
Abstract
Miller J, Watrous AJ, Tsitsiklis M, et al. Nat Commun. 2018;9(1):2423. doi:10.1038/s41467-018-04847-9. The hippocampus plays a vital role in various aspects of cognition including both memory and spatial navigation. To understand electrophysiologically how the hippocampus supports these processes, we recorded intracranial electroencephalographic activity from 46 neurosurgical patients as they performed a spatial memory task. We measure signals from multiple brain regions, including both left and right hippocampi, and we use spectral analysis to identify oscillatory patterns related to memory encoding and navigation. We show that in the left but not right hippocampus, the amplitude of oscillations in the 1- to 3-Hz “low-theta” band increases when viewing subsequently remembered object-location pairs. In contrast, in the right but not left hippocampus, low-theta activity increases during periods of navigation. The frequencies of these hippocampal signals are slower than task-related signals in the neocortex. These results suggest that the human brain includes multiple lateralized oscillatory networks that support different aspects of cognition.
Commentary
Are the principles of neuronal coding comparable across species? Or is the human brain code just another language? In trying to understand how our brain represents information, we look for a common syntax with rats and mice. The hippocampus is a Rosetta stone for understanding this neuronal grammar.
In most rodents, representing and memorizing space, time, and experiences by the hippocampus is paced by extracellular theta oscillations at 4 to 8 Hz. This rhythm segregates neuronal firing in sequences of action potentials from the different set of neurons that make an ensemble. 1 Ensemble activity encodes aspects of experience and correlate with spatiotemporal features of the tasks. 2,3 Membrane potential fluctuations in these neurons reflect the underlying rhythmic synaptic currents. 4 Recent data suggest it is even possible to extract such a fine-grained structure from the time–frequency analysis of ongoing oscillations. 5 The question of whether our brain uses similar codes is open though.
Extrapolating the theta syntax from rodents to human has been a challenge. The current view is that human theta oscillations are slow (1-4 Hz) and brief (0.5-1 seconds). 6 Issues regarding accessibility to the entire structure and the healthy human hippocampus with minimally invasive electrode arrays have typically hampered progress. A new study now suggests we should look even closer, at left and right, for our brain code. In their recent work published in Nature Communication, the Jacobs laboratory makes a turn of the screw to the human theta conundrum. 7
Using a virtual reality task that separates periods of focused spatial navigation and memory encoding, Miller et al report on the associated changes of slow (1-4 Hz) and fast (4-10 Hz) theta oscillations. Slow theta activity was more prominent in hippocampal recordings, whereas high theta dominated at the lateral temporal cortex. Strikingly, hippocampal oscillations associated with successful memory stood out on the left, while navigational theta signals dominated the right hippocampus. These observations were possible due to the task design, which provided enough samples for pairwise comparisons of remembered versus forgotten items.
Understanding how spatial memory and navigational signals are encoded in our brain is critical for treating cognitive comorbidities. A distinctive human feature is lateralization of hippocampal function, which differentiates between episodic and spatial memories. 8 Evidence for left–right asymmetries of the vertebrate brain may support emergence of functional lateralization from sensory inputs. 9 Indeed, in rats reared in an enriched environment experience enhances gamma oscillations of the right side, a process that is dependent on N-methyl-D-aspartate receptor (NMDA) receptors. 10 Consistently, some forms of plasticity are lateralized in the rodent hippocampus. 11 Perhaps, these interhemispheric functional differences could arise from microcircuit specialization due to subtle asymmetries in synaptic receptor composition and distribution.
In their report, Miller et al use a predictive model to decode spatial memory from the oscillatory power across frequencies and found cognitive-related signals at hippocampal electrodes. The effect was significant for the 1 to 3 Hz band following item presentation, but it lateralized differently according to the navigational or mnemonic load of the task. However, oscillations associated with items lasted for a few cycles embedded in the broadband spectrum, and no clear difference was found for the high theta band at 3 to 10 Hz. Therefore, even after considering potential interhemispheric differences, human hippocampal theta still remain ill-defined as compared to rats and mice. In these species, high theta oscillations associated with locomotion (∼8 Hz; type 1 theta) are present in the dorsal hippocampus, which is equivalent to posterior in human. 12 In the ventral portion (anterior), theta oscillations are short lived, slower, and more influenced by emotional factors (type 2 theta 13,14 ).
The lack of evidence of sustained human theta oscillations has some echoes in bats. 15 Critically, the CA1 pyramidal cell layer of both humans and bats is dispersed, in contrast to rats and mice. 16 Laminar arrangement, together with the geometrical disposition of dendritic axes and input layers are critical for shaping extracellular signals. This is especially relevant for the human anterior hippocampus, characterized by complex tridimensional bending of different subfields. Given ethical and clinical constraints in sampling the entire human hippocampus, it is difficult to conclude whether differences in theta physiology are due to anatomical, behavioral, or evolutionary factors.
An intriguing finding by Miller et al 7 was the clear spectral peak found at ∼8 Hz in recordings from the left temporal cortex. Others found similar bouts of high theta oscillations in posterior hippocampal locations during walking, 17,18 and the authors’ laboratory has just reported a trend during virtual navigation. 19 Possibly, different theta generators may contribute along the human anterior-to-posterior hippocampal axis at left and right and propagate differently to the overlying temporal cortex partly explaining different spectral dynamics (Figure 1).
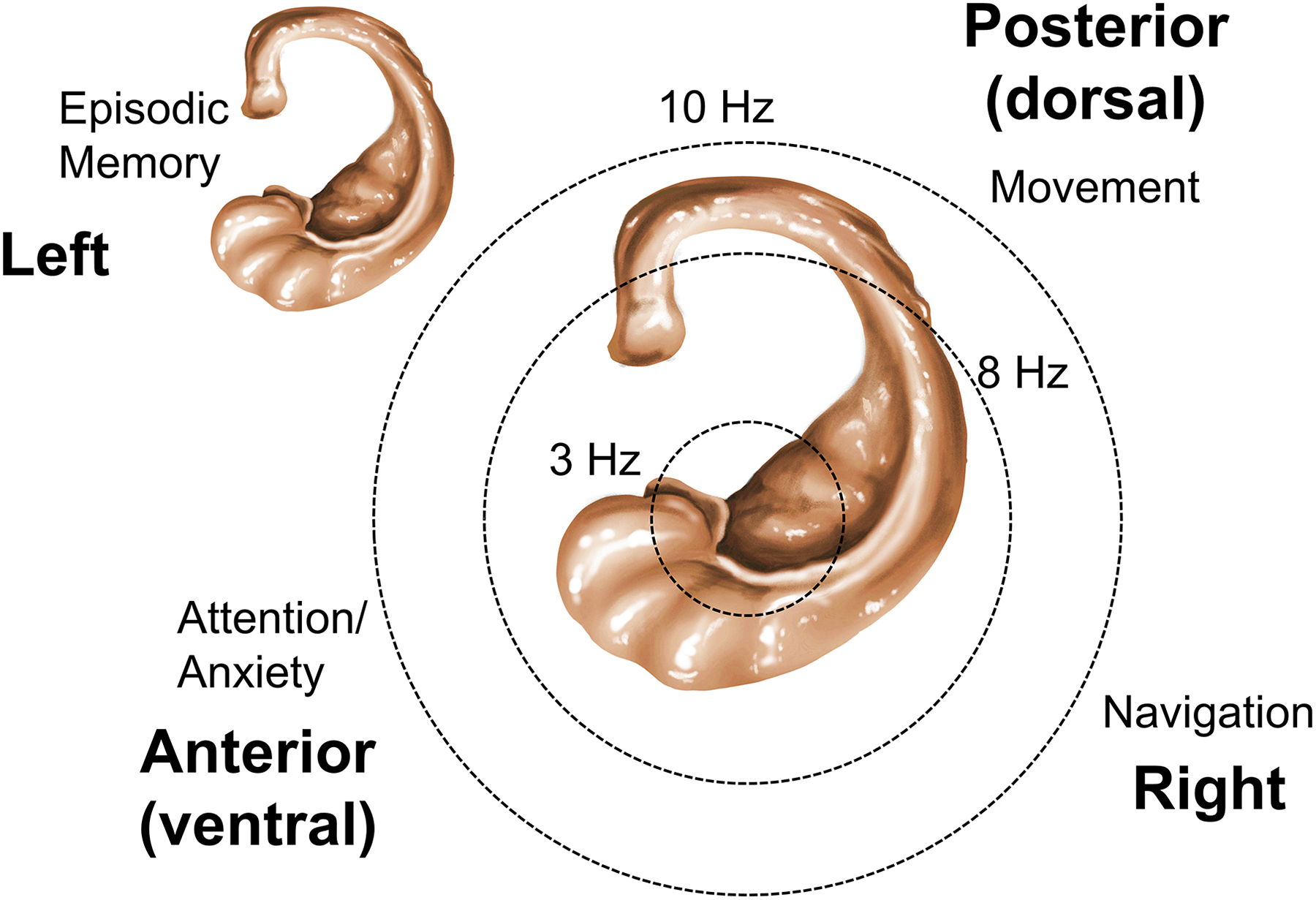
Theta oscillations may be generated differently in the human hippocampus along the anterior-to-posterior axis and in the left and right hemispheres. Theta dominant frequency and behavioral influences (navigational, mnemonic, emotional, and attentional) may be distributed along these axes. Drawing of the human hippocampus by Aimé Menendez de la Prida.
From a systems perspective, human cognition emerges from basic electrophysiological processes that evolved across vertebrates to mammals. The hippocampus is one of our phylogenetically oldest brain regions, but we still need to understand whether cognitive function is implemented similarly across species. Deep in our temporal lobe, there is an answer to this question in the electrical beating of the hippocampus. We all have a neuronal fossil in our head, like a Rosetta stone, to decipher the neuronal grammar.