
Abstract
Introduction
Esophageal cancer is one of the most lethal malignancies and ranks as the sixth leading cause of cancer-related deaths worldwide.1,2 More than 70% of esophageal cancer cases occur in China, with esophageal squamous cell carcinoma (ESCC) being the dominant histological type. 3 Currently, the prognosis of patients with ESCC remains poor, with a 10% 5-year survival rate. 4 The lack of robust prognostic and predictive biomarkers is a major impediment to developing a more precise “individualized” approach for treating patients with ESCC.
It has been widely recognized that the MYC oncogene encodes a family of transcription factors that play a significant role in cancer. 5 MYC proteins drive tumorigenesis by inducing comprehensive transcriptional reprogramming, which promotes cell proliferation, metastasis, and genomic instability.5,6
The ability of MYC to regulate transcription relies on its dimerization with MYC-associated factor X (MAX), which results in the formation of a DNA-binding domain that recognizes specific sequences in the regulatory regions of target genes. 7 MAX dimerization protein 1 (MXD1), a member of the MXD family, has been shown to antagonize MYC transcriptional activity by competing for MAX. 8 The increase in MXD1 expression is accompanied by a switch from the MYC–MAX complexes to the MXD–MAX complexes, which block MYC-induced cell growth and transformation, and therefore suppress carcinogenesis. 9
Although the role of MXD1 as a tumor suppressor gene has been established in previous studies,10,11 little is known regarding the clinical and biological significance of MXD1 in ESCC. In the present study, we combined data from Gene Expression Omnibus (GEO) and the Cancer Genome Atlas (TCGA) regarding ESCC to identify the clinical and genomic significance of MXD1 in ESCC, which was further confirmed by immunohistochemistry (IHC) at the protein level.
Materials and Methods
Patients and Data Sources
The set of array-based RNA expression data for ESCC were obtained from human ESCC datasets (GSE23400, GSE20347, GSE29001, GSE38129, GSE26886, and GSE53625), which were downloaded from the GEO website https://www.ncbi.nlm.nih.gov/gds/).
The GSE53625 cohort was derived from a cohort of 179 ESCC patients with follow-up information (minimum of 5 years). 12 All patients had surgically proven primary ESCC and underwent esophagectomy (R0 resection) at the Cancer Institute and Hospital of the Chinese Academy of Medical Sciences between December 2005 and December 2008. All clinical and microarray data of these patients were publicly downloaded from the GEO website (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE53625).
TCGA Cohort
TCGA data were downloaded from the UCSC Cancer Browser website, together with accompanying clinical information. The downloaded RNA-Seq gene expression data were produced using the Illumina HiSeq platform, and then RSEM-normalized and log2 transformed.
Bioinformatic Analysis
SubMap is an unsupervised subclass mapping method that reveals common subtypes between independent datasets. 13 The expression profile of the TCGA ESCC cohort was compared with another published dataset containing 47 patients with melanoma who responded to immunotherapies using the SubMap method. 14 The prediction process was implemented using the SubMap module in the GenePattern platform (https://cloud.genepattern.org/gp/pages/). The Limma R package was used to identify significantly differentially expressed genes (DEGs) between the MXD1-low and MXD1-high groups based on the threshold of fold change and false discovery rate (FDR) or p-value. The intersection of the DEGs was further analyzed by gene ontology (GO) term and Kyoto Encyclopedia of Genesand Genomes (KEGG) enrichment using the R package clusterProfiler. 15 The heat maps of the representative DEGs were generated using the ComplexHeatmap package in R version 3.6.1. 16 The relative proportions of 22 types of tumor-infiltrating immune cells were calculated using the CIBERSORT algorithm. 17
Immunostaining of Tissue Arrays
ESCC tissue arrays (HEsoS180Su08) were obtained from Shanghai Outdo Biotech (Shanghai Biochip Co., Ltd). Rabbit polyclonal antibody anti-human MXD1 (Atlas Antibody, HPA001599) was diluted 1:500 in DAKO antibody diluent. Rabbit monoclonal antibody anti-human programmed cell death 1 ligand (PDL1) (ABCARTA, PA167) and mouse anti-human CD8 monoclonal antibody (ABCARTA, PA067) were used. The EnVision+ detection system (Dako) was used according to the manufacturer's instructions. Immunostained microarrays were scored by multiplying the intensity (0-3) and extent (0-100) of staining for each tissue point, as previously described. 18
Statistical Analyses
Statistical analyses were performed using SPSS (version 23.0; IBM), GraphPad Prism version 8.00 (GraphPad Software), and R version 3.6.1 (R Core Team). Pearson's chi-square test and Fisher's exact test were used to comparing the categorical variables. A Pearson's correlation analysis was used to evaluate the association between 2 continuous variables. Survival analysis was performed using a log-rank test. Univariate and multivariate regression analyses were performed using the Cox proportional hazards regression model to determine the parameters that were significantly correlated with prognosis. A two-sided P-value of <.05 was considered significant.
Results
Prognostic Value and Clinical Relevance of MXD1 in ESCC
A total of 6 public microarray datasets were used to compare MXD1 expression between ESCC tumor tissues and matched normal tissues (Supplemental Table S1). A significant decrease in MXD1 messenger RNA (mRNA) expression was observed in the tumoral tissues (Figure 1A).

MXD1 expression in ESCC and its clinical relevance. (A) Differential expression of MXD1 between tumoral and normal tissue in ESCC. ***P < .001. Decreased expression of MXD1 is associated with the aggressive TNM stage in the GSE53625 (B) and TCGA cohort (C). (D) Survival analysis between MXD1-low and MXD1-high group in the GSE53625 cohort. (E) Representative plot shows MXD1 protein expression was negatively correlated with the TNM stage. (F) Decreased expression of MXD1 protein is associated with the aggressive TNM stage in the validation cohort. (G) Survival analysis between MXD1-low and MXD1-high groups in the validation cohort.
In the GSE53625 cohort, patients with lower MXD1 expression were associated with a more aggressive TNM stage, although the difference did not reach statistical significance (Figure 1B). This finding was further confirmed in the TCGA cohort (Figure 1C). We next assessed the prognostic significance of MXD1 levels in ESCC. In the GSE53625 cohort, the MXD1-low group demonstrated significantly reduced overall survival compared to the MXD1-high group (P = .014; hazard ratio [HR] 1.61, 95% CI 1.10-2.37) (Figure 1D). After adjusting for known risk factors, including age and TNM stage, the multivariate Cox analysis showed that the patients in the MXD1-low group have 1.51 times higher risk of death than those in the MXD1-high group (P = .038, Table 1).
Multivariate analysis by Cox proportional hazards models of overall survival in GSE52625 ESCC cohorts

Abbreviations: MXD1, MYC associated factor X dimerization protein 1; MXD1-H, MXD1-high; MXD1-L, MXD1-low; ESCC, esophageal squamous cell carcinoma.
To confirm the clinical significance of MXD1 in ESCC, MXD1 protein expression was examined using an ESCC tissue microarray (TMA) containing 114 primary ESCC cases, which was utilized as the validation cohort (Figure 1E). The results showed that the expression of MXD1 was negatively correlated with the TNM stage (P = .018, Figure 1F) and overall survival (P = .046) (Figure 1G).
Mutation Profile of Patients with MXD1-low and MXD1-high ESCC
Somatic mutations can play a crucial role in various types of human cancers, whereas some may also act as biomarkers and therapeutic targets. In the TCGA ESCC cohort, we observed that patients with MXD1-low ESCC harbored significantly more nonsynonymous somatic mutations in NFE2L2 (25% vs 7% [P = .037]) and KIAA1324L (12% vs 0.0% [P = .026]) (Figure 2A).

Somatic mutation profile in MXD1-low and MXD1-high group of patients with ESCC. (A) Important somatic mutation differences between the MXD1-low and MXD1-high ESCC in TCGA cohort. (B) Differentially distributed mutation classifications between MXD1-low and MXD1-high groups in the TCGA ESCC cohort.
To obtain a more comprehensive view of the mutation profile, we studied mutational signatures by pathways and gene groups. In the TCGA ESCC cohort, the top 4 most common mutated pathways were the Notch, Wnt, Hippo, and RTK-RAS signaling pathways. MXD1-low patients with ESCC harbored more nonsynonymous somatic mutations in the RTK-RAS signaling pathway than the MXD1-high patients, although no statistical difference was observed (MXD1-low vs MXD1-high: 46.4% vs 27.5% [P = .065]) (Figure 2B). Despite the above-mentioned discrepancies regarding specific mutated genes, MXD1-low tumors presented a similar tumor mutational burden (TMB) as MXD1-high tumors (median TMB 3.17 vs 3.24 mut/Million Basepairs (MB), P = .768).
Transcription Features of MXD1-low and MXD1-high Patients With ESCC
To further elucidate the biological features of MXD1-low and MXD1-high ESCC patients, we performed differential expression analysis between the 2 groups. A total of 191 upregulated and 289 downregulated genes were identified using the criteria of FDR-adjusted P ≤ .05, and |fold change| ≥ 1.0 (Supplemental Table S2).
Gene set variation analysis showed that in the MXD1-low patients with ESCC, gene sets involved in epithelial–mesenchymal transition (EMT) (eg, hallmark_epithelial_mesenchymal) and the extracellular matrix (ECM) (eg, reactome_extracellular_matrix) were repeatedly observed (Figure 3A and B) Furthermore, we found that the expression of a group of key EMT-related genes in ESCC was significantly correlated with MXD1 (Figure 3C to F). Meanwhile, in the MXD1-high group, multiple gene sets were found to be significantly enriched, including the differentiation-related gene sets (eg, reactome_keratinization; bosco_epithelial_differentiation_module). Interestingly, immune-related gene sets (eg, reactome_innate_immune_system) were enriched in the MXD1-high group (Figure 3A and B).

Transcriptomic characteristics of MXD1-low and MXD1-high patients with ESCC. (A) Enriched gene sets in MXD1-low and MXD1-high patients with ESCC. Gene sets that were upregulated in (Left) MXD1-high ESCC patients and (Right) MXD1-low ESCC patients were evaluated. (B) Heatmap according to representative genes expression of the gene sets highlighted in (A). (C) to (F) Gene expression correlation between MXD1 and EMT-related genes.
Potential Impact of MXD1 on Tumor Immune Microenvironment
To further investigate the potential role of MXD1 expression in the tumor immune microenvironment (TIME), we focused on the correlation between MXD1 expression and immune-related gene set, which included 2498 genes from multiple immune-related signatures (eg, antigen processing and presentation; cytokine receptors and cytokines) (Supplemental Table S3).
Differential expression analysis identified 6 upregulated and 26 downregulated genes between the MXD1-low and MXD1-high groups using the FDR-adjusted P ≤ .05. Most DEGs were annotated in the signature of antimicrobials (eg, ANXA6, S100A16, and PGLYRP4), cytokines (eg, CMTM3, TNFSF12, and IL1RN), and cytokine receptors (IL20RB and ANGPTL4) (Figure 4A).
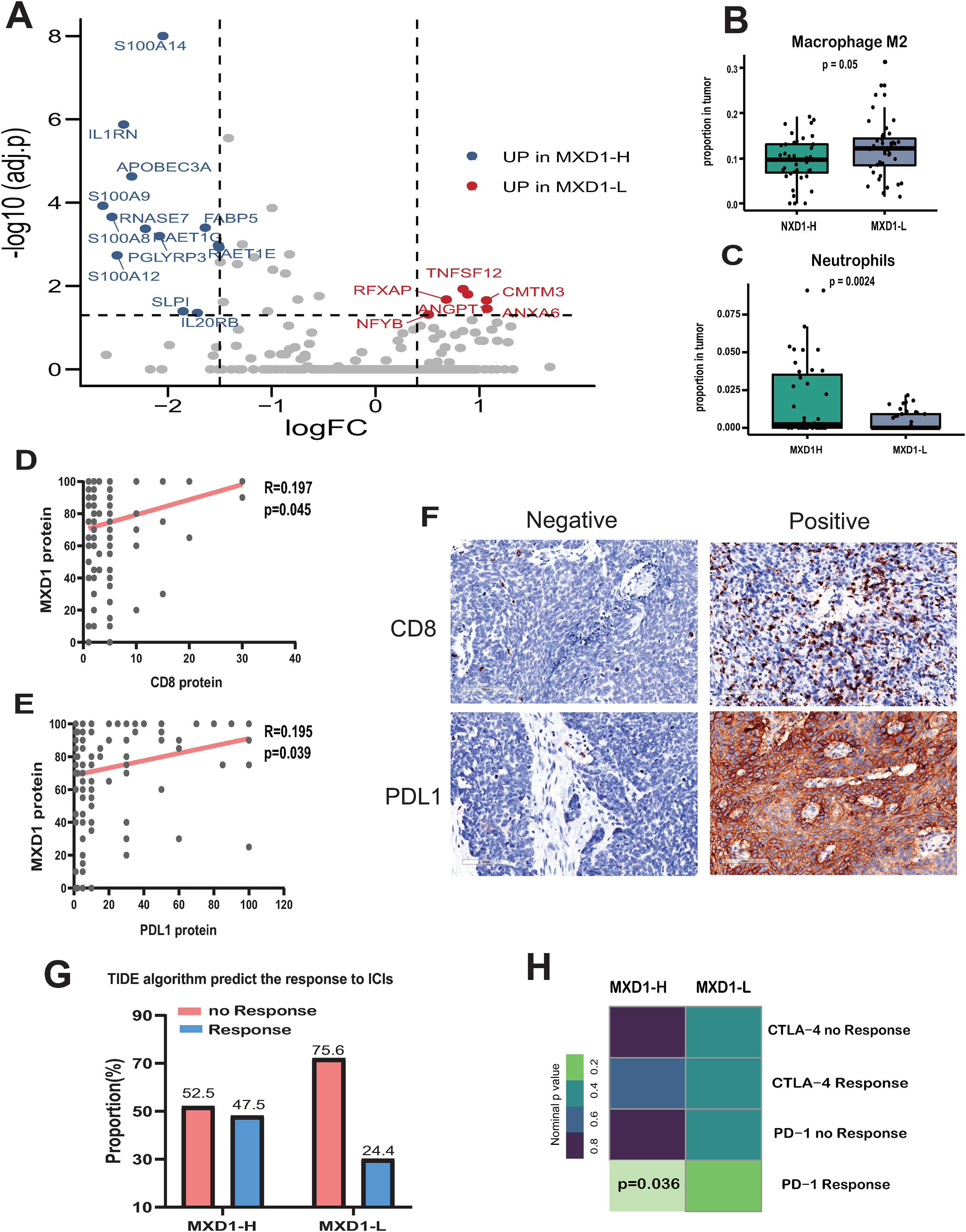
MXD1 and TIME characterization. (A) Differently expressed immune-related genes between MXD1-low and MXD1-high patients with ESCC. Genes that were upregulated in (Left) MXD1-high ESCC patients and (Right) MXD1-low ESCC patients were evaluated. (B, C) The different distribution of immune cell infiltration between MXD1-low and MXD1-high ESCC. Correlations between MXD1 expression and CD8 (D) and PDL1 (E). (F) Representative plots show PDL1 and CD8 protein expression in ESCC tissue microarray. (G) TIDE algorithm predicts the response to ICIs in MXD1-low and MXD1-high groups. (H) SubMap analysis showed that the MXD1-high group was more sensitive to the anti-PD-1 agents.
To gain further insight into the association between MXD1 and the TIME, we calculated the relative proportions of 22 types of tumor-infiltrating immune cells between the 2 groups using the CIBERSORT algorithm. We found that the MXD1-low group contained a significantly lower proportion of neutrophils (P = .002), but a higher proportion of M2 macrophages (P = .050) (Figure 4B and C). These results indicate that aberrant immune infiltration in the TIME might play a role in the aggressive phenotype of MXD1-low ESCC.
To further examine the correlation between MXD1 and the TIME, IHC was used to detect PDL1 and CD8 expression in the ESCC TMA mentioned above. The expression of MXD1 was positively correlated with PDL1 (P = .045) and CD8 (P = .039) (Figure 4D to F).
As the TIME has been proven to have a significant impact on immunotherapy, we investigated the potential effect of MXD1 on immunotherapy sensitivity. We first applied the tumor immune dysfunction and exclusion (TIDE) algorithm to predict the likelihood of response to immune checkpoint inhibitors (ICIs) in the TCGA ESCC cohort. The MXD1-high group (47.5%, 19 of 40) was more likely to respond to ICIs than the MXD1-low group (24.4%, 10 of 41) (P = .038, Figure 4G). In addition, we used the subclass mapping method to compare the expression profile of ESCC with another published dataset including 47 patients with melanoma that responded to ICIs. 14 Similarly, the MXD1-high group was more sensitive to anti-programmed cell death 1 (PD-1) treatment than the MXD1-low group (P = .036, Figure 4H).
Discussion
The prognosis of ESCC remains poor, partly due to the absence of prognostic biomarkers that can identify high-risk patients and thus assign them to risk-adapted monitoring and treatment regimens. In this study, we found that MXD1 was significantly decreased in tumor tissues, which was associated with the aggressive TNM stage and poor prognosis. Therefore, a low MXD1 expression may serve as a biomarker for identifying high-risk patients.
The promotion of terminal differentiation induced by MXD1 activity has been well established.19,20 MXD1 expression has been observed in differentiating post-mitotic cells in the suprabasal layers 21 and in invasive ductal breast carcinomas of a well-differentiated phenotype. 22 These observations are consistent with the clinical relevance of MXD1 observed in our analysis. In addition, the enrichment of tumor differentiation-related gene sets in the MXD1-high ESCC confirmed this association.
Previous studies have reported that MXD1 binds to E-box DNA in the promoters of MYC target genes and represses MYC-mediated transcription, which revealed the competing relationship between MXD1 and MYC for binding to MAX. 23 It is impressive that in the current study, the transcription profile of MXD1-low ESCC was characterized by the upregulation of the EMT and ECM-related gene sets. To date, little is known regarding the link between MXD1 and EMT or the ECM process. It is possible that MXD1 modulates these essential processes in an MYC-dependent fashion as recent studies have demonstrated that activation of MYC signaling can promote the EMT/ECM process.24–26 Further studies are necessary to confirm these observations.
Recently, increased attention has been paid to the crucial role of the TIME in tumor development and progression. 27 The present study showed the potential role of MXD1 in the recruitment of immune and stromal cells in the TIME. MXD1-low ESCC had a higher proportion of M2 macrophages, which are thought to closely resemble tumor-associated macrophages and contribute to the invasion and metastasis of tumor cells.28–30 In addition, the expression of the MXD1 protein was positively correlated with PDL1 and CD8 expression. Lastly, in silico analysis predicted that MXD1-low ESCC had a relatively low sensitivity to immunotherapy. These findings justify further studies to verify MXD1 function and reveal the regulatory mechanism underlying this phenomenon.
The current study had several limitations. First, the robustness of MXD1 as a prognostic indicator in ESCC requires further validation in large or prospective cohort studies. Second, these potential MXD1 novel functions require further validation before they can be used for the clinical management of patients with ESCC. Nevertheless, the results of the current study provide a clearer portrait of the role of MXD1 in ESCC.
Supplemental Material
sj-docx-1-tct-10.1177_15330338211052142 - Supplemental material for MXD1 is a Potential Prognostic Biomarker and Correlated With Specific Molecular Change and Tumor Microenvironment Feature in Esophageal Squamous Cell Carcinoma
Supplemental material, sj-docx-1-tct-10.1177_15330338211052142 for MXD1 is a Potential Prognostic Biomarker and Correlated With Specific Molecular Change and Tumor Microenvironment Feature in Esophageal Squamous Cell Carcinoma by Feng Du, Dezuo Dong, Xiaodong Zhang and Jun Jia in Technology in Cancer Research & Treatment
Supplemental Material
sj-xls-2-tct-10.1177_15330338211052142 - Supplemental material for MXD1 is a Potential Prognostic Biomarker and Correlated With Specific Molecular Change and Tumor Microenvironment Feature in Esophageal Squamous Cell Carcinoma
Supplemental material, sj-xls-2-tct-10.1177_15330338211052142 for MXD1 is a Potential Prognostic Biomarker and Correlated With Specific Molecular Change and Tumor Microenvironment Feature in Esophageal Squamous Cell Carcinoma by Feng Du, Dezuo Dong, Xiaodong Zhang and Jun Jia in Technology in Cancer Research & Treatment
Footnotes
Acknowledgements
The authors would like to thank Yanjie Xiao, Youwu Shi, Jing Sun, Zhiwei Sun, Ying Yang, and Jing Yu for their contributions and gratefully acknowledge the funding support from the sources indicated. The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interests Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors Note
Some or all data, models, or code generated or used during the study are available from the corresponding author by request. Tissue arrays of ESCC (HEsoS180Su08) were obtained from Shanghai Outdo Biotech (Shanghai Biochip Co., Ltd) approved by the National Human Genetic Resources sharing Service Platform (China, 2005DKA21300) for the Medical Research ethical review panel.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by Beijing Excellent Talent Cultivation Subsidy Young Backbone Individual Project; Science Foundation of Peking University Cancer Hospital (A002226) and National Natural Science Youth Science Foundation of China (81902371).
Supplemental Material
Supplemental material for this article is available online.
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.