
Abstract
Objective:
To explore the correlation of formyl peptide receptor 1 expression with drug resistance and the functions of formyl peptide receptor 1 in drug-resistant bladder cancer.
Methods:
Expression of formyl peptide receptor 1 in T24 and T24/DDP cisplatin-resistant bladder cancer cell lines was tested by quantitative real-time Polymerase Chain Reaction and Western blotting. After incubation of T24/DDP with N-formyl-Met-Leu-Phe, the phosphor proteins were tested by Western blot analysis. We characterized the functions of formyl peptide receptor 1 in T24/DDP cells by assessing proliferation, migration, and changes of cell cycles.
Results:
Formyl peptide receptor 1 was expressed in both T24 and T24/DDP, and it was overexpressed in T24/DDP compared with T24. Formyl peptide receptor 1 activation promoted the expression of the messenger RNA of resistance-related proteins, such as multidrug resistance-associated protein 1 (MRP1) and lung resistance-related protein (LRP). The expression of 4 signal pathway proteins were upregulated: signal transducer and activator of transcription 3, Janus kinase 2, extracellular regulated protein kinases, and protein kinase B, while the expression of phosphatidylinositol 3-kinase was observed to be downregulated in drug-resistant bladder cancer cells. Formyl peptide receptor 1 activation also improved the expression of phospho-signal transducer and activator of transcription 3 and phospho-extracellular regulated protein kinases 1/2 and promoted the proliferation and migration of T24/DDP cells. In addition, formyl peptide receptor 1 inhibition led to the change in the cell cycle in T24/DDP.
Conclusions:
The overexpression of formyl peptide receptor 1 may be related to drug-resistant bladder cancer and promotes the deterioration of drug-resistant bladder cancer.
Introduction
Bladder cancer is the most common tumor in the urinary system. Currently, clinical treatment of bladder cancer utilizes cystectomy supplemented with chemotherapy. With the development of treatment, the mortality of bladder cancer has decreased significantly, but the recurrence rate of bladder cancer is still high. Clinically, 70% to 80% of patients with bladder cancer belong to nonmuscle invasive bladder cancer, and most patients will relapse within 5 years; the prognosis is poor. Due to the lack of effective treatment to reduce the recurrence of bladder cancer and drug resistance, the quality of survival and life of patients has not undergone a satisfactory improvement. Drug resistance is part of the major causes of bladder cancer recurrence. Currently, a number of mechanisms related to drug resistance have been reported, including reducing the accumulation of drugs in tumor cells, the increase in the antiapoptosis ability, the improvement in the abilities of detoxification and restoration, and the overexpression of certain Multiple drug resistance (MDR) proteins, such as the ATP binding cassette (ABC) transporter. 1 Our laboratory has successfully established a stable bladder cancer resistant cisplatin cell line (T24/DDP), but its resistance mechanism has not been thoroughly elucidated to date. 2,3
Human formyl peptide receptor 1 (FPR1) and its variants, FPR2 and FPR3, belong to the 7 transmembrane G-protein coupled receptor family. These receptors were first described in immune cells and are involved in the regulation of the innate immune response. 4 -6 In addition, FPRs are also expressed in certain tissue cells, such as hepatocytes, lung epithelial cells, and stellate cells. 7 -9 N-Formyl-Met-Leu-Phe (fMLF) is a specific exogenous agonist of FPRs. N-Formyl-Met-Leu-Phe is highly compatible with FPR1, binds to FPR2 in a low affinity mode, and does not bind to FPR3, and Boc-MLF is an antagonist of FPR1.
In recent years, it has been demonstrated that FPRs play an important role in the development and progression of tumors. It has been reported that the high expression of FPR1 is associated with a high risk of tumor formation and poor prognosis. 7,10 -13 For instance, in human gastric carcinoma (GC) cells, overexpression of FPRs, upon activation, promotes the epithelial–mesenchymal transition, migration, and proliferation. 13 However, it is interesting that in animal experiments, GC cells with silenced FPR1 formed more rapidly growing tumors in immune-compromised mice. The study showed that FPR1 may promote the expression of antiangiogenic factors by GC cells, thereby reducing the blood supply to the tumor. In contrast to GC cells, a different effect of FPRs was observed in malignant tumors. In human glioma, FPR1 was observed to be overexpressed by glioblastoma multiforme cells of the more highly malignant. 12 However, to the best of our knowledge, the relationship between FPRs and drug-resistant bladder cancer has not been studied. Our study seeks to explore this relationship and study the specific roles and mechanisms of FPRs in drug-resistant bladder cancer.
Materials and Methods
Cell Culture
T24 human bladder cancer cell line was purchased from the Chinese Academy of Sciences Culture Collection. The T24 cell line was cultured in RPMI 1640 medium (Gibco; USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Ausbian, Australia) at 37°C and 5% CO2. The drug-resistant cell line was T24/DDP, which was established by a concentration gradient increment method, and finally, the concentration of cisplatin was 0.8 µg/mL.
RNA Purification and Quantitative Real-Time PCR Analysis
Total RNA from T24 cells and T24/DDP cells was purified with Trizol reagent (Ambition, Life Technologies, USA), according to the manufacturer’s instructions. Under the function of the M-MLV reverse transcriptase (Promega, USA), 2 µg of total RNA was used to synthesize complementary DNA using Oligo (dT) (Dingguo Biotech, China), dNTPs (Genview, USA), and RNase inhibitors (Dingguo Biotech, China). Quantitative real-time PCR (qRT-PCR) was conducted with TransStart Top Green qPCR SuperMix Kit (Beijing TransGen Biotech, China) and the LightCycler 480 Real Time PCR System. The PCR primer sequences used were as follows: GAPDH forward: 5′-TTTGGTATCGTGGAAGGACT-3′ and reverse 5′-AGTAGAGGCAGGGATGATGT-3′; FPR1 forward: 5′-TGGGACGACATTGGCCTTTC-3′ and reverse 5′-GGATGCAGGACGCAAACAC-3′; FPR2 forward: 5′-CTGTACTTTCAACTTTGCATCC-3′ and reverse: 5′-ATTTCCCAACTCCACTTACC-3′; FPR3 forward: 5′-AGGATCTAAGCTGGTGGTGT-3′ and reverse: 5′-GTGACTCCGTGGACTAGCA-3′; MRP1 forward: 5′-TTGCCGTCTACGTGACCATT-3′ and reverse: 5′-AGGCGTTTGAGGGAGACACT-3′; LRP forward: 5′-TATGTGCCATCTGCCAAAGT-3′ and reverse: 5′-CATGTAGGTGCTTCCAATCA-3′; P-gp forward: 5′-GATATTGCCTGGTTTGATGA-3′ and reverse: 5′-TGCATTTTGTGTTAAGACGC-3′; Bcl2 forward: 5′-GACAAGATCGCCCTGTGGAT-3′ and reverse: 5′-AGGGCCAAACTGAGCAGAGT-3′; GST forward: 5′-TTCCTGTGGCATAATGTGAT-3′ and reverse: 5′-CTGATTCAAAGGCAAATCTC-3′. Target gene expression was calculated using the 2-△△ct method. 14
Western Blot Analysis
T24 and T24/DDP cells treated with fMLF (Sigma-Aldrich) were collected and washed with PBS 3 times when they were in exponential growth. Total proteins were lysed by RIPA buffer with 1 mM PMSF (Dingguo Biotech) and 1 mM phosphatase inhibitors (Dingguo Biotech). Proteins were separated on 8% to 12% SDS-polyacrylamide gels and were later transferred to PVDF membranes (Bio-Rad Laboratories, USA) for 50 minutes at 12 V. Tris-buffered saline (Beijing Liuyi Biotechnology, China) with 0.05% Tween-20 (Euroimmun, Germany) and 3% to 5% BSA were used for blocking for 2 hours. Subsequently, the membranes were incubated with specific primary antibodies (Abcam, UK) against FPR1, signal transducer and activator of transcription 3 (STAT3), Janus kinase 2 (JAK2), extracellular regulated protein kinases 1/2 (ERK1/2), phosphatidylinositol 3-kinase (PI3K), protein kinase B (AKT), phosphor-STAT3 (p-STAT3), phosphor-ERK (p-ERK), and β-actin (ZhongShan Co, China) at 4°C overnight. The PVDF membranes were washed 3 times with PBST and the membranes were subsequently incubated with goat anti-mouse or goat anti-rabbit IgG antibodies for 1.5 hours. The bands were revealed by the enhanced HRP-DAB chromogenic kit. The Image-Pro Plus v 6.0 software (Media Cybernetics, USA) was used to analyze the bands’ integrated optical density value. β-actin was used as the internal reference.
Cell Viability Assay
The effects of fMLF on the viability of T24/DDP cells were detected using the Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc, Shanghai, China). T24/DDP cells (6 × 103 cells/well) were seeded into a 96-well plate. After overnight incubation, various concentrations of fMLF (0, 10, 100, 1000 nM; Sigma Aldrich), which was dissolved in DMSO (Sigma) at a final concentration <0.1%, were added to the cells. T24/DDP cells cultured with RPMI-1640 were used as a negative control, while RPMI-1640 without cells was treated as the vehicle control. After treatment with fMLF for 24, 48, and 72 hours, 10 µL CCK-8 solution was added into each well. The cells were then incubated for 1 hour at 37°C, and a microplate reader was later used to detect the absorbance (Bio-Rad Laboratories) at a wavelength of 450 nm.
Cell Migration Assay
A line was drawn on the back of the 24-well plate, and the culture-insert 2 well (Ibidi, Germany) was subsequently added into the well with the line. Next, 70 µL of T24/DDP cell suspension (5 × 105 cells/mL) was added to each well of the culture insert, and 200 µL RPMI-1640 medium was added to the outside of the culture insert. Twenty-four hours later, the culture insert was removed using sterile tweezers. The remaining medium was discarded, and the cells were washed 3 times with PBS, and 2 mL 1% FBS RPMI-1640 medium supplemented with 1 µM fMLF was added to each well. T24/DDP cells cultured with 1% FBS RPMI-1640 were used as a negative control. Cells were imaged after incubation for 0, 24, 48, or 72 hours and calculated with ImageJ software.
Cell Cycle Distribution Analysis
T24/DDP cells treated with fMLF and Boc-MLF (Tocris, United Kingdom) for 48 hours were collected and washed with PBS 3 times. Next, the cells (1 × 106 cells/mL) were fixed with precooled 70% alcohol at 4°C overnight. The next day, the cells were washed twice with PBS, and the buffer was removed from the cell pellet. Next, 0.5 mL PI/RNAse solution (Thermo Fisher Scientific, USA) was added to the cells and incubated for 15 to 30 minutes at room temperature in the dark. Flow cytometry (Beckman Coulter CytoFLEX, USA) was used to test the samples and Modfit 4.0 to analyze the statistics.
Statistical Analysis
All the experiments were repeated 3 times. Statistical analysis was prepared using SPSS version 17.0. Two independent sample t tests and 1-way analysis of variance were used to estimate analyses. P < .05 was considered to indicate statistical significance.
Results
Overexpression of FPR1 Is Associated With Resistance of Bladder Cancer
First, in order to explore the differential expression of FPRs in both cell lines, we separately validated at the gene and protein levels. As shown by qRT-PCR analysis, the expression of FPR1 in T24/DDP cells was lower than in T24 cells (Figure 1A). However, FPR2 and FPR3 were all mostly not expressed in both cell lines (the results are not included). However, as shown by Western blot analysis, the expression of FPR1 protein in T24/DDP cells was higher than in T24 cells (Figure 1B and C). Furthermore, after T24/DDP cells were treated with fMLF for 48 hours, qRT-PCR analysis showed that FPR1 promoted the expression of MRP1 and LRP (Figure 2). We conclude that high expression of FPR1 may be associated with drug resistance of T24/DDP bladder cancer cell.
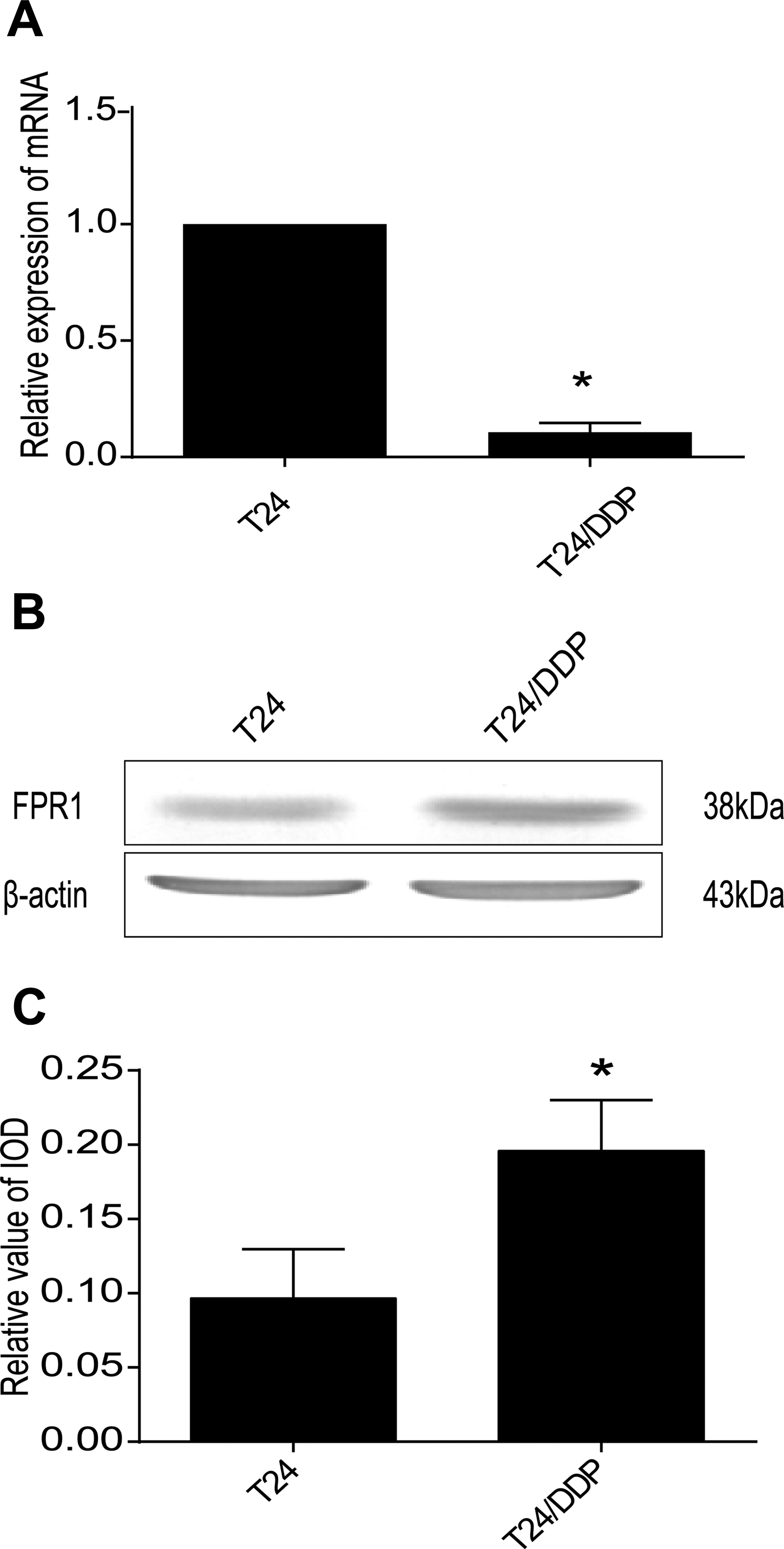
The different expression of FPR1 in T24 and T24/DDP cell lines. A, Quantitative real-time PCR analysis revealed FPR1 expression in 2 cell lines, *P = .01 and <.05. B, Western blot detected a protein band of approximately 38 kDa corresponding to FPR1 in all cells and it is overexpressed in T24/DDP cell line. C, The IOD analysis of Figure 1B, *P = .006 versus control. FPR indicates formyl peptide receptor; IOD, integrated optical density.
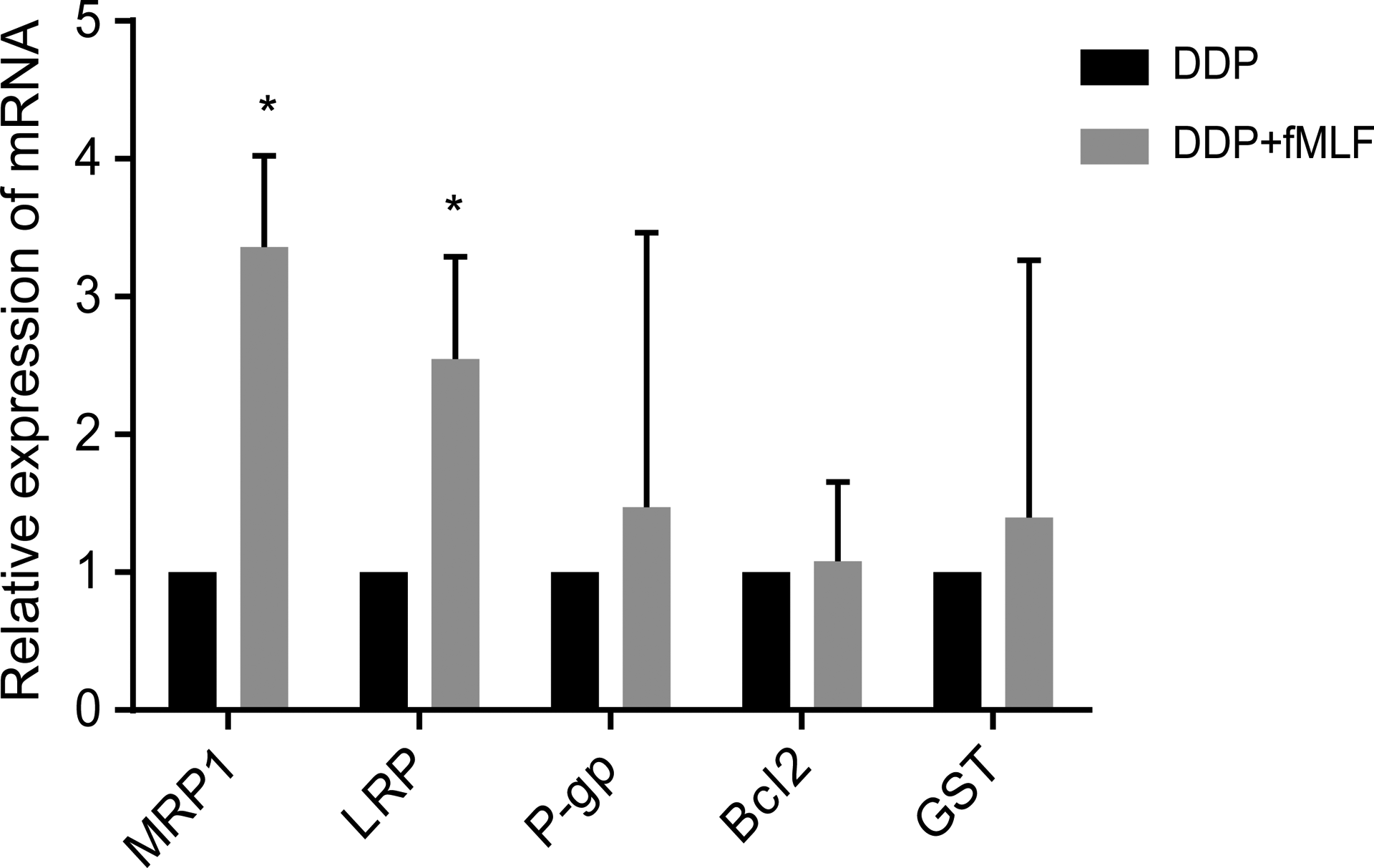
The FPR1 activation improved the expression of MRP1 and LRP. Quantitative real-time PCR analysis revealed that the expressions of resistance-related proteins messenger RNA (MRP1, LRP) were increased. *P < .05. Values shown are mean (standard deviation, SD) from triplicate representative experiments. FPR indicates formyl peptide receptor.
Activation of FPR1 Activates p-STAT3 and p-ERK
To verify which signal pathway is involved in the FPR1 regulation, we detected the expression of STAT3, JAK2, ERK1/2, PI3K, and AKT in T24 and T24/DDP cell lines by Western blot. The expressions of 4 proteins were upregulated: STAT3, JAK2, ERK, AKT, while the expression of PI3K was found to be downregulated in the T24/DDP cell line (Figure 3).
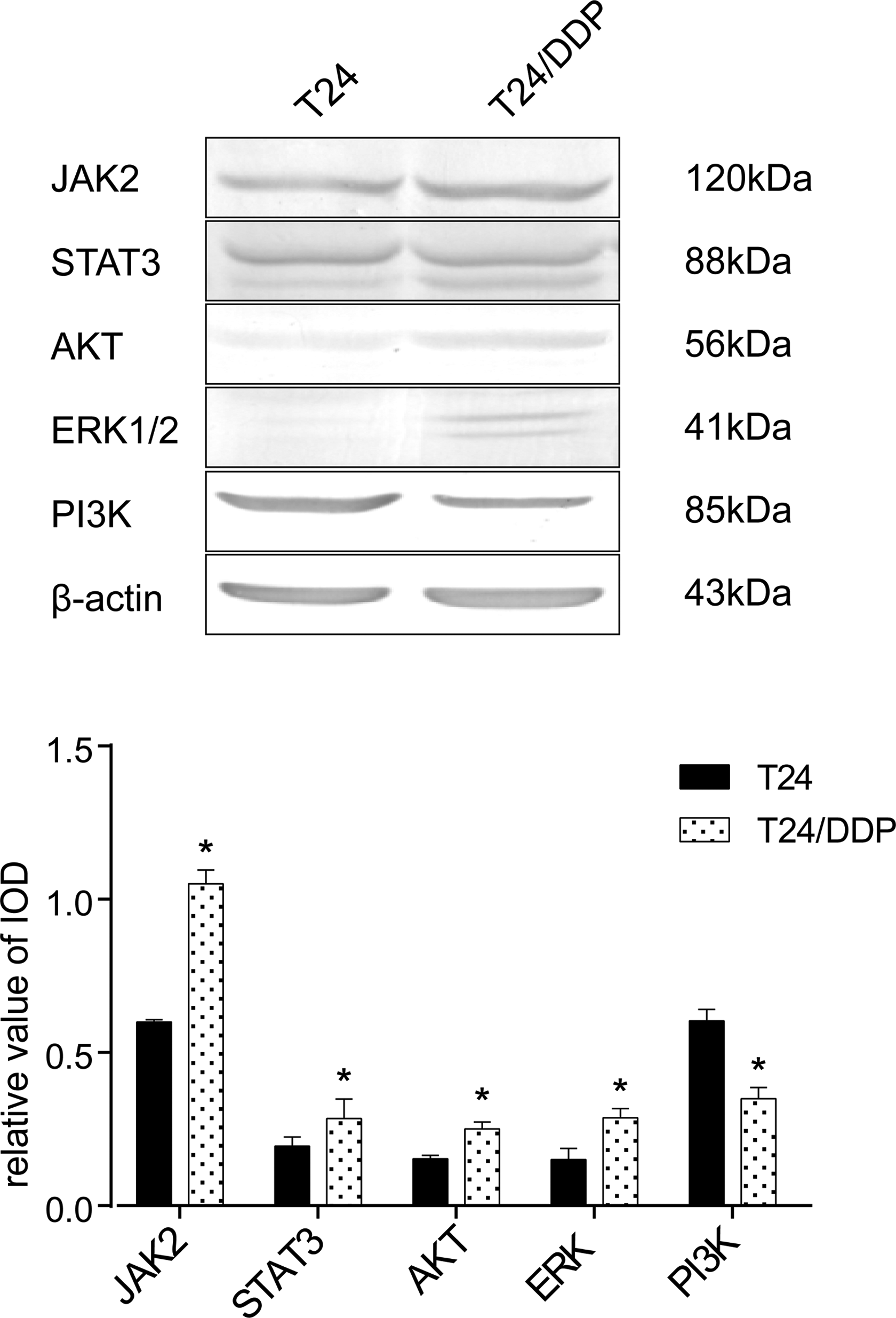
Involvement of JAK2, STAT3, PI3K, AKT, ERK-1/2 in human bladder cancer T24/DDP cells. Western blot detected several signal pathway proteins. All results were analyzed by Image-Pro Plus software for gray values, only PI3K was downregulated and the others upregulated in T24/DDP cells; 120 kDa correspond to JAK2, *P < .01; 88 kDa correspond to STAT3, *P = .042 and <.05; 56 kDa correspond to AKT, *P < .01; 41 kDa correspond to ERK1/2, *P < .01; 85 kDa correspond to PI3K, *P < .01. Values shown are mean (standard deviation, SD) from triplicate representative experiments. AKT indicates protein kinase B; ERK, extracellular regulated protein kinases; JAK2, Janus kinase 2; PI3K, phosphatidylinositol 3-kinase; STAT3, signal transducer and activator of transcription 3.
To investigate the role of FPR1 in drug-resistant bladder cancer, we next stimulated T24/DDP cells with the FPR1 specific agonist, fMLF. T24/DDP treated with fMLF had a significantly higher level of p-STAT3 and phosphor-ERK1/2 expression than normal T24/DDP cells. In addition, with the increase in incubation time and fMLF concentration, the amount of phosphor proteins was also increased (Figures 4 and 5). The results demonstrated that the activation of FPR1 regulated the ERK1/2 and STAT3 signaling pathways.

Activation of FPR1 activates phosphorylated STAT3 and ERK. A, T24/DDP cells were incubated with fMLP (1 µM) for 0, 10, 30, 60, 90 minutes, and cell lysates were analyzed for phosphorylated STAT3 and ERK1/2 by Western blot analyses. B and C, The gray value of the band is analyzed by Image-Pro Plus software and the statistics were analyzed by SPSS 17.0. When cells were incubated for 60 and 90 minutes, *P 60 min (p-STAT3) < .005, P 60 min(p-ERK1/2) < .041, P 90 min(p-STAT3) < .005, P 90 min(p-ERK1/2) < .001, respectively versus control. The data represent the mean (SD) from triplicate measurements. ERK indicates extracellular regulated protein kinases; FPR, formyl peptide receptor; PI3K, phosphatidylinositol 3-kinase; SD, standard deviation; STAT3, signal transducer and activator of transcription 3.
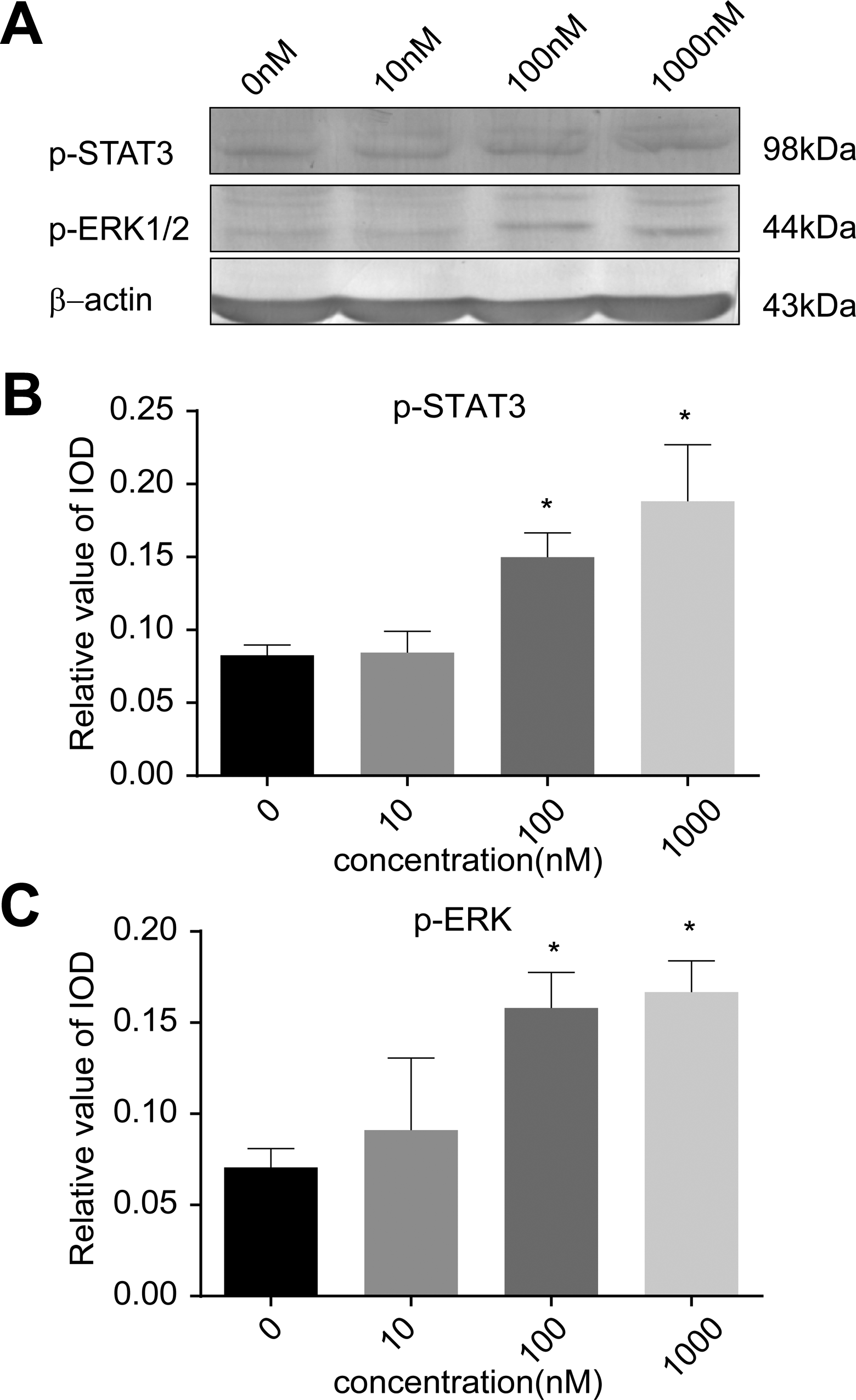
Activation of FPR1 activates phosphorylated STAT3 and ERK. A, T24/DDP cells were incubated with fMLF (0 nM, 10 nM, 100 nM, 1000 nM) for 90 minutes, and cell lysates were analyzed for phosphorylated STAT3 and ERK1/2 by Western blot analyses. B and C, The gray value of the band is analyzed by Image-Pro Plus software and the statistics were analyzed by SPSS 17.0. When cells were incubated with 100 nM and 1000 nM fMLF, *P 100nM (p-STAT3) < .01, *P 1000nM(p-STAT3) < .01, *P 100nM(p-ERK1/2) < .01, *P 1000nM (p-ERK1/2 ) < .01, respectively versus control. The date represent the mean (SD) from triplicate measurements. ERK indicates extracellular regulated protein kinases; fMLF, N-Formyl-Met-Leu-Phe; FPR, formyl peptide receptor; SD, standard deviation; STAT3, signal transducer and activator of transcription 3.
FPR1 Promotes Drug-Resistant Bladder Cancer Proliferation
To access the function of FPR1 in drug-resistant bladder cancer, we used the CCK-8 method to detect the proliferation of T24/DDP cells after FPR1 was activated by fMLF (0, 10, 100, 1000 nM). When cells were treated with fMLF for 24, 48, or 72 hours, the vitality of T24/DDP cells was significantly enhanced compared with the control group. After 24 hours of incubation of T24/DDP cells with fMLF at concentrations 10 to 1000 nM, CCK-8 analysis showed the cell survival rates were 102.23%, 109.69%, and 114.52%, respectively. After 48 hours of incubation of T24/DDP cells with fMLF at concentrations 10 to 1000 nM, CCK-8 analysis showed the survival rates were 112.15%, 113.73%, and 123.44%, respectively. After 72 hours of incubation of T24/DDP cells with fMLF at concentrations 10 to 1000 nM, CCK-8 analysis showed the survival rates were 125.07%, 148.54%, 147.70%, respectively (Figure 6). The results demonstrated that FPR1 activation accelerated the proliferation of T24/DDP cells. In addition, the vitality of T24/DDP cells was improved with the increase in the concentration of fMLF.
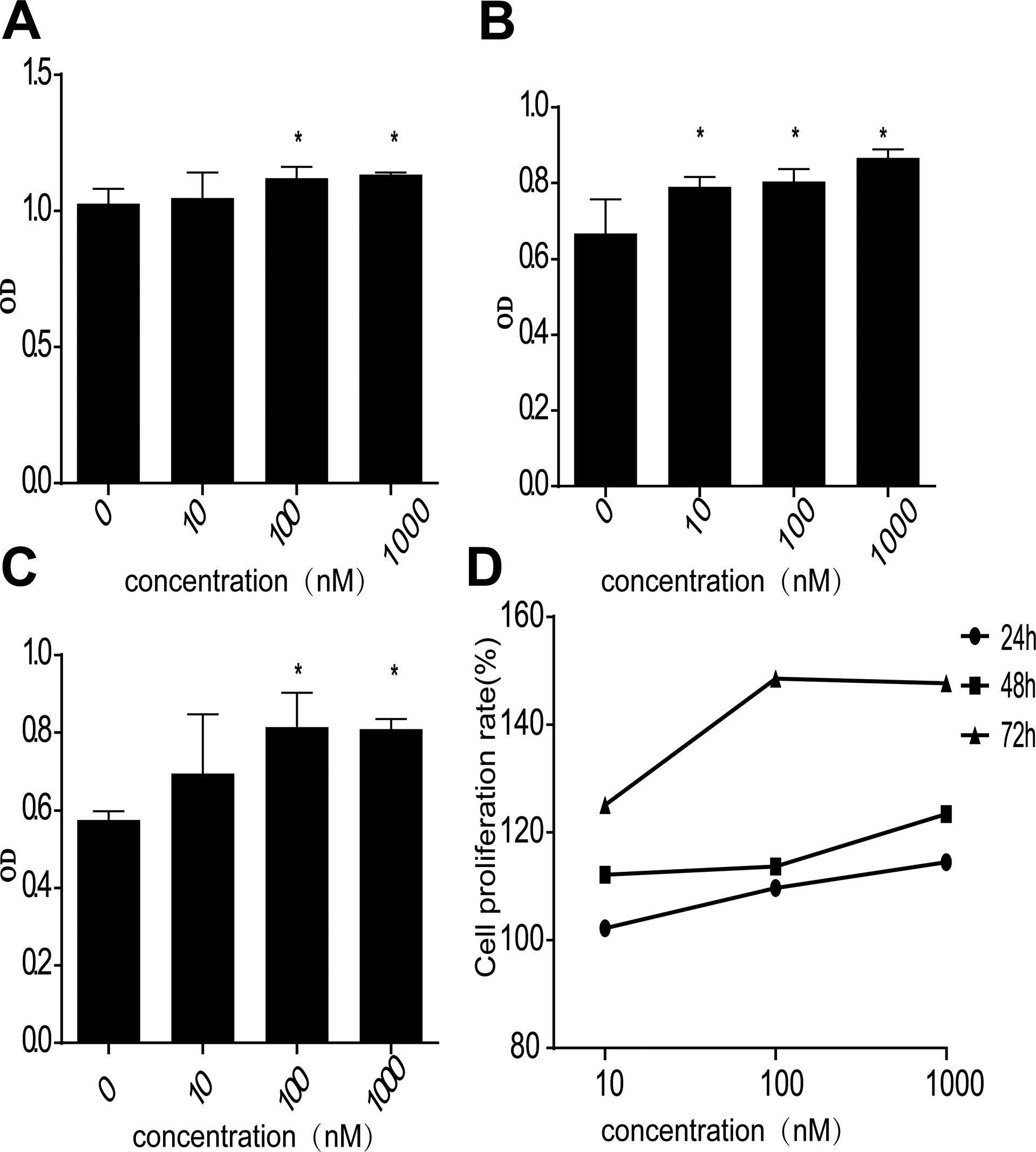
FPR1 promotes the proliferation of drug-resistant bladder cancer. A, T24/DDP cells were treated with fMLP for 24 hours, (B) T24/DDP cells were treated with fMLP for 48 hours, (C) T24/DDP cells were treated with fMLP for 72 hours, OD values were measured at 450 nm by CCK-8, (D) proliferation rate of T24/DDP cells treated with different concentrations of fMLP at different times. Values shown are mean (standard deviation, SD) from triplicate representative experiments. *P < .05. CCK-8 indicates Cell Counting Kit-8; FPR, formyl peptide receptor.
FPR1 Promotes Drug-Resistant Bladder Cancer Migration
To investigate the effect of FPR1 on T24/DDP cell migration, we performed a cell scratch test. The results showed that FPR1 activation improved T24/DDP migration. After T24/DDP cells were treated with fMLF for 24, 48, or 72 hours, the healing rates were 34.76%, 65.69%, and 92.35%, and the control groups were 30.09%, 47.09%, and 60.02%, respectively (Figure 7).

FPR activation promoted the migration of drug-resistant bladder cancer. Use culture-insert 2 well to create the same size of the scratches. Experiment group were cultured in 1% FBS RPMI-1640 medium with 1 µM fMLP, and the control group were just cultured in 1% FBS RPMI medium for 0, 24, 48, 72 hours. Finally calculate the cell healing rate. Values shown are mean (standard deviation, SD) from triplicate representative experiments. *P < .05. FPR indicates formyl peptide receptor.
FPR1 Regulates the Changes in Resistant Bladder Cancer Cell Cycle
To study the effects of FPR1 on the T24/DDP cell cycle, T24/DDP cells were treated with fMLF and Boc-MLF and were later tested by flow cytometry. The T24/DDP cell cycle did not change significantly after activating FPR1 (the results are not shown here), but when FPR1 was inhibited by Boc-MLF, the cell cycle was blocked at the G1 phase (Figure 8).
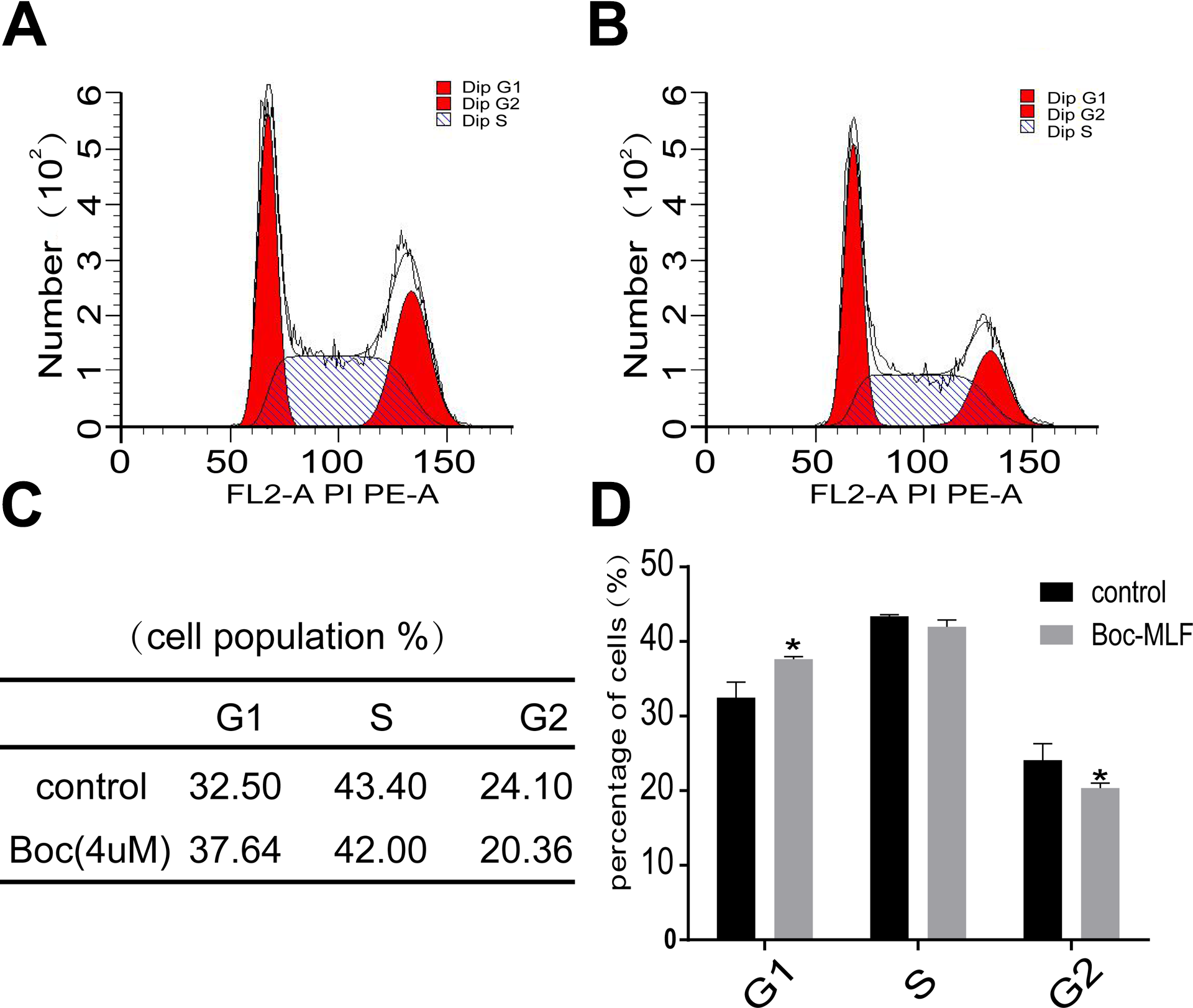
FPR1 regulate the cell cycle of drug-resistant bladder cancer. A, Control group was cultured normally. B, Experiment group was treated with Boc-MLF (4 µM) for 48 hours. C, The percentage of G1, S, and G2 phases in the 2 groups. D, G1 phase was increased and G2 phase was decreased compared to the control group. Values shown are mean (standard deviation, SD) from triplicate representative experiments. *P < .05. FPR indicates formyl peptide receptor.
Discussion
In this report, we demonstrated that drug-resistant bladder cancer cells (T24/DDP) have a higher expression level of FPR1 than normal bladder cancer cells (T24). Moreover, when FPR1 was activated with fMLF, the expression of resistance-related proteins and messenger RNA was increased, and the abilities of T24/DDP cell proliferation and migration were improved. When FPR1 was inhibited by Boc-MLF, the cell cycle was arrested in the G1 phase. Our laboratory has studied the association of Annexin A1 with drug-resistant bladder cancer but did not study its ligand, FPR1. To the best of our knowledge, this study is the only one to investigate the relationship between FPR1 and bladder cancer, especially in a drug-resistant bladder cancer cell line.
At first, FPRs were mainly found expressed in some inflammatory cells and played an important role in immune inflammatory response. 15,16 Because immune escape of the tumor is similar to the inflammatory response, many researchers have focused on the relationship between FPRs and tumors. In recent years, it was verified that FPRs were expressed on the surface of some malignant tumors, and the roles of FPR1 are complex. 17 Several reports have shown that FPR1 was associated with poor prognosis of tumors and had negative effects on the development of tumors, such as gastric cancer, 11 astrocytoma, 18 and melanoma. 19 The loss-of-function mutation of FPR1 has a negative effect on the survival of patients with breast cancer. 20 In contrast to breast cancer, FPR1 activation promoted SW480 cell migration and invasion, and depletion of FPR1 markedly improved the survival rate of the colorectal cancer model mice. 21 Even in the same tumor, the roles of FPR1 are controversial. In gastric cancer, Cheng et al reported that high FPR1 expression was significantly associated with invasion and infiltration of GC and was a risk factor for survival in GC. 11 However, Prevete et al reported that FPR1 could inhibit tumor angiogenesis and may make a positive effect on treating tumors in mouse models. 13,22 In addition, FPR1 is required for the correct positioning of dendritic cells close to dying cancer cells. Also, FPR1 plays an important role in immunotherapy of tumors. 23 To the best of our knowledge, only one article showed the relationship between the expression of FPR1 and drug-resistant tumors. That study verified that FPR1 was overexpressed in drug-resistant pediatric acute lymphoblastic leukemia, 24 but they did not deeply study the mechanism of the specific relationship between FPR1 and drug resistance. In a previous study from our laboratory, we observed that the expression of ANXA1 was associated with bladder cancer resistance and relapse. 25 ANXA1 is an endogenous ligand of FPR1. To date, no studies have shown that FPR1 is linked to bladder cancer, particularly drug-resistant bladder cancer. Our research has decreased the gaps in this field of study. Interestingly, the expression of FPR1 at the gene level was not consistent with that of the protein level. On the premise of ensuring the reliability of the results, we consulted the literature and observed that transcript levels are not sufficient to predict protein levels in many cases, thereby explaining genotype–phenotypic relationships. 26 It is well-known that protein is a manifestation of cell life activity; therefore, we believe that the high expression of FPR1 is related to drug resistance. Different expression of FPR1 in normal bladder cancer and drug-resistant bladder cancer may provide a new ideal and theoretical support for clinical treatment of drug-resistant bladder cancer in the future.
Currently, multidrug resistance of cancers is one of the most difficult issues to treat clinically. Resistance is also the primary reason for the recurrence of bladder cancer after radiotherapy and chemotherapy failure. The mechanism of drug resistance is highly complex. At present, it is believed that the causes of drug resistance include the reduction of drug accumulation and the enhancement of the detoxification ability and antiapoptotic ability of cells. Regulation of signaling pathways is the main factor for multidrug resistance in cancer. In recent years, it was verified that the ERK1/2 signaling pathway is not only associated with the progression of cancer but also participates in the process of cancer resistance. The mechanism may be regulating the expression of the MDR1 gene and protein. Studies have also shown that the ERK/RSK/MEK signaling pathway can improve the expression of P-gp protein. 27,28 The PI3K/AKT signaling pathway is a classical signaling pathway that promotes cell proliferation and inhibits apoptosis. Phosphatidylinositol 3-kinase activation can increase the expression of MDR1 and inhibit apoptosis of tumor cells. 29,30 Activation of the JAK2/STAT3 signaling pathway can result in resistance to CHOP in diffuse large B-cell lymphoma. 31 In addition, blocking STAT3 signaling can enhance cisplatin antitumor activity in ovarian cancer. 32 In our research, although the expression of some signal pathway proteins was significantly different and involved in the drug resistance of bladder cancer, they were not necessarily involved in the regulation of FPR1. However, after FPR1 was activated, STAT3 and ERK1/2 signal pathways were activated, and the migration and proliferation of cells were significantly enhanced. High expression of FPR1 leads to the activation of STAT3 and ERK1/2 signaling pathways and promotes the proliferation and migration of T24/DDP cells; therefore, bladder cancer cells can escape the drugs and finally result in drug resistance.
Factors that affect tumor cell cycle are another important aspect of tumor cell resistance. Our study found that when FPR1 was inhibited by its specific inhibitor, Boc-MLF; the T24/DDP bladder cancer cell cycle was blocked in G1 phase. G1 phase is the preparation period of DNA synthesis. In other words, when lacking FPR1 or inhibiting FPR1 activation, the T24/DDP cell cycle was affected, and the cell proliferation rate slowed. In general, by binding ANXA1, FPR1 induces different pathways and regulates the functions of cells. Yang et al identified that ANXA1 was the chemotactic agonist for FPR1 and accounted for most of the FPR1 agonist activity in vivo. ANXA1 was released by necrotic tumor cells and promoted tumor cell growth and invasion by binding FPR1. 33 In our study, after FPR1 was inhibited by Boc-MLF, and the binding of ANXA1 and FPR1 may be reduced, thereby inhibiting the cell cycle and decreasing cell proliferation.
In summary, FPR1 may activate the JAK2/STAT3 and ERK1/2 signaling pathways to regulate the functions of drug-resistant bladder cancer cells and result in the escape of the cancer. However, our study only involved in vitro studies. The next step would be to collect tissue samples to verify the expression of FPR1 in vivo and establish a cisplatin-resistant bladder cancer mouse model to validate whether our in vitro results are consistent with in vivo experiments.
Footnotes
Abbreviations
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Beijing Natural Science Foundation (7172106) and Beijing Municipal Administration of Hospitals’ Ascent Plan, Code: DFL20150701.