both the CFA—F(3, 13) = 0.90, P = .47—and RFA—F(3, 13) = 0.35, P = .79. There was a significant difference in size of the CFA between groups: F (3,13) = 4.75; P = .02. Compared with the normal group, total CFA (distal + proximal representations) within the intact hemisphere was significantly smaller in the infarct/training group (P = .015) but not in the infarct/no-training group; P = .47). There was no difference between the normal and normal/training groups (P = .99). Although the RFA was also smaller in the infarct/training group, this difference was not significant between groups: F(3, 13) = 1.95; P = .17. When the distal and proximal representations were analyzed separately, neither distal nor proximal forelimb representations in the CFA or RFA were significantly different between groups even though there was a trend for distal CFA and RFA in the infarct/training group to be smaller: distal CFA, F(3, 13) = 2.06, P = .16; distal RFA, F(3, 13) = 1.94, P = .17; proximal CFA, F(3, 13) = 1.17, P = .36; proximal RFA, F(3, 13) = 0.55, P = .66.
Previous studies have shown that use of the less-impaired limb has an exaggerated, inhibitory influence over the damaged hemisphere after a unilateral lesion in the motor cortex of the rat.5,6 The present results suggest that use of the more-impaired limb may also have an inhibitory influence over the intact hemisphere. In a recent review of clinical transcranial magnetic stimulation (TMS) studies, Corti et al 7 reported several studies that demonstrated inhibition of the intact hemisphere with low-frequency TMS (≤1 Hz) or excitation of the damaged hemisphere with high-frequency TMS (≥1 Hz). In both paradigms, function is improved in the more-impaired limb presumably by restoring interhemispheric balance.
Contradicting some previous studies, 8 postinfarct training did not improve motor performance. It is possible that the study was not appropriately powered to demonstrate behavioral differences between groups, but except for the latest time point, the graphs do not even suggest a trend for better performance in the trained group. Alternatively, the lack of a training effect on functional recovery may be a result of the relatively large infarct used in the present study. Typically, focal cortical infarcts using similar methods (ET-1, pial strip, and photocoagulation) and limited to CFA alone result in transient deficits.9,10 The present study examined the effects of a larger infarct encompassing both the CFA and RFA, resulting in a substantially slower rate of spontaneous recovery and presumably maximizing the contribution of the intact, contralesional hemisphere in functional recovery. Even though training sessions were conducted daily for 28 days, the rats did not show any significant signs of improving beyond that of the nontrained rats.
Note in Figure 1, that the performances in the infarct/training and infarct/no-training groups begin to diverge after the fourth week of training. It is plausible that the duration of the present training regimen could be extended to facilitate recovery. However, even with the lack of functional recovery assessed after the fourth week of training, there was a statistically significant physiological effect within the intact hemisphere. ICMS maps may provide a particularly sensitive assessment of the functional status of the intact hemisphere (Figure 2). If rebalancing hemispheric interactions through inhibition of the intact hemisphere accompanies recovery of function, the present results suggest that repetitive use of the impaired limb may have a beneficial effect on recovery during early stages of rehabilitation, prior to observable facilitation of performance.
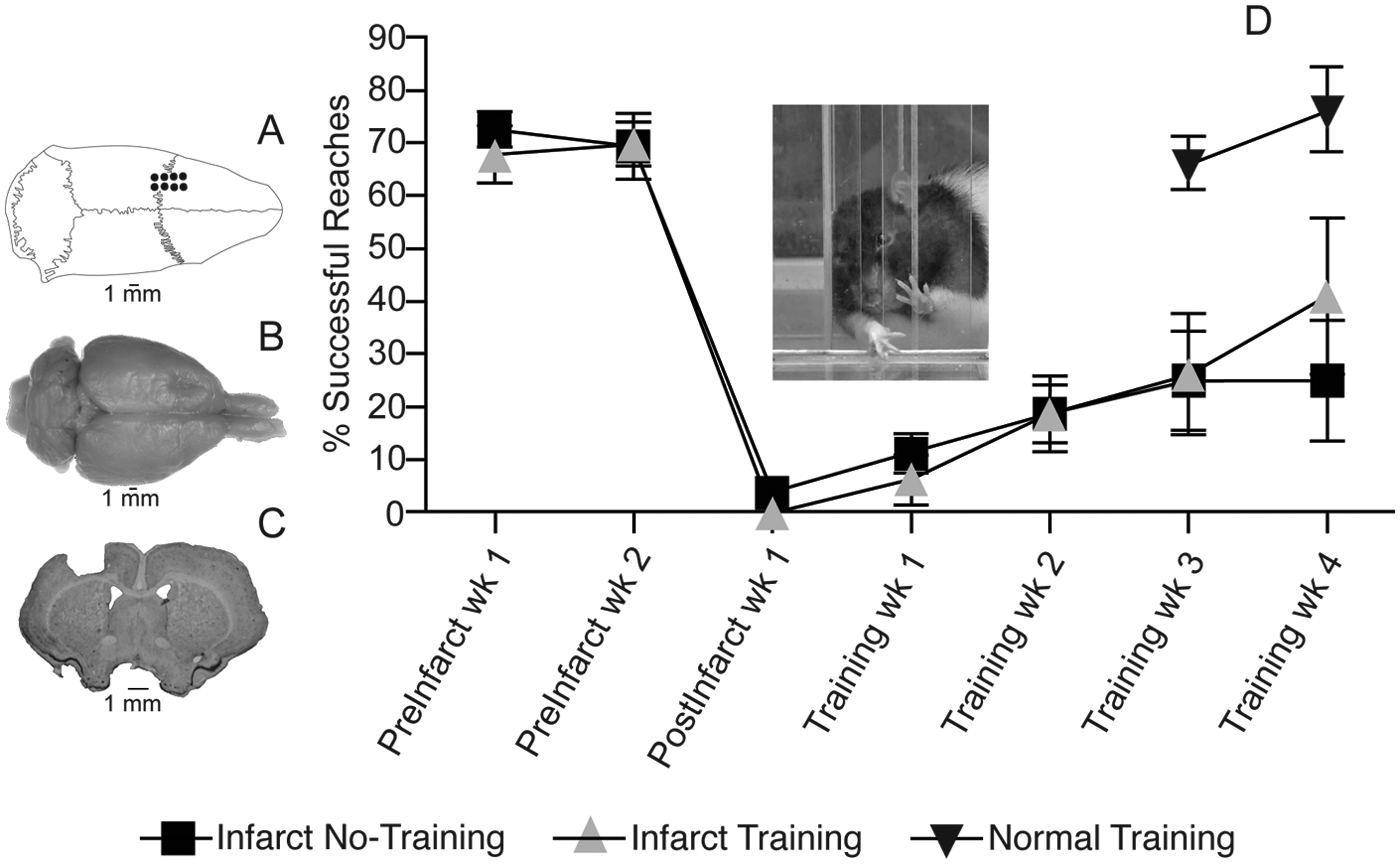
Motor performance after ET-1 lesion in the motor cortex. A. Dorsal view of skull showing location of boreholes for ET-1 injections. B. Dorsal view of fixed brain showing the location of ET-1 lesion in the left motor cortex. C. Coronal section through the level of the caudal forelimb area (CFA) at bregma showing ET-1 lesion. The lesion extended through all cortical layers but did not invade the underlying white matter. The lesion volume was derived from coronal sections imported into NIH Image J software and estimated by the Cavalieri method: infarct/training group = 17.35 ± 0.25 mm3; infarct/no-training group = 17.04 ± 0.49 mm3 (P > .05). Lesion volumes are similar to those predicted based on distribution of ET-1 injections (16 mm3). D. Motor performance on single-pellet retrieval task. Rats were trained daily to reach through an opening in a Plexiglas chamber to retrieve small 45-mg food pellets (Bioserve, Frenchtown, New Jersey) located on a shelf placed above the floor outside the chamber (see inset). 4 Once forelimb preference for unrestricted reaching was established, a movable wall inside the chamber allowed reaching with only the right or left forelimb. Food pellets were delivered 1 at a time for 60 trials during each session of training. A trial ended with a successful reach and retrieval to the rat’s mouth, or after an unsuccessful retrieval when a pellet was contacted but not grasped, or after 5 failed reach attempts without contacting the pellet. A training session ended after 20 minutes regardless of the number of trials completed. Postinfarct training was conducted daily for 28 days; training for the normal group was conducted daily for 14 days (see text). Performance was assessed once per week during 2 weeks of baseline training, 1 week after the infarct prior to rehabilitative training, and once per week after rehabilitative training.
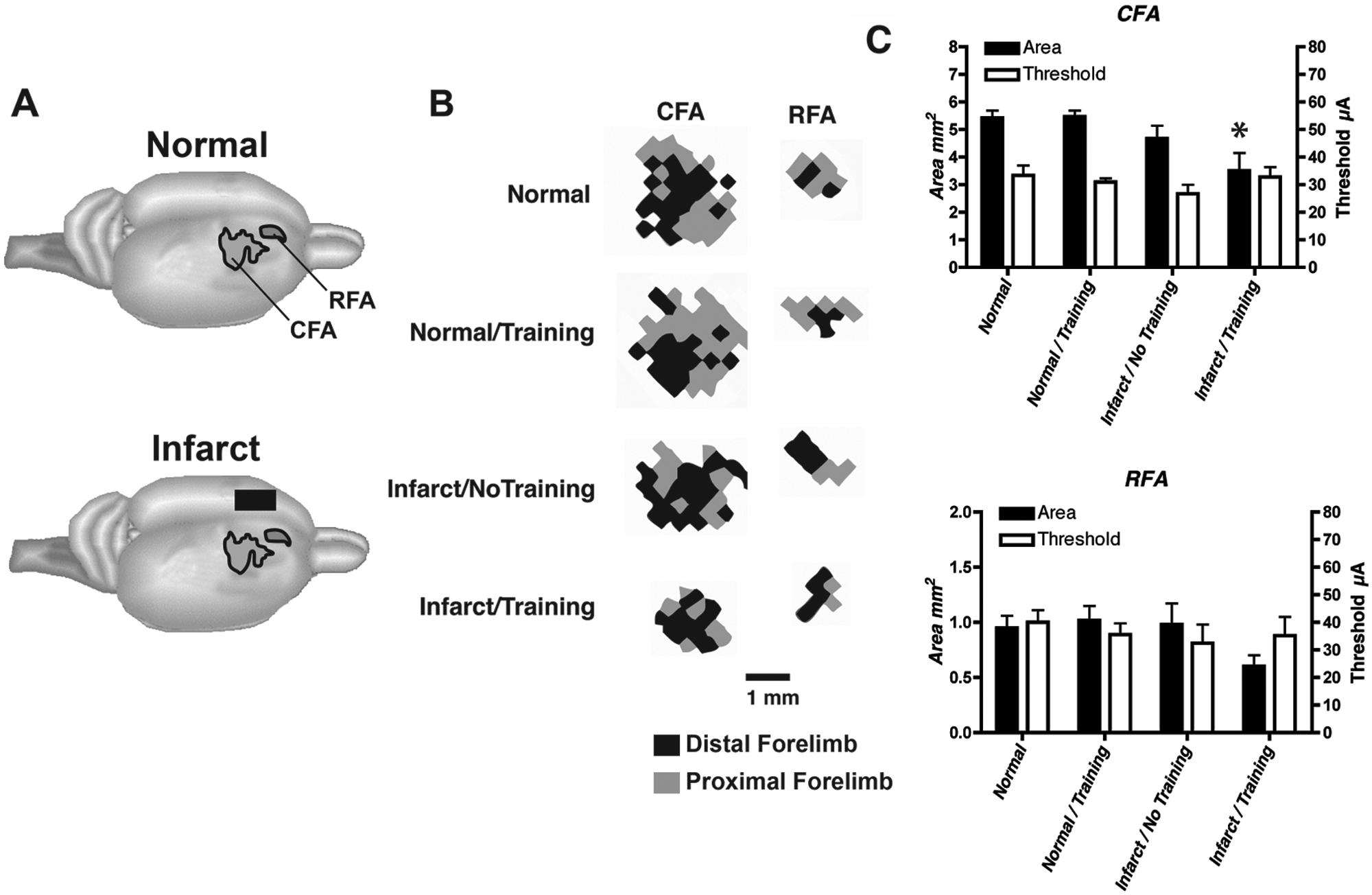
Neurophysiological maps of the caudal and rostral forelimb areas (CFA and RFA, respectively) in the contralateral, intact hemisphere after an ET-1 lesion in the motor cortex. An ICMS stimulus was delivered as a train burst of 13, 0.2-ms cathodal, monophasic pulses, delivered at 350 Hz by a constant-current stimulator (Model BSI-2, BAK Electronics Inc, Mount Airy, Maryland) at a rate of 1 train/s. High-resolution motor maps were derived in both CFA and RFA (350-µm resolution). A. Location of CFA and RFA motor maps. B. Representational maps of CFA and RFA were delineated with customized software and analyzed quantitatively with NIH Image software. Representative maps from each group are displayed. The maps shown are derived from the right hemisphere of rats with a right forelimb preference. The distal forelimb is shown in black, and proximal forelimb is shown in gray. C. Double-Y plot showing map areas and movement thresholds. Areal measurements (mean ± SEM) of CFA and RFA in intact and infarcted rats are presented on the left y-axis. Current thresholds (mean ± SEM) for evoking a forelimb movement in CFA and RFA are presented on the right y-axis. There were no significant threshold differences among the 4 groups in CFA or RFA. Only the infarct/training group had a significantly smaller CFA map relative to normal rats. Abbreviations: ICMS, intracortical microstimulation; SEM, standard error of the mean. *P < .05.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by NIH Grant NS30853 (RJN) and IDDRC Center Grant HD02528.