
Abstract
Prostate cancer is the most common solid cancer and genetic factors play important roles in its pathogenesis. XPD is one of the 8 core genes involved in the nucleotide excision repair pathway. The relationship between Asp312Asn, Lys751Gln, and Arg156Arg polymorphisms in XPD and prostate cancer risk is a controversial topic. Therefore, we conducted a meta-analysis to explore the relationship between these 3 polymorphisms and the risk of developing prostate cancer. We searched the electronic literature in PubMed and Google Scholar for all relevant studies (last updated January 1, 2017). The pooled odds ratios and 95% confidence intervals for the associations between the Asp312Asn, Lys751Gln, or Arg156Arg polymorphisms in XPD and prostate cancer risk were calculated. To evaluate the effects of specific study characteristics on the association of these 3 polymorphisms and prostate cancer risk, we performed subgroup analysis if 2 or more studies were available. After an extensive literature review, 7 publications regarding Asp312Asn genotype distribution with 8 case–controls, 9 publications regarding Lys751Gln genotype distribution with 10 case–controls, and 3 publications regarding Arg156Arg genotype distribution with 4 case–controls were selected. The results showed that Asp312Asn (odds ratio = 1.34, 95% confidence interval: 0.96-1.87, P = .000), Lys751Gln (odds ratio = 0.98, 95% confidence interval: 0.89-1.08, P = .986), and Arg156Arg (odds ratio = 1.05, 95% confidence interval: 0.91-1.22, P = .57) polymorphisms do not increase the risk of prostate cancer in the dominant model. Further, in the subgroup analysis by ethnicity, no relationships were observed between Lys751Gln and Arg156Arg polymorphisms and prostate cancer risk. However, stratified analysis by ethnicity revealed that Asp312Asn affects African (odds ratio = 1.57, 95% confidence interval: 1.06-2.33, P = .382) and Asian populations (odds ratio = 2.09, 95% confidence interval: 1.39-3.14, P = .396) in homozygote comparison. In conclusion, this meta-analysis suggests that there is no general association between the Asp312Asn, Lys751Gln, and Arg156Arg polymorphisms in XPD and prostate cancer susceptibility.
Introduction
Prostate cancer (PCa) is the most common solid cancer and the second leading cause of cancer-related deaths in the Western male population. 1 In the United States, there were an estimated 220 800 new cases of PCa and 27 540 deaths from this disease in 2015. 2 In Asian countries, especially China, the incidence rate and mortality rate associated with PCa are increasing rapidly with a change in lifestyle. 3 The etiology of PCa remains largely unknown; however, genetic factors may play important roles in its pathogenesis. 4 Steinberg et al 5 demonstrated that men with 1 first-degree relative and 2 first-degree relatives affected by PCa have 2 times and 5 times higher risk of suffering from the disease, respectively, than that in men with no family history of PCa.
XPD, also known as excision repair cross-complementation group 2 (ERCC2), 6 is one of the 8 core proteins (ERCC1 [excision repair cross-complementation group 1], XPA [xeroderma pigmentosum group A], XPB [xeroderma pigmentosum group B], XPC [xeroderma pigmentosum group C], XPD [xeroderma pigmentosum group D], XPE [xeroderma pigmentosum group E], XPF [xeroderma pigmentosum group F], and XPG [xeroderma pigmentosum group G]) of the nucleotide excision repair (NER) pathway, which repairs DNA damage caused by ionizing radiations and chemotherapy. 7 XPD maps to chromosome 19q13.3 and is composed of 23 exons. 8 Liao et al 9 performed a meta-analysis for PCa to explore the association between Asp312Asn polymorphism in XPD and PCa risk, and their results indicated that this polymorphism is a risk factor for PCa. However, Zhu et al 10 showed that there was no overall association between Asp312Asn and PCa risk. Later, Ma et al 11 showed that the Asp312Asn polymorphism of XPD did contribute to the risk of PCa, but Lys751Gln did not. Similar contrasting results have also been reported about the association between the Lys751Gln polymorphism in XPD and the risk of developing PCa. Mirecka et al 12 showed that the Arg156Arg polymorphism has no relationship with PCa risk. However, the results reported by Zhou et al 13 indicated that Arg156Arg polymorphism increased the susceptibility to high-risk PCa. No meta-analysis has yet been conducted to explore the relationship between the Arg156Arg polymorphism in XPD and the risk of PCa.
Thus, the association between the Asp312Asn, Lys751Gln, and Arg156Arg polymorphisms in XPD and the risk of developing PCa is a controversial topic. Therefore, we conducted a meta-analysis based on current published literature to explore the relationship between these 3 polymorphisms and PCa risk.
Materials and Methods
Search Strategy
A comprehensive literature search was performed to identify publications on the association between the Asp312Asn, Lys751Gln, or Arg156Arg polymorphisms in XPD and PCa risk. We searched the electronic literature in PubMed and Google Scholar for all relevant studies using the following key words: “XPD or xeroderma pigmentosum group D or ERCC2 or excision repair cross-complementation group 2,” “variant or variation or polymorphism,” and “PCa or prostatic neoplasms or prostate cancer” (last updated January 1, 2017). All the available publications were retrieved for evaluating their eligibility. To further identify all the potentially eligible articles for the meta-analysis, the reference lists of reviews and the retrieved articles were searched manually at the same time. Only publications with available full texts were included in the meta-analysis.
Inclusion and Exclusion Criteria
To minimize heterogeneity and to facilitate an appropriate interpretation of the findings, the studies were included for our meta-analysis based on the following criteria: (1) assessment of the association between XPD polymorphisms and PCa risk using a case–control design, (2) the frequency available for each genotype in both cases and controls, and (3) rational test method. The major criteria for study exclusion were (1) reviews, editorials, comments, or animal studies; (2) absence of case–control study; and (3) duplication of previous publications.
Quality Assessment
The Newcastle–Ottawa Scale (NOS) was used to assess the quality of the included studies. 14 The NOS contains 8 items for both cohort and case–control studies. The scale assesses the quality of case–control studies based on 3 areas: selection, comparability, and exposure. A star rating system is used to judge the methodological quality. Selection has a maximum of 4 stars, comparability has a maximum of 2 stars, and exposure has a maximum of 3 stars. The NOS quality scores ranged from 0 to 9 stars, and article scores with 6 or more stars were considered to be qualified. 15
Data Extraction
All the extracted data were stored in the 2013 Microsoft Excel file format. Data were extracted from the included studies as follows: the name of the first author, year of publication, country of the first author, ethnicity, source of control, and the number of cases and controls.
Statistical Analysis
The meta-analysis was performed for XPD polymorphisms that were reported in at least 2 of the included studies. The pooled odds ratios (ORs) and 95% confidence intervals (CIs) for the associations between the Asp312Asn, Lys751Gln, or Arg156Arg polymorphisms in XPD and PCa risk were calculated. To evaluate the effects of specific study characteristics on the association between the 3 polymorphisms and PCa risk, we performed subgroup analysis if 2 or more studies were available. The following genetic models were used: homozygote comparison (Asn/Asn vs Asp/Asp, Gln/Gln vs Lys/Lys, or AA vs CC), heterozygote comparison(Asp/Asn vs Asp/Asp, Lys/Gln vs Lys/Lys, or AC vs CC), recessive model (Asn/Asn vs Asp/Asp+Asp/Asn, Gln/Gln vs [Lys/Lys+Lys/Gln] or AA vs. [CC+AC]), and dominant model (Asn/Asn+Asp/Asn vs Asp/Asp, [Gln/Gln+Lys/Gln] vs Lys/Lys, or [AA+AC] vs CC). Moreover, the I2 statistic was used to determine heterogeneity. I2 values from 0% to 25% indicated low heterogeneity, those between 25% and 50% indicated moderate heterogeneity, and those ≥50% indicated high heterogeneity. 16 The fixed-effects model was used when I2 < 50%; otherwise, the random-effects model was employed. 17 Furthermore, we conducted the Begg test to assess the publication bias, and it was considered statistically significant when P value was less than .05. All the statistical analyses were performed using the STATA 12.0 statistical software package (Stata Corporation, College Station, Texas).
Results
Eligible Studies
A flow diagram of our literature search is shown in Figure 1. Finally, 11 publications were included in our meta-analysis. Table 1 shows the details of these studies, which included 7 publications 12,18 –23 with 8 case–controls that presented Asp312Asn genotype distributions, 9 publications 12,18 –22,24 –26 with 10 case–controls that presented Lys751Gln genotype distributions, and 3 publications 12,13,20 with 4 case–controls that presented Arg156Arg genotype distributions. The quality of studies based on the NOS score is presented in Table 2.

Flowchart showing the selection process for the included studies.
Characteristic of Included Studies.
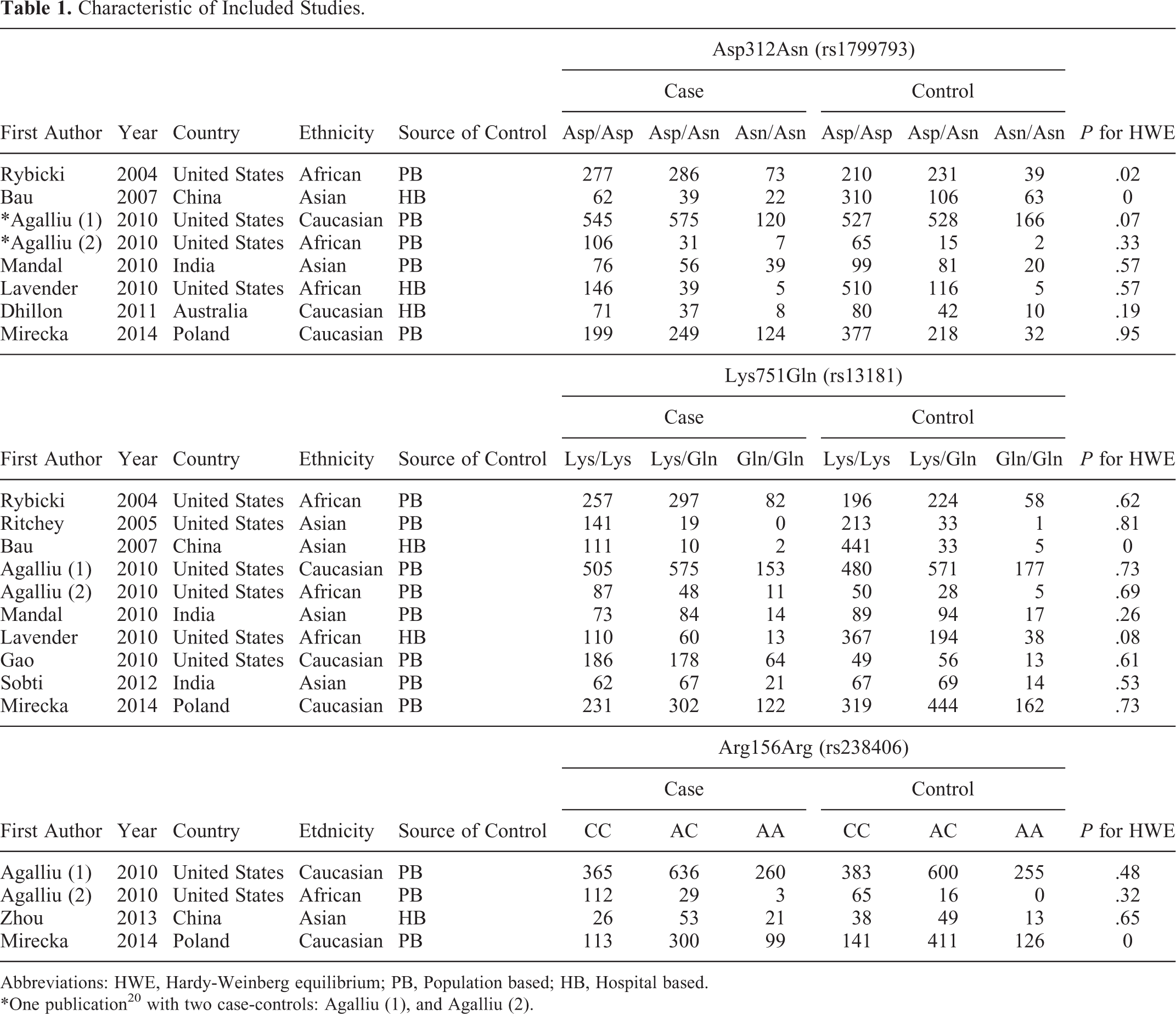
Abbreviations: HWE, Hardy-Weinberg equilibrium; PB, Population based; HB, Hospital based.
*One publication20 with two case-controls: Agalliu (1), and Agalliu (2).
Quality Assessment for All of the Included Studies.

Note. a: three stars; b: two stars; c: four stars; d: one star.
Meta-Analysis
Asp312Asn and PCa risk
For the Asp312Asn polymorphism in XPD, we applied the homozygote and heterozygote comparisons and did not find any increased risk associated with the polymorphism (OR = 1.93, 95% CI: 0.95-3.95, P = .000; OR = 1.24, 95% CI: 0.95-1.60, P = .001). In addition, in the dominant model, we determined an OR of 1.34 (95% CI: 0.96-1.87, P = .000; Figure 2), which indicated that the polymorphism was not associated with an increased PCa risk. Similar results were also obtained with the use of the recessive model: OR was 1.78 (95% CI: 0.94-3.38, P = .000), indicating that the polymorphism was not significantly associated with an increased PCa risk. Subgroup analysis by ethnicity showed no association between the Asp312Asn polymorphism and PCa risk in Caucasian populations in all the genetic models. However, the polymorphism was found to be associated with PCa risk in African populations under all the models and in Asian populations under 3 models (homozygote comparison, dominant model, and recessive model). We also performed subgroup analysis based on the source of controls. In the subgroup analysis for studies with HB controls, an association between Asp312Asn and PCa risk was observed under 3 models (homozygote comparison, heterozygote comparison, and dominant model); no significant association between this polymorphism and PCa risk was found for the PB controls in any of the models. The detailed data are presented in Table 3.

The forest plot of dominant model between Asp312Asn and PCa risk.
Results of Analysis.
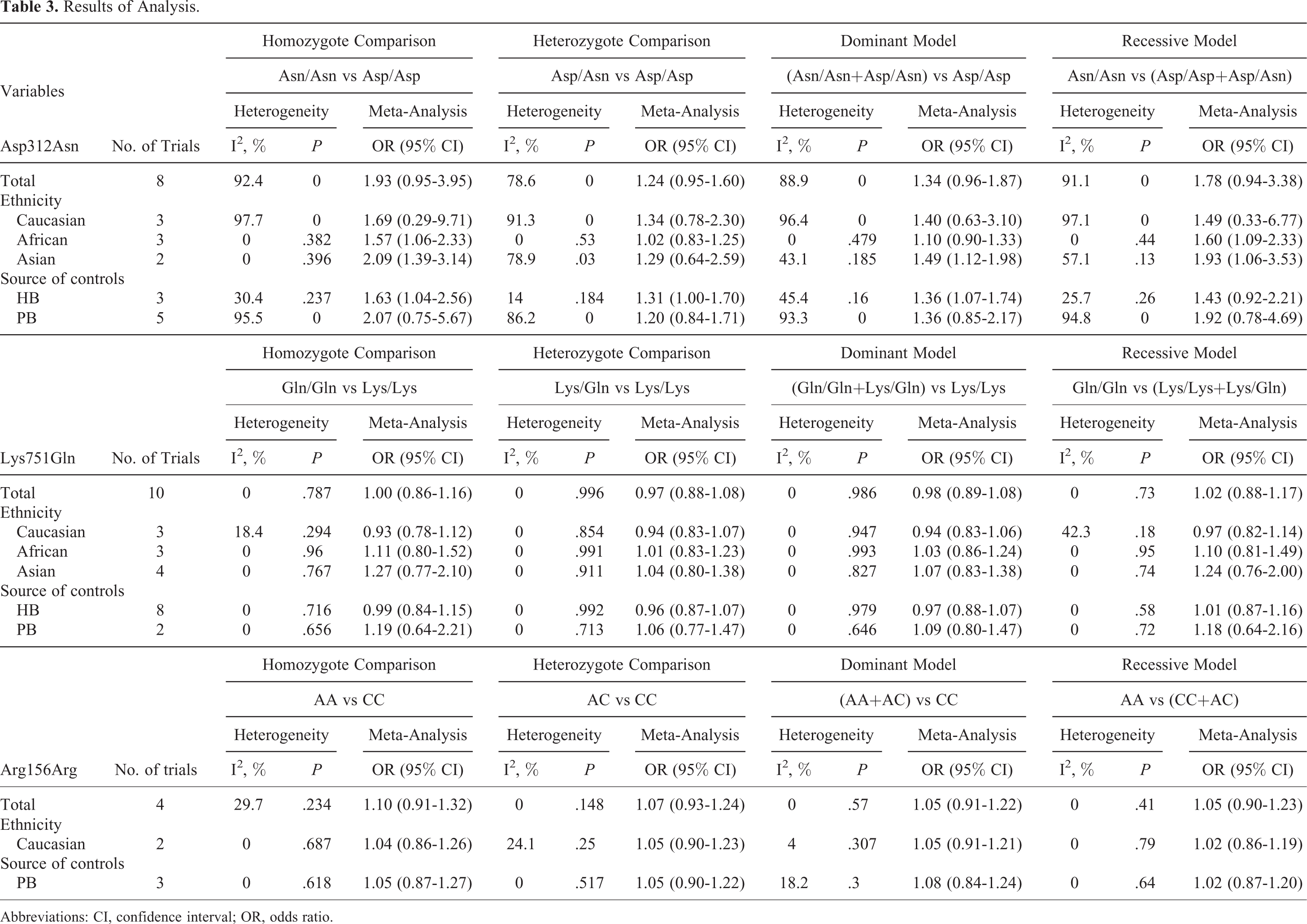
Abbreviations: CI, confidence interval; OR, odds ratio.
Lys751Gln and PCa risk
No significant association between Lys751Gln and PCa susceptibility was found with any of the models (Figure 3). With regard to ethnicity, no significant association between Lys751Gln and PCa risk was detected in Caucasian, African, and Asian populations under all the models. Similarly, when stratified by the source of controls, no relationship was found in the HB or PB group under all the models. The detailed data are presented in Table 3.

The forest plot of dominant model between Lys751Gln and PCa risk.
Arg156Arg and PCa risk
No significant association between the Arg156Arg polymorphism in XPD and PCa susceptibility was observed in any of the 4 models (Figure 4). Moreover, in subgroup analysis by ethnicity, no significant association was observed in the Caucasian population. Subgroup analyses stratified by the source of controls also showed the same result for the PB controls under all the models. The detailed data are presented in Table 3.
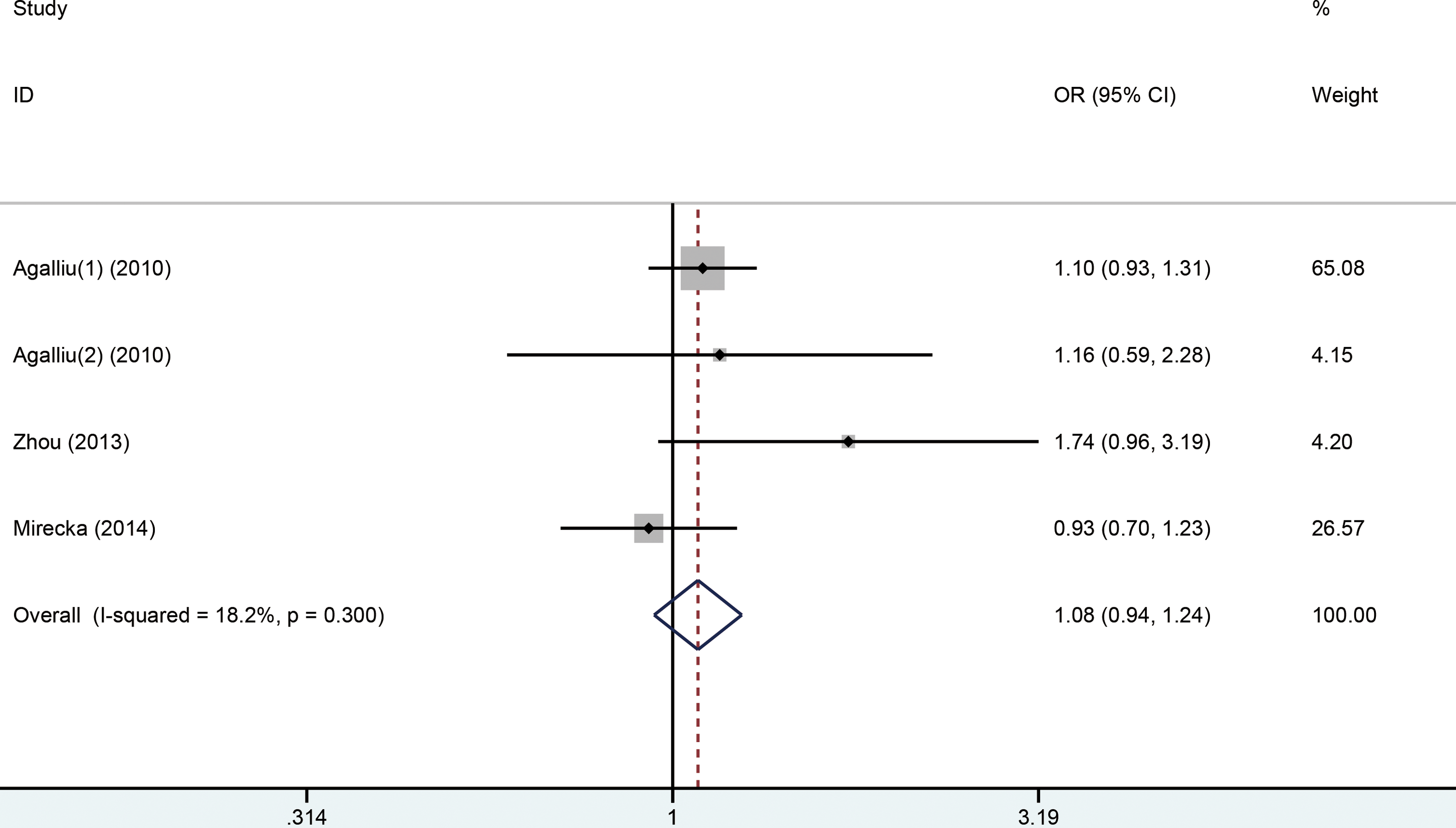
The forest plot of dominant model between Arg156Arg and PCa risk.
Publication Bias
We used Begg funnel plot to assess the publication bias. Our results did not reveal any evidence of obvious asymmetry for the Asp312Asn (P = .373; Figure 5), Lys751Gln (P = .113; Figure 6), and Arg156Arg (P = .219; Figure 7) polymorphisms in homozygote comparisons.

The funnel plot of different model between Asp312Asn and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.

The funnel plot of different model between Lys751Gln and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.

The funnel plot of different model between Arg156Arg and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.
Sensitivity Analysis
A sensitivity analysis was performed to test whether the results of this study were affected by the results of the individual studies and to gauge the stability of the results. We removed studies sequentially from the analysis; however, the result did not change with the removal of any of the studies, suggesting that our results for the Asp312Asn (Figure 8), Lys751Gln (Figure 9), and Arg156Arg (Figure 10) polymorphisms were consistent.
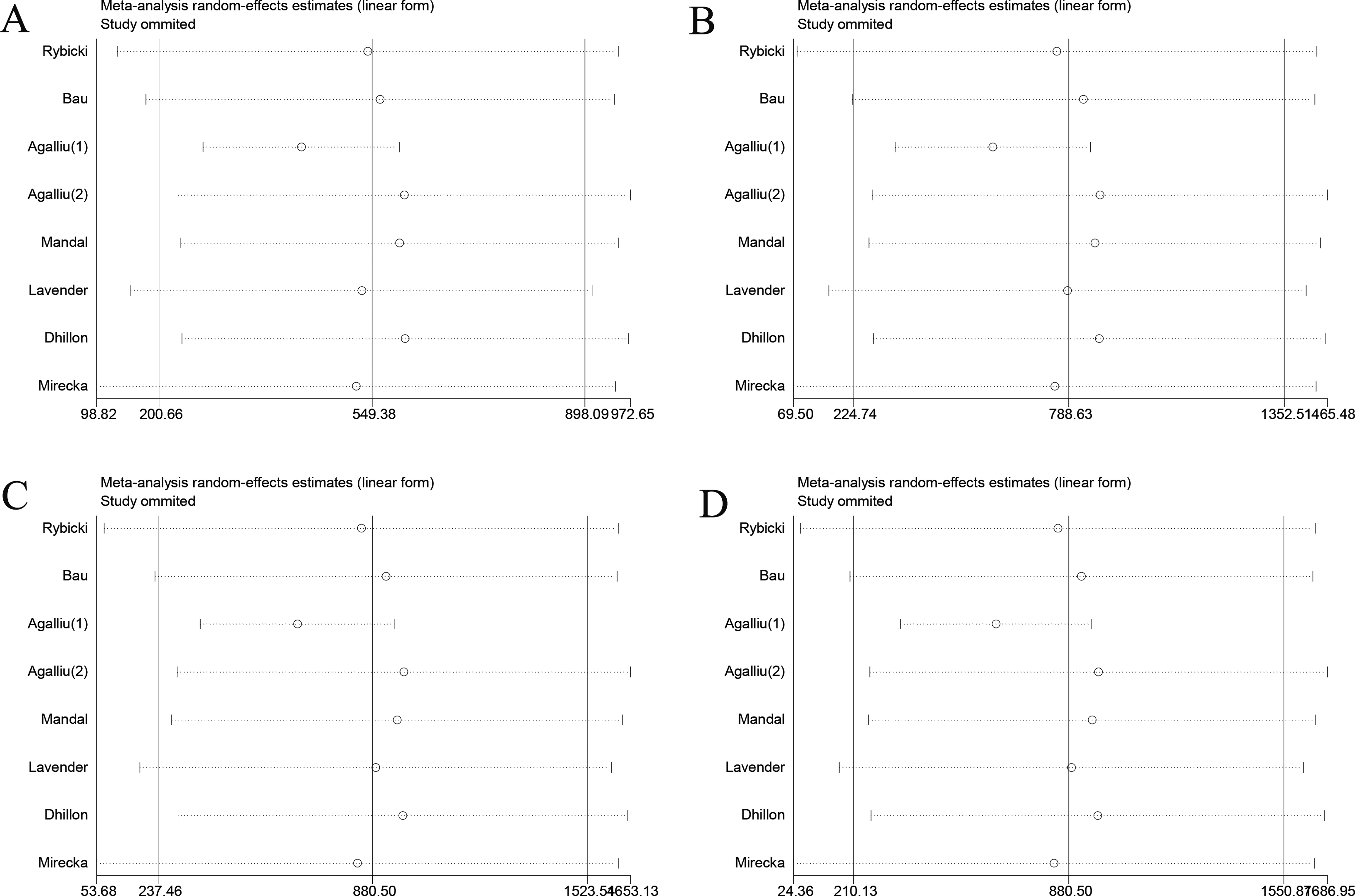
The sensitivity analysis of different model between Asp312Asn and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.

The sensitivity analysis of different model between Lys751Gln and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.

The sensitivity analysis of different model between Arg156Arg and PCa risk. A, Homozygote comparisons. B, Heterozygote comparisons. C, Dominant model. D, Recessive model.
Discussion
The NER pathway is an important DNA repair mechanism that prevents the accumulation of damaged DNA, especially bulky helix-distorting DNA lesions generated from exposure to chemical carcinogens and ionizing radiation, and protects against cancer progression. 27 XPD is a core gene of the NER pathway, and genetic polymorphisms in XPD can cause subtle structural alterations in the repair machinery and modulate susceptibility to cancer. 28 Until now, no genome-wide association study has been conducted for investigating the relationship between XPD and PCa. However, 2 polymorphisms (Asp312Asn and Lys751Gln) in the XPD gene have been largely investigated in genetic epidemiological studies, and 1 other polymorphism (Arg156Arg) has been investigated in a few studies.
In our meta-analysis, the association between the Asp312Asn, Lys751Gln, and Arg156Arg polymorphisms in XPD and PCa risk was evaluated in 7044, 8392, and 4114 participants, respectively. Our results showed that Asp312Asn, Lys751Gln, and Arg156Arg did not increase the risk of PCa. Moreover, in the subgroup analysis by ethnicity, no relationships were observed between Lys751Gln and Arg156Arg polymorphisms and PCa risk. However, stratified analysis by ethnicity showed that the Asp312Asn polymorphism might affect the African and Asian populations. In the subgroup analysis by the source of controls, the Asp312Asn polymorphism was found to possibly contribute to PCa risk in the HB population but not in the PB population; Lys751Gln and Arg156Arg did not contribute to PCa in either population.
Over the decades, many genetic studies and meta-analyses have been performed to investigate the relationship between XPD polymorphisms and the risk of PCa. The most recent meta-analyses showing the association between 2 XPD polymorphisms (Asp312Asn and Lys751Gln) and PCa risk were reported in 2012 and 2013. 9 –11,29,30 A previous meta-analysis 9 included 5 studies (1257 cases and 1956 controls) for the Asp312Asn polymorphism and 6 studies (1451 cases and 2375 controls) for the Lys751Gln polymorphism. Their results indicated that the Asp312Asn polymorphism is a risk factor for PCa development and no significant associations were found between Lys751Gln polymorphism and PCa risk. Zhu et al 29 included 6 studies for Asp312Asn (2714 cases and 3307 controls) and Lys751Gln (3271 cases and 3578 controls) and demonstrated that individuals carrying Asp312Asn were associated with an increased risk of developing PCa. In subgroup analyses, it was found that Asp312Asn polymorphism was associated with PCa risk in Asian populations. Compared to individuals with Lys751Gln, those with Lys751Gln did not possess increased PCa risk, and no association was found in the subgroup analyses. Similar conclusions were derived by Mi et al. 30 Moreover, Zhu et al 10 indicated that Asp312Asn might be a risk factor for developing PCa among Asian and African men, but protective for Caucasian population. The latest meta-analysis conducted in 2013 involved 8 studies including 2620 cases and 3225 controls for the Asp312Asn genotypes and 10 articles involving 3230 cases and 3582 controls for the Lys751Gln genotype. 11 Their results indicated that the Asp312Asn polymorphism contributed to the risk of PCa. Comparatively, our meta-analysis included 1 new study on Asp312Asn and Lys751Gln. The new study 12 included 572 patients with PCa and 627 healthy controls for the Asn312Asp polymorphism and 655 cases and 925 controls for Gln751Lys SNP. In addition, the NOS, which was not conducted in previous 5 meta-analyses, 9 –11,29,30 was used to assess the quality of the included studies in our analysis. Moreover, to the best of our knowledge, this is the first study to analyze the relationship between Arg156Arg polymorphism and PCa risk. We also performed sensitivity analysis, which was not used in previous 2 meta-analyses, 9,29 to evaluate the stability of our study by sequentially removing studies from the analysis.
The Asp312Asn and Lys751Gln polymorphisms in XPD are interesting Single-nucleotide polymorphism (SNP) sites. Previous meta-analyses have shown that the XPD Asp312Asn polymorphism is associated with hepatocellular carcinoma, 31 esophageal cancer, 32 gastric cancer, 33 and bladder cancer, 34 but not with cutaneous basal cell carcinoma, 35 non-Hodgkin lymphoma, 36 breast cancer, 37 or head and neck cancer. 38 In addition, some other meta-analyses have indicated that the XPD Lys751Gln polymorphism is associated with hepatocellular carcinoma, 31 esophageal cancer, 32 breast cancer, 37 and gastric cancer, 33 but not with cutaneous basal cell carcinoma, 35 non-Hodgkin lymphoma, 36 or bladder cancer. 34 Importantly, no meta-analysis has been conducted so far for the XPD Arg156Arg polymorphism. Further, no obvious association between the Asp312Asn and Lys751Gln polymorphisms in XPD and cancer risk has been identified so far. The association between the Asp312Asn and Lys751Gln polymorphisms in XPD and cancer, which is a controversial topic, still needs to be confirmed.
Interestingly, our findings contradict those of some, but not all, previous epidemiological studies that contributed data to the meta-analysis. For example, Agalliu et al 20 indicated a lack of correlation between PCa and the SNPs in DNA repair genes, which is consistent with our findings. However, Bau et al 19 suggested that Asp312Asn may be associated with the development of PCa, which is not consistent with our findings. Considering that a variety of factors may be associated with PCa risk, the contrasting results regarding the association between the Asp312Asn polymorphism and PCa risk in different ethnicities in our meta-analysis are not surprising.
Considering the complex etiology of cancer, the risk of PCa cannot be related to a single factor. Lifestyle, hormonal changes, and gene-wide variations play important roles in the development of PCa. 39,40 Therefore, it is not surprising that Lys751Gln and Arg156Arg do not contribute to the risk of PCa. Contrasting results for this association were also observed in the subgroup analysis by control source; a significant increase in PCa risk was associated with Asp312Asn in the HB population, but not in the PB population. A possible explanation for this difference is that the HB studies may have certain biases for such controls and may only represent samples of an ill-defined reference population, suggesting that they may not be representative of the general population. Alternatively, the numerous participants in the PB control could have been susceptible individuals. 41 However, only 3 studies were performed with the HB population, and therefore, more studies should be conducted in the future to determine this aspect.
Our meta-analysis is not without limitations. First, we only included studies published in English; studies in other languages cannot be disregarded. Second, further analysis based on age, lifestyle, and other factors could not be performed because of insufficient data. Third, the sample size is still small, although we included more studies compared to previously reported meta-analyses. Since meta-analysis is a methodology for pooling the results of multiple studies that evaluate the same general hypothesis, it is based upon the “entirety” of studies. The greater the “entirety,” the greater is the likelihood of identifying the “truth.” Routine differences between studies exist similar to the differences among participants. Therefore, to decrease the differences among studies, the number of included studies must be increased. 42 Thus, more studies on the 3 kinds of XPD polymorphisms should be undertaken in the future with larger sample sizes. Fourth, several studies were not consistent with the Hardy-Weinberg equilibrium, however, exclusion of these studies from each analysis did not change the result. Finally, there was high heterogeneity in our analysis for the Asp312Asn polymorphism; further analysis with more studies on Asp312Asn is required to improve the statistical power of the results.
In conclusion, the results of this meta-analysis suggest that there is no association between the Asp312Asn, Lys751Gln, and Arg156Arg polymorphisms of XPD and PCa susceptibility. However, the Asp312Asn polymorphism might be a risk factor for PCa in the African and Asian populations. More studies on these 3 polymorphisms in XPD should be undertaken in the future.
Footnotes
Abbreviations
Authors’ Note
Weijin Fu, Feifan Xiao, and Ruoheng Zhang contributed equally to this work and should be considered as co-first authors.
Acknowledgments
The authors gratefully acknowledge the support from National Natural Science Foundation of China (Grants number: 81060214, 81160097, 21505025); the Guangxi Natural Science Foundation (Grants number: 2011GXNSFA018175, 2012GXNSFBA053087, 2013GXNSFGA019005).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by National Natural Science Foundation of China (Grants number: 81060214, 81160097, 21505025) and the Guangxi Natural Science Foundation (Grants number: 2011GXNSFA018175, 2012GXNSFBA053087, 2013GXNSFGA019005).