
Abstract
Objective:
The association of variants in CLU, CR1, PICALM, BIN1, ABCA7, and CD33 genes with late-onset Alzheimer’s disease (LOAD) was evaluated and confirmed through genome-wide association study. However, it is unknown whether these associations can be replicated in admixed populations.
Methods:
The association of 14 single-nucleotide polymorphisms in those genes was evaluated in 280 LOAD cases and 357 controls from the Colombian population.
Results:
In a multivariate analysis using age, gender, APOE∊4 status, and admixture covariates, significant associations were obtained (P < .05) for variants in BIN1 (rs744373, odds ratio [OR]: 1.42), CLU (rs11136000, OR: 0.66), PICALM (rs541458, OR: 0.69), ABCA7 (rs3764650, OR: 1.7), and CD33 (rs3865444, OR: 1.12). Likewise, a significant interaction effect was observed between CLU and CR1 variants with APOE.
Conclusion:
This study replicated the associations previously reported in populations of European ancestry and shows that APOE variants have a regulatory role on the effect that variants in other loci have on LOAD, reflecting the importance of gene–gene interactions in the etiology of neurodegenerative diseases.
Keywords
Introduction
Currently, more than 35.6 million people worldwide have dementia and it is predicted that this figure will tend to double every 20 years. 1 Alzheimer’s disease (AD) is the main form of dementia and it affects mainly elderly people; however, it is not a normal part of the aging process.
While autosomal dominant genetic mutations in amyloid beta precursor protein (APP), presenilin 1 (PSEN1), and presenilin 2 (PSEN2) genes are responsible for 1% to 5% cases of AD, characterized by an early-onset familial AD, 2 over 90% of cases involving the late-onset and sporadic forms are due to the presence of multiple genetic susceptibility variants, environmental factors, and complex interactions between them. 3 Nevertheless, estimates of heritability of up to 80% from twin studies highlight the importance of genetic factors in the development of AD. 4,5
Several case–control studies and more than 40 genome-wide association studies (GWASs) (http://www.genome.gov/gwastudies) have been conducted to date in order to identify genetic susceptibility factors. So far, Allele e4 of apolipoprotein E (APOE∊4) is the genetic risk factor with the strongest effect with more reproducibility among associated variants, having a population attributable risk of 25.5%. 6 Other loci with modest effect have been associated with AD in several studies 6 -11 and collected in the AlzGene platform (www.alzgene.org). The results obtained so far have shown new biological pathways involved in the development of AD such as lipid metabolism, inflammation, and cell membrane transport. 12 Although the relationship between these new genes and pathophysiological hypotheses for AD (amyloid cascade hypothesis and τ) was initially unclear, it has now been established that some genes may have a pleiotropic effect and might have a role on amyloid metabolism. For example, it has been detected that the possible involvement of CLU (clusterin or apolipoprotein J) in AD pathology shows a strong analogy with apolipoprotein E (APOE). CLU directly interacts with amyloid-β (Aβ) and crosses the blood–brain barrier, helping Aβ elimination, 13 and like APOE, it acts as a chaperone protein preventing aggregation. 14 Moreover, CLU has an established role as an inhibitor of cytolysis by complement. 15
Most of these studies have been conducted in populations of European origin, so care must be taken when extrapolating the findings from 1 population to another, as different demographic processes produce differences in the frequencies of the potential risk alleles among populations. 16 Colombian and Latin American populations in general exhibit admixture of European, Native American, and African ancestry as a consequence of the population dynamics that started from the times of European colonization. This is how the admixture of Latin American populations can be biethnic or triethnic, considering the degree of contribution of European, Native American, and African ancestral populations. 17 -19
In the present study, we determined the genetic susceptibility of the Colombian population to the late-onset AD (LOAD), through an evaluation of variants in genes previously reported in GWAS taking into account the effect of its ancestral composition.
Methods
Study Population
Participants were sampled in different facilities in 2 cities (Neiva and Medellin) between the years 2010 and 2014. In Neiva, they were referred by neurologists from the Hospital Universitario de Neiva and the group of Neuropsychology of the Universidad Surcolombiana. In Medellin, participants were referred by the group of Neuroscience of the Universidad de Antioquia.
Potential participants were assessed by an interdisciplinary group, which included neurological, neuropsychological, and psychiatric evaluation. The diagnosis of possible AD was made according to the international diagnostic criteria Diagnostic and Statistical Manual of Mental Disorders (Fourth Edition). 20 Similarly, the age-matched control group was defined. Controls were collected from a collaborative group in each region and were confirmed as healthy by medical examination and neuropsychological assessment by expert neurologists and neuropsychologists.
After assessment of the participants and their medical history, all individuals over the age of 60, with no psychiatric history (eg, bipolar affective disorder, schizophrenia) or any other disease that could have influenced the results of the cognitive tests (eg, Parkinson’s disease, multiple sclerosis, epilepsy, meningitis), were included. Patients (cases) aged between 60 and 65 were defined as sporadic, given that none of their first-degree relatives had a family history of dementia. Given the fact that there is a group in Medellin (Antioquia) that have E280A mutation in the PSEN1 gene, all participants from Antioquia were evaluated for the presence of the mutation. However, due to limitations of the study design, other mutations related to EOAD were not taken into account.
The sampled population was fully informed about the study and agreed to join the research after signing the informed consent (for patients, informed consent was signed by relatives or by a legal representative responsible), which was previously approved by the ethics committees of the respective centers (Universidad Surcolombiana and Universidad de Antioquia), in compliance with the Declaration of Helsinki guidelines.
Genotyping
Peripheral blood samples were collected in two 4-mL EDTA tubes for DNA extraction from leukocytes using a phenol–chloroform method previously described. 21 Fourteen single-nucleotide polymorphisms (SNPs) previously reported in GWAS in the CR1 (rs4844610), BIN1 (rs7561528, rs744373), CLU (rs11136000, rs1532278, rs9314349), PICALM (rs677909, rs541458, rs3851179), ABCA7 (rs3752229, rs3764650, rs3752246), and CD33 (rs3826656, rs3865444) genes were selected. This study also included 2 APOE gene variants (rs429358 and rs7412).
Genotyping of SNPs was performed by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). Products generated were visualized through electrophoresis on agarose gels stained with ethidium bromide (0.5 μg/mL) using a suitable molecular weight marker for the expected sizes. For variants in APOE, visualization of restriction products was performed by vertical electrophoresis in polyacrylamide gels postcleavage with HhaI, as described in a previous protocol. 22 Controls of each genotype were used in all runs.
Moreover, given the genetic background of the Colombian population, the proportion of European, Native American, and African ancestry was estimated using a panel of 30 ancestry informative markers (AIMs). These markers were chosen from panels for Latino populations reported by Parra et al, 23 Shriver et al, 24 and Molokhia et al. 25 The AIMs used have high discrimination power (δ > 45%) between European, African, and Native American ancestries. Genotyping of the 30 AIMs was performed by PCR-RFLP or PCR and capillary electrophoresis on an ABI-PRISM 310 (Universidad de Antioquia) genetic analyzer (Perkin Elmer - Applied Biosystems), according to the type of marker SNP or indel. Details about the location and allele frequencies of the AIMs in ancestral and study populations (cases and controls) are shown in Supplementary Table 1.
All genotype readings both for GWAS-reported variants and AIMs were performed by 2 independent researchers and were done with no previous knowledge of the group to which the samples belonged. The genotyping process was repeated in those cases in which there was no clarity regarding the genotype or if there was no consensus in the reading.
Statistical Analysis
Descriptive statistics were estimated, and comparisons between the case and control groups were established using a Student’s t test for continuous variables and a χ2 test for categorical variables. Comparisons between groups for continuous variables that were not normally distributed were performed applying nonparametric tests. Statistical Package for the Social Sciences (SPSS 18) was used for these analyses. 26
Allelic frequencies, genotypic frequencies, and the Hardy-Weinberg equilibrium (HWE) were calculated using PLINK v 1.07 (http://pngu.mgh.harvard.edu/∼purcell/plink/index.shtml). 27 Only markers with a call rate over 90% and without significant deviations from HWE equilibrium after a Bonferroni correction were used for estimating ancestry and for association analyses.
The individual and average ancestry was estimated using ADMIXMAP v3.8 for Windows. 28 The ADMIXMAP employs a combination of conventional (frequentist) and Bayesian methods to model the proportion of individual admixture from a prior distribution of allele frequencies of AIMs in the ancestral populations. This is done in order to obtain a posterior distribution of the ancestry contribution to each of the samples. To estimate the ancestry of the sample under study, a model that involved a population derived from 3 ancestral populations (African, European, and Native American) and nondispersion of allele frequencies was implemented. 29
Genetic associations between SNPs and LOAD were evaluated with SNPassoc included in the statistical package R v 3.1.1 (http://www.r-project.org/). 30 The Akaike information criterion implemented in SNPassoc was used to determine the best genetic model for each marker analyzed. 31 Genetic models were defined taking the minor (ie, less frequent) allele as the reference allele. For example, the dominant model compared carriers of at least 1 copy of the minor allele with noncarriers, the additive model compared the number of copies of the minor allele (0, 1, or 2), and so on. Multivariate logistic regressions models were built, including covariates such as age at the time of the evaluation, gender, presence of APOE∊4 allele, and admixture (using 2 ancestral components to avoid colinearity). The robustness of the associations detected was checked applying a permutation test with 1000 permutations.
On the other hand, the interaction between the presence of APOE∊4 and each variant was assessed using logistic regression including gender, age, and admixture as covariates. Likewise, a stratified analysis was completed for cases and controls in order to determine whether the presence of the APOE∊4 allele significantly modified associations between the variants and LOAD.
Results
Characteristics of the Study Groups
We analyzed a total of 637 individuals over 60 years old, who were divided into groups of 280 cases and 357 controls. Sociodemographic characteristics of the study population and comparisons between groups for the evaluated variables are presented in Table 1. There were no differences by gender, but for the age and presence of APOE∊4 variables, there were significant differences between case and control groups. The mean age was higher in cases compared to the control group (P < .001). Consistent with previous reports, a higher frequency of the APOE∊4 risk allele was found in cases (46.1%) compared to controls (22.8%). Also, when it comes to the distribution of APOE genotypes, a higher frequency of ∊3/∊4 and ∊4/∊4 genotypes was observed in cases compared to controls. The genotype ∊3/∊3 was more common in controls (70.4%). The genotype ∊2/∊2 was not present in the sample.
Characteristics by Case–Control Status in the Study Population.
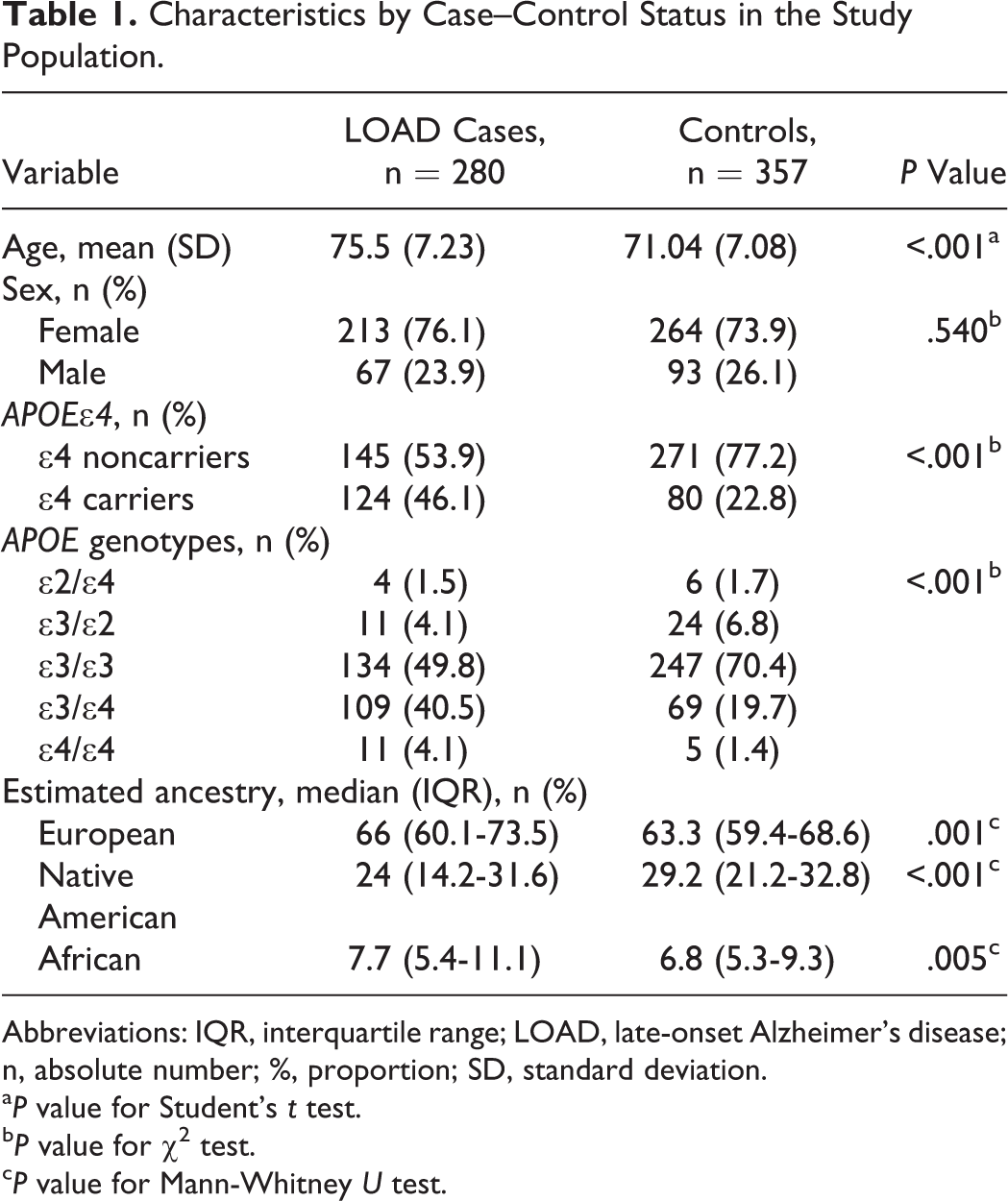
Abbreviations: IQR, interquartile range; LOAD, late-onset Alzheimer’s disease; n, absolute number; %, proportion; SD, standard deviation.
a P value for Student’s t test.
b P value for χ2 test.
c P value for Mann-Whitney U test.
In regard to the estimation of ancestry, HWE was tested for each AIM in the global population and independently in case and control groups. After applying a Bonferroni correction for multiple comparisons, a deviation from HWE was observed in the global population and in the case and control groups for the MID52 marker, so it was excluded from further analyses. Significant differences in ancestry proportions for the 3 ancestral components were found (Table 1). Native American ancestry was higher in controls than in cases (P < .001), and European and African ancestries were higher in cases than in controls (P = .001 and .005, respectively).
Association Analysis
All SNPs were found to be in HWE in both cases and controls. Since age, presence of APOE∊4 allele, and admixture proportions showed significant differences between cases and controls, association analyses were carried out using models of multivariate logistic regression in which the gender variable was also included (Table 2). After making adjustments for age, gender, presence of APOE∊4 allele, and admixture covariates, variants in BIN1, CLU, PICALM, CD33, and ABCA7 genes were significantly associated with differential risk to have LOAD (Table 2). These results contrasted to a certain extent with exploratory analyses performed without adjusting for covariates, which showed that the presence of population structure caused by admixture events was confounding the association due to inherent differences between cases and controls that are not attributable to the state of the disease (data not shown). However, upon permutation testing, none of the detected genetic associations remained significant.
Genetic Associations With LOAD.
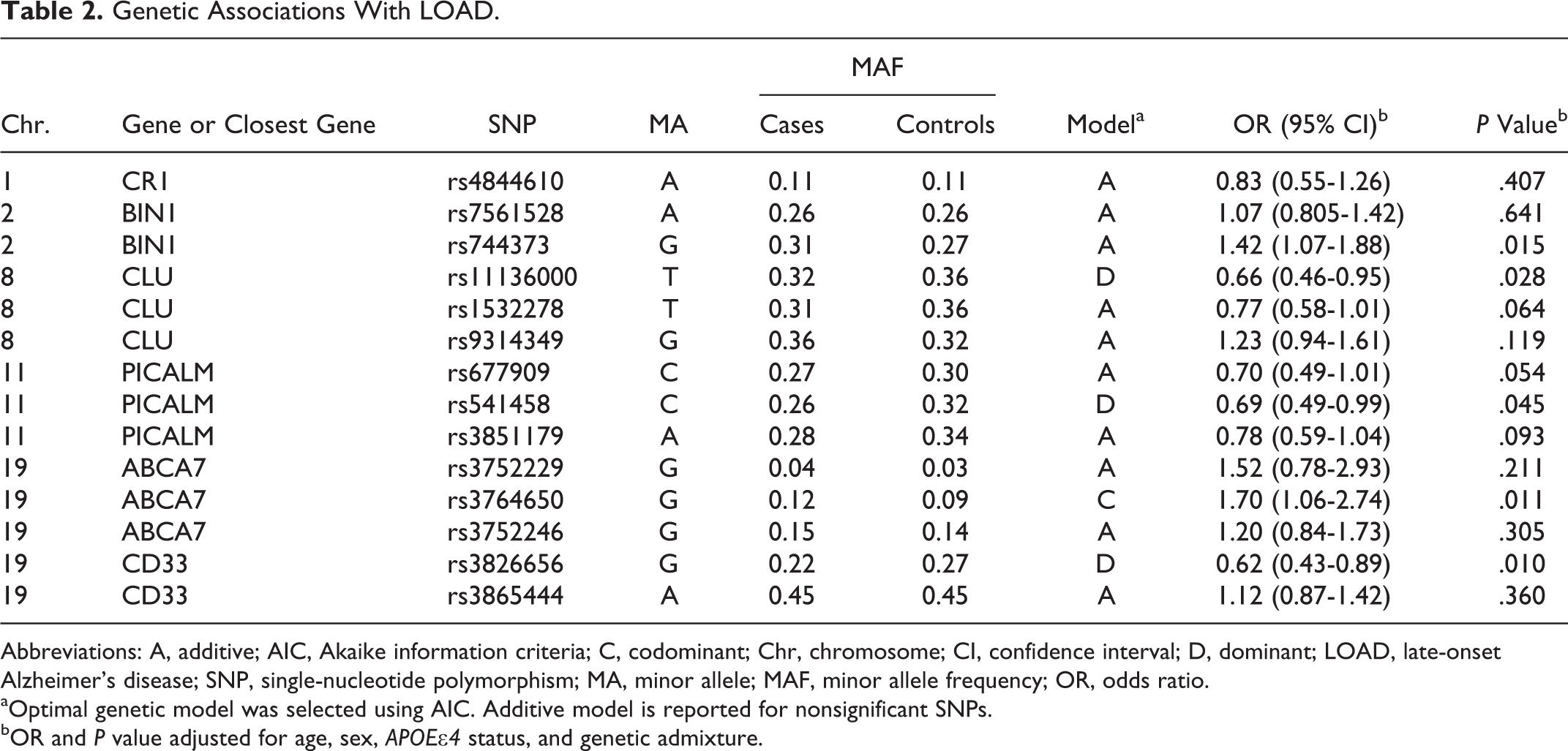
Abbreviations: A, additive; AIC, Akaike information criteria; C, codominant; Chr, chromosome; CI, confidence interval; D, dominant; LOAD, late-onset Alzheimer’s disease; SNP, single-nucleotide polymorphism; MA, minor allele; MAF, minor allele frequency; OR, odds ratio.
aOptimal genetic model was selected using AIC. Additive model is reported for nonsignificant SNPs.
bOR and P value adjusted for age, sex, APOE∊4 status, and genetic admixture.
Interaction With APOE∊4
The presence of the APOE∊4 allele modified the associations of SNPs in the CR1 and CLU genes with LOAD (Table 3). The 3 variants evaluated in the CLU gene (rs11136000, rs1532278, and rs9314349) as well as the rs4844610 variant in the CR1 gene showed statistically significant interactions with the presence of the APOE∊4 allele.
Interaction Analyses Between SNPs and APOE∊4 Status.
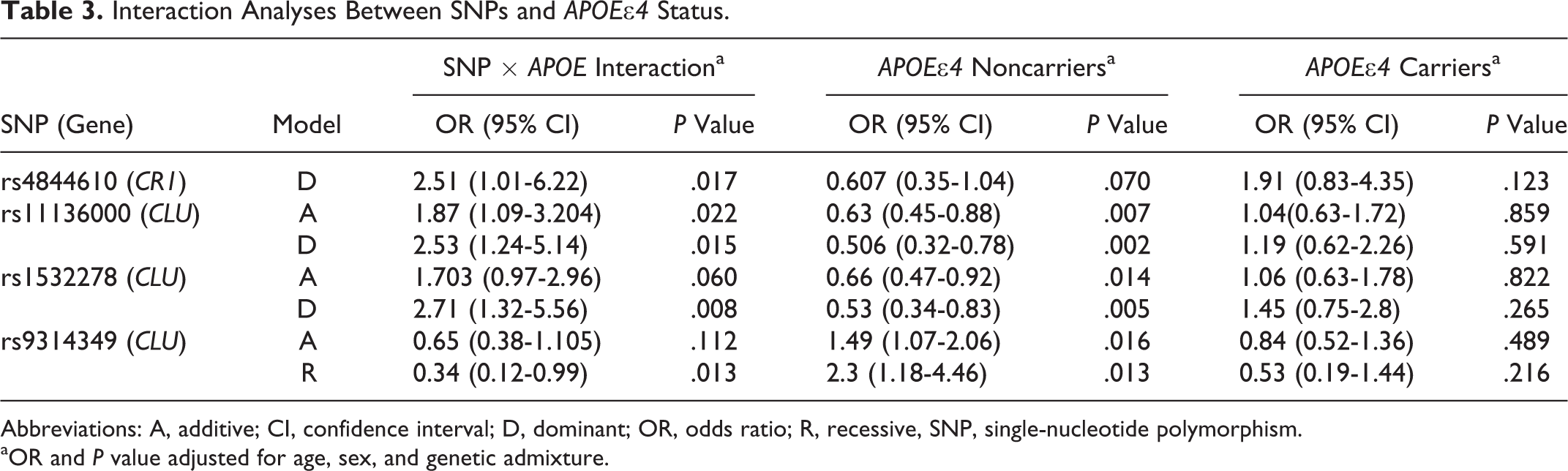
Abbreviations: A, additive; CI, confidence interval; D, dominant; OR, odds ratio; R, recessive, SNP, single-nucleotide polymorphism.
aOR and P value adjusted for age, sex, and genetic admixture.
Stratifying the data by APOE∊4 status (carriers and noncarriers of the ∊4 allele) in cases and controls and adjusting for age, gender, and admixture covariates, significant associations were observed only for CLU variants in noncarriers. Among carriers of APOE∊4, these variants did not show any effect on the risk of LOAD. Minor alleles of the rs11136000 and rs1532278 CLU variants were associated with protection under the additive and dominant models, while the rs9314349 variant in the same gene was associated with an increased risk in the additive and recessive models. After the permutation test, the associations found for the rs11136000 and rs1532278 SNPs in the group of noncarriers of APOE∊4 remained significant (additive model corrected, P = .04 and dominant model corrected, P = .02; dominant model corrected, P = .03, respectively). The rs4844610 variant in CR1 did not show significant associations with LOAD in the stratified analyses, though this SNP showed values close to significance (P = .071) among noncarriers.
Discussion
In the present study, the association between the status of LOAD and 14 gene variants in 6 genes involved in 3 biological pathways (inflammation, cholesterol metabolism, and transport at the level of membrane endocytosis) recently associated with the pathophysiology of the disease was evaluated in the Colombian population. In addition to the known confounding variables such as age, gender, and the presence of the APOE∊4 allele, we took into consideration the genetic background of the admixed population in order to adjust the associations. The results obtained indicate that variants in genes acting on the 3 pathways are associated with susceptibility to the development of LOAD in the Colombian population.
Given the differences in the prevalence of AD, allelic frequencies, and the pattern of linkage disequilibrium (LD) between populations, and given that most genetic studies for AD have been carried out in populations of European descent, replication studies in populations with a different ethnic composition are necessary to determine the true effect size of a given locus in a specific population. 32 This is highly relevant when taking into consideration the context of the potential gene–gene and gene–environment interactions that could modify the risk of LOAD.
A significant association between the G allele of the BIN1 rs744373 variant and an increased risk of LOAD was observed, consistent with that reported in previous studies for Caucasian 8,9,11,33 and Japanese 34 populations. However, the effect size observed in our study may be somewhat inflated due to the sample size, compared to other studies. In contrast, no association with this variant was found in Korean 34,35 and Han 36 populations. Other variants in BIN1 have shown significant association in Caucasians, 7,10,37 Hispanics from the Caribbean, 38 and the Africans. 39 The bridging integrator 1 (BIN1) protein has been shown to be involved in receptor-mediated endocytosis and endocytic recycling, which could be affecting APP traffic. 40,41 It has also been reported a possible interaction between BIN1 and τ in which the major BIN1 isoform expressed in neurons was associated with modified species of τ found in AD. 42
Phosphatidylinositol Binding Clathrin Assembly Protein (PICALM) is also involved in the mechanism of receptor-mediated endocytosis, and it has been related to the right synaptic transmission 43 and to the trafficking and processing of APP. 44 In our study, of the 3 PICALM variants evaluated, only the C allele of rs541458 was associated with a protective effect against AD, in line with the previous reports of the Caucasians. 45,46 Associations between LOAD and other variants in this gene have been reported in Caucasian, 9,10,47,48 Asian, 34,35 and African 49 populations.
The T allele of rs11136000 in CLU was significantly associated with a reduction of LOAD risk under the dominant model. Two large GWASs in 2009 first reported an association of this variant, with an average odds ratio (OR) of 0.84 for the T allele in populations of Belgium, Finland, Italy, and Spain, 6,8 which is consistent with that reported in this study. Subsequently, other studies reported the same association in Caucasian 9,45 -47 and Chinese 50 populations. Other CLU variants have also been associated with AD. 7,10,38 The CLU is a multifunctional protein that plays an important role in the metabolism and elimination of Aβ-peptide similar to that of APOE. 13 It has been determined that the T allele in the rs11136000 SNP had functional relevance being associated with an increased expression of the CLU1 isoform of the gene. 51
Under a codominant model, the heterozygous genotype T/G of the ATP binding cassette subfamily A member 7 (ABCA7) rs3764650 variant was associated with an increased risk. This same effect was detected for the G allele in previous GWAS in Caucasian 52 and African American 49 populations. In contrast, this variant has shown contradictory results and indeed to have an opposite effect in the Han population. 36,53 -55 In the present study, the heterozygous genotype association may be the result of the low frequency of the G/G genotype in our sample. Discrepancies across populations in the risk alleles and the models that explain the associations found may be due to the differences in the frequency of the alleles that are in LD with the causal variants. The frequency of rs3764650-G allele is 0.33 in Asians (0.27 Han population), 0.28 in Africans, 0.11 in Europeans, and 0.08 in Native Americans. In Colombia, the frequency reported by the 1000 Genomes Project is 0.09, the same found in the present study in controls. Other variants in ABCA7 were significantly associated with AD in Caucasians. 7,37 According to recent studies, the mechanism by which ABCA7 is associated with LOAD is the elimination of Aβ, since it has been shown that expression of ABCA7 increases microglial phagocytosis of Aβ, which is consistent with the increased accumulation of Aβ in ABCA7-deficient mice. 56 In turn, the G allele associated with an increased AD risk was significantly associated with a decreased expression of ABCA7. 57
The G allele of the rs3826656 variant, located near the 5′ region of the CD33 gene, was associated with protection under the dominant model. Bertram et al. (2008) first reported the G allele in this variant as associated with AD under a dominant model in the same way than the present study. 58 However, the CD33 variant commonly associated with AD protection was rs3865444, 7,10,11,36,59 for which no significant association was found in this work. A study undertaken in Han population replicated the association between rs3826656 and LOAD in carriers of the APOE∊4 allele. Nevertheless, the allele associated with protection was the A allele and the recessive genotype A/A, opposite to our findings. 60 It is noteworthy that allele frequencies for this variant differ greatly among populations. While in Europeans the A allele has a frequency of 0.78, in the Han population the frequency is 0.32. The Colombian population has a frequency of 0.76, as described in the 1000 Genomes Project, similar to that in Europeans and consistent with our study (0.74 in controls). Additionally, the association with the opposite allele on the same marker may be due to the differences in the LD structure, so that the LD between the actual causal variant and the evaluated variant (rs3826656) may differ among populations, or as a consequence of the observed interaction with the APOE∊4 allele. 61 Recently, 2 studies that analyzed the contribution of this gene to the pathology of AD were published. Griciuc et al showed that the minor allele of rs3865444 (T), which confers protection against AD, was associated with decreased levels of CD33 and decreased levels of insoluble Aβ-42 in the brain. Thus, CD33 could be inhibiting phagocytosis of Aβ-42 by microglia. 62 The second study found an association of the rs3865444 risk allele (C) with an increased expression of CD33 on the surface of monocytes, increased CD33-immunoreactive microglia, a decrease in the internalization of Aβ-42, and an accumulation of amyloid plaques, confirming previous findings. 63
In addition to the associations found, the APOE∊4 allele was confirmed as a genetic risk factor with a remarkable effect on the development of AD consistent with previous reports in Colombian 64,65 and other Latino populations. 66,67 In the present study, ∊4 allele frequency was 0.608 in cases compared to 0.392 in controls, conferring a risk 3 times higher compared to the other alleles (OR = 3.11, 95% confidence interval = 2.15-4.51, P < .0001), adjusted for age, gender, and admixture.
Moreover, the 3 CLU variants evaluated (rs11136000, rs1532278, and rs9314349) and a CR1 variant (rs4844610) showed significant interactions with the APOE∊4 allele. When performing the stratified analysis, the association with the 3 CLU variants was detected only in the group of noncarriers of APOE∊4. Although there was no significant association for CR1, a trend toward association was seen in the group of noncarriers of APOE∊4. Lambert et al reported a significant interaction between 3 CLU variants (rs11136000, rs2279590, and rs9331888) and 2 CR1 variants (rs6656401 and rs3818361) with APOE∊4. Another recently published study confirmed the CLU and CR1 interaction with APOE, finding such association in a stratified analysis. 68 At the biological level, there is evidence of the interaction between CLU and APOE, which can be given by their involvement in cholesterol metabolism and/or their role in eliminating the Aβ peptide. Moreover, it has been noted that reduced levels of CLU are correlated with the number of ∊4 alleles, suggesting a compensatory induction of CLU in the brain of individuals with AD, with ∊4 allele of APOE presenting low brain levels of APOE. 69 Regarding CR1, it has been found that the interaction CR1 × APOE modulates the accumulation of Aβ even in older individuals who are cognitively normal. 70 Likewise, an investigation reported an interaction between CR1 (rs4844609) and APOE∊4, stimulating an episodic memory decline and mediating an increase in Aβ pathology. 71 This suggests that elimination of Aβ is the mechanism in which CR1, CLU, and APOE converge, causing an effect on AD pathology; however, the biology of these interactions remains to be determined.
Here, we confirm the modulation of the genetic effect of susceptibility genes for AD (such as CLU and CR1) by the presence of the APOE∊4 allele, which highlights the importance of gene–gene interactions on the genetic regulation of neurodegenerative diseases and suggests that they may be key to identify the “missing heritability” of complex diseases.
In the present study, we observed population stratification in cases and controls showing significant differences in the proportions of genetic ancestry. This finding opens the way for a new study about the ancestry effect on AD in admixed populations, and the implementation of mapping by admixture LD would be a great contribution to evaluate this issue. Previous studies have shown that associations between genetic ancestry and risk of complex traits in recently admixed populations (such as the Colombian population) are due, at least in part, to genetic differences between populations of different continental origin. 72
Stratification was corrected in the association analyses calculating the proportions of individual admixture obtained using 29 AIMs. The power of discrimination between populations of this panel of markers is very robust, and the results of the ancestry estimation are consistent with previous reports for our population using different panels of markers. 18,73 -75
Furthermore, this research has an important limitation with regard to the sample size. It is clear that small sample sizes reduce the power to detect significant associations with less frequent variants or with variants with a small effect on the presentation of LOAD, and they also increase the risk of reporting false positives. However, given the fact that the analyzed variants have been previously reported in GWAS, this risk is greatly reduced and the reliability of the results increases as the direction of the association and the magnitude of the effect were similar to the first studies. Despite the limited sample size used in our study, it had a statistical power of 71% to identify variants conferring a risk (OR) of ≥1.5 (or ≤0.67), for a significance level of .05, with an allele frequency of 0.2, for markers in complete LD with the causal variant (calculated using Genetic Power Calculator, http://pngu.mgh.harvard.edu/∼purcell/gpc/). 76
Although more than a hundred years have passed since the first report of AD and even though there is still no effective treatment, great breakthroughs have been made in understanding its pathophysiology. Such breakthroughs will allow the picture to be extended in terms of the therapeutic approach and will even allow the treatment of biological pathways common to different neurodegenerative diseases. 77 However, the genetic context of the study population in terms of the study of a complex disease like LOAD and differences in individual genetic risk should be carefully considered.
Footnotes
Acknowledgments
The authors are very grateful to the volunteers who participated in this project. Special thanks to Hospital Universitario de Neiva, Grupo de Neuropsicología de la Universidad Surcolombiana, and Grupo de Neurociencias de la Universidad de Antioquia for allowing recruitment of participants and clinical information access.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: D.J.M. was supported by Colciencias National Doctoral scholarships. This research was supported by a grant from Colciencias, Departamento Administrativo de Ciencia, Tecnología e Innovación, República de Colombia (Grant 1124-493-26212).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.