
Abstract
The dopamine receptors (DRs) family includes 5 members with differences in signal transduction and ligand affinity. Abnormal DRs expression has been correlated multiple tumors with their clinical outcome. Thus, it has been proposed that DRs-targeting drugs—developed for other diseases as schizophrenia or Parkinson’s disease—could be helpful in managing neoplastic diseases. In this review, we discuss the role of DRs and the effects of DRs-targeting in tumor progression and cancer cell biology using multiple high-prevalence neoplasms as examples. The evidence shows that DRs are valid therapeutic targets for certain receptor/disease combinations, but the data are inconclusive or contradictory for others. In either case, further studies are required to define the precise role of DRs in tumor progression and propose better therapeutic strategies for their targeting.
Introduction
The family of dopamine receptors (DRs) includes 5 G protein–coupled receptors—DRD1, DRD2, DRD3, DRD4, and DRD5—with different anatomical distribution, expression levels, and functional properties. 1 For example, DRD1 and DRD2 located in the brain play a role in memory and learning, but peripheral DRD1, DRD2, and DRD4 regulate renal function, blood pressure, and intestinal motility. 2,3 Alterations in DR-signaling have been identified in schizophrenia, Parkinson’s disease, Tourette’s syndrome, and attention deficit hyperactivity disorder. 3,4 Thus, multiple drugs that target the dopaminergic system have been developed. 1,2,5
The DRs are classified into 2 subfamilies based on the G proteins that mediate their signal transduction. 1,6 The D1-like subfamily includes DRD1 and DRD5, which are coupled to Gαs or Gαolf proteins, controlling the activation of the enzyme adenylyl cyclase (AC). The D2-like subfamily comprises DRD2, DRD3, and DRD4, coupled to Gαi/o, govern AC inhibition. Because of their opposite signaling, each subfamily seems to play contrary roles in cellular functions. For example, in neurons, DRD1 stimulation triggers PKA activation, leading to the phosphorylation of cAMP-regulated neuronal phosphoprotein (DARPP-32). 7,8 DARPP-32 phosphorylation is coupled to the de-inhibition of the phosphatase PP1, which dephosphorylate histones, regulating gene expression and the activity of multiple effector proteins such as transcription factors, ionotropic receptors, and ionic channels. 8 On the contrary, activation of D2-like receptors negatively regulates PKA and DARPP-32. 6,9
The interplay between DRs is further complicated by the differences in ligand affinity within the members of the subfamilies. DRD1 binds dopamine with 10-times lower affinity than DRD5 (Ki: 1 µM vs. 100 nM). 10 DRD3 and DRD4 display a similar affinity for dopamine (Ki: 10 nM) which is higher than that of DRD2 (Ki: 100 nM). 10 Furthermore, each DR has different pharmacological properties. 1,10
The idea that DRs could be playing key roles in tumor progression was first supported by the finding that cancer patients that consumed antipsychotic drugs simultaneously to the antineoplastic treatment displayed better clinical responses. 5,11 Further studies showed that schizophrenic or Parkinson’s disease patients who receive ziprasidone, asenapine, quetiapine, clozapine, or aripiprazole (agonists or non-selective antagonist of DRs), 11 or levodopa (a prodrug that yields a non-selective agonism of DRs) 12 have a lower risk of developing diverse types of cancer compared to the general population. 13,14 However, female schizophrenic patients treated with DRD2 antagonists of variable selectivity (haloperidol, risperidone, paliperidone, or amisulpride) have a higher risk of developing breast cancer. 15,16
To date, multiple studies have shown that DRs are differentially expressed in several tumors and that each tumor type has a specific pattern of DRs expression. For example, DRD4 is overexpressed only in Acute Myeloid Leukemia (AML), and DRD3 is expressed only in glioblastoma but at lower levels than in normal brain (Figure 1). Alterations in the expression of DRs have been reported not only in cancer cells but also in tumor-associated cells. Furthermore, forced changes in DRs expression alters diverse functions of cancer cells, indicating that modulation of the receptors’ activity impacts tumor biology (Table 1). Thus, DRs have been pointed as potential therapeutic targets to improve clinical response in cancer patients. 2,5,11,14

Dopamine receptors (DRs) gene expression in tumors. A, mRNA expression of DRs in different tumors (red boxes) and their corresponding normal tissues (black boxes). Statistical analysis was performed using Welch’s t-test. B, Comparison of the expression of DRs in brain tumors vs. normal tissue. Plots were generated using the UCSC Xena platform with data from the TCGA TARGET GTEx database.
Reported Effects of Dopamine Receptor-Targeting in Cancer.

However, the possible application of drugs that target DRs in cancer therapy is still unclear. We found that most of the drugs studied have moderate or low specificity, suggesting that the reported biological effects may be (at least partially) caused by the activity of the drugs on other DRs, other GPCRs, or other cellular targets (see Table 2). For example, pimozide, a DRD2 antagonist employed as second-line therapy for Tourette’s Disorder that has shown antineoplastic effects, can also bind to σ receptors, the serotonin receptor 5-HT7, or calmodulin. 33 Despite their reduced specificity, some of the drugs that target DRs have advantages that make them attractive candidates for developing new anti-cancer therapies, like FDA-approval or specific biodistribution patterns.
Selectivity of Dopamine Receptor-Targeting Drugs Employed in Cancer Studies.
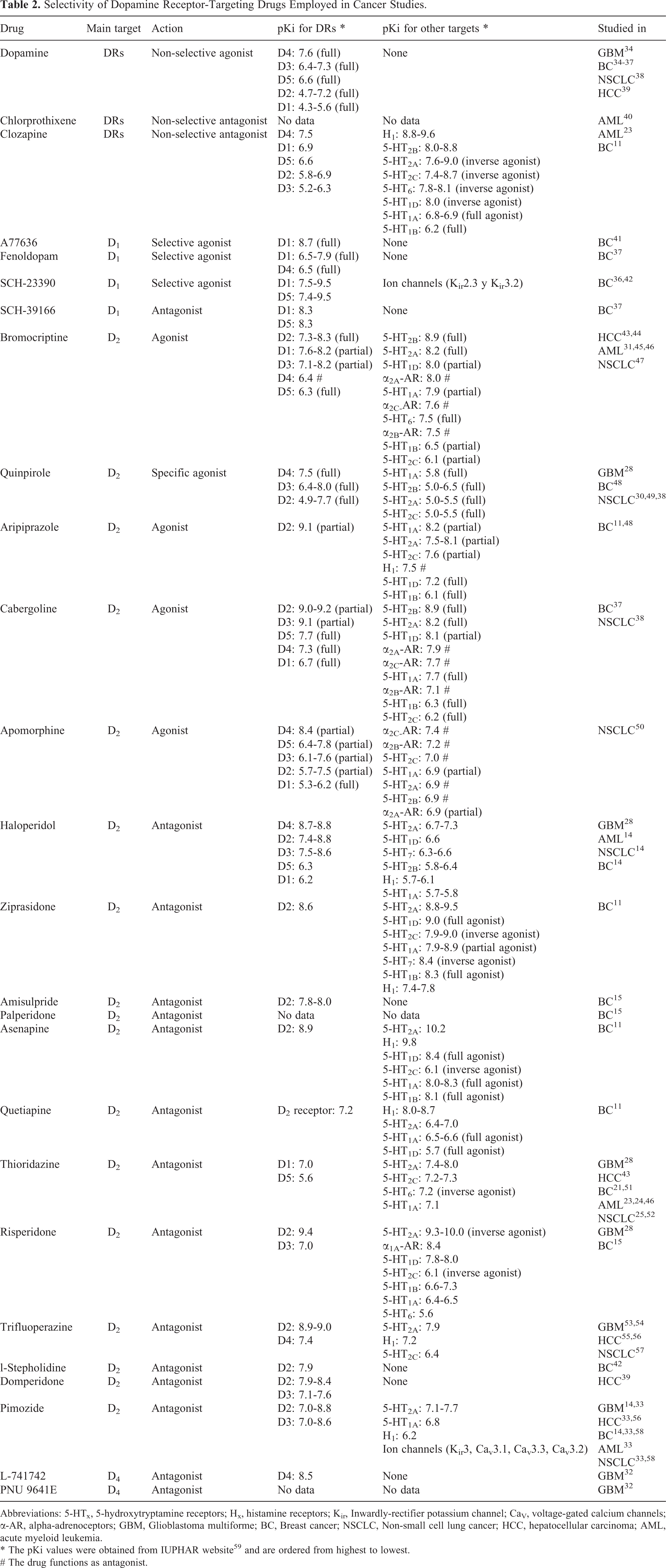
Abbreviations: 5-HTx, 5-hydroxytryptamine receptors; Hx, histamine receptors; Kir, Inwardly-rectifier potassium channel; CaV, voltage-gated calcium channels; α-AR, alpha-adrenoceptors; GBM, Glioblastoma multiforme; BC, Breast cancer; NSCLC, Non-small cell lung cancer; HCC, hepatocellular carcinoma; AML, acute myeloid leukemia.
* The pKi values were obtained from IUPHAR website 59 and are ordered from highest to lowest.
# The drug functions as antagonist.
Herein we compile and discuss evidence of the role of DRs in cancer. We mainly focused on: i) the expression of the DRs in high-prevalence human tumors and its correlation with the clinical outcome; ii) the functions that DRs can modulate in cancer cells; iii) the differences in DRs expression/function between specific subtypes of a particular tumor; and iv) the role of DRs in the subpopulation of cancer stem cells (CSCs).
DRs in Glioblastoma (GBM)
Gliomas represent 80% of the malignant tumors from the brain. GBM, the most frequent and aggressive form of glioma, has a 1-year survival rate of 37.4% and a median survival time of 18 months. 60 Analysis performed using gene-expression data from The Cancer Genome Atlas (TCGA) showed that increased expression of DRs and enzymes participating in dopamine synthesis correlate with GMB patients survival. 34
The 2 members of the D2 family, DRD2 and DRD4, are upregulated in GBM tumors compared to adjacent normal tissue. 32,61 High expression of DRD2 or DRD4 is an independent predictor of reduced survival in GBM patients. 32,61 Increased DRD2 expression is found more frequently in primary GBM than in secondary GBM—which presents a significantly better prognosis. 61,62
Mechanistic studies have shown that: i) DRD2 expressed by GBM cells is activated through autocrine signaling by dopamine, and ii) DRD2 activation triggers mitogenic signals and induces phenotypic changes that favor tumor progression. 2,18,28,34 Accordingly, DRD2 silencing inhibits proliferation of patient-derived GBM cells in culture and reduces the growth of xenografts in mice. 28
The importance of D2-like receptors in GBM progression has been corroborated in studies employing the antipsychotic drug trifluoperazine, a DRD2/DRD4 antagonist. In vitro, trifluoperazine reduces the proliferation and motility of GBM cells by a calmodulin-dependent mechanism. 53 Trifluoperazine is also effective in GBM xenografts, where it inhibits tumor growth and reduces the number of metastatic lesions. 53 Similarly, the small molecule ONC201, a competitive antagonist of DRD2 and DRD3, 29,63 induces apoptosis of GBM cells, including those resistant to temozolomide, bevacizumab, or radiation. 29 In mouse models, ONC201 crosses the blood-brain barrier, inhibiting GBM growth and increasing the median survival time when combined with radiotherapy. 29,64 To date, clinical trials using ONC201 have shown signs of efficacy in biomarker-defined recurrent GBM patients, 64,65 as well as in pediatric and adult H3 K27M-mutant glioma. 65 -67 Noteworthy, ONC201 is also in Phase II clinical trials for AML, 68,69 breast cancer, 69,70 colorectal cancer, 69,71 lung cancer, 71 endometrial cancer, 69 -71 and multiple myeloma. 72
The key role of D2-like receptors in GBM progression led to the analysis of their participation in CSCs biology. DRD2 is expressed in glioma stem cells incrementing the malignancy of tumors. 2,62 DRD2 silencing, 62 but also DRD4 silencing 32 or trifluoperazine treatment, 53 reduces the clonogenicity of GBM cells, an in vitro subrogate measurement of the CSC content. ONC201 reduces the self-renewal of glioblastoma CSCs in vitro 64 and inhibits the proliferation of CSCs in 3D neurospheres culture established from freshly isolated human glioblastoma tumors. 29 Similarly, the DRD4-selective antagonists L-741742 and PNU-96415E decrease GBM stem cell viability and block their clonogenicity but do not affect normal neural stem cells. 32 Furthermore, L-741742 and PNU-96415E display a synergic effect when combined with the first-line cytotoxic agent temozolomide. 32
Altogether, the above evidence shows that DRD2 and DRD4 can be considered therapeutic targets in GBM and suggest that DRD2/DRD4 antagonist as adjuvant therapy would be beneficial for GBM patients. The efficacy of ONC21 in clinical trials supports this hypothesis, but other drugs are still to be tested in patients.
On the other hand, the role of the D1-like receptors in GBM is much less clear. GBM cell lines overexpress DRD1, DRD2, and DRD5 in culture. 2,18,34 Thus, it is possible that the overexpression of those receptors promotes aggressive phenotypes. It has been reported that reduced levels of DRD1 and DRD5 mRNAs favor the survival of GBM patients. 34 However, DRD5 activation functions as an anti-oncogenic signal in GBM cells, inducing autophagic cell death. 73 This contradictory evidence shows that there is not enough information to consider that modulation of D1-like receptors would be beneficial for GBM patients and calls for further studies.
DRs in Breast Cancer
Breast cancer is the most frequent cancer type in women worldwide, causing more than 600,000 deaths per year. 74 Expression of DRs is higher in malignant breast tumors than in benign ones, which, in turn, have increased levels of the receptors than normal mammary tissue. 13 Exogenous administration of dopamine (the natural agonist for all DRs) reduces tumor growth and angiogenesis in vivo. 35,36,75 However, it is unclear if those effects are mediated by a subfamily or a specific dopamine receptor. Moreover, studies aimed to elucidate the role of specific receptors in breast cancer have shown contradictory results.
Breast cancer patients with tumoral DRD1 overexpression have reduced overall and recurrence-free survival compared to patients that do not express the receptor. 37 DRD1 is overexpressed in breast cancer cell lines negative for estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2), suggesting that its signaling is important for the biology of triple-negative (TN) cancer cells. 37,41 Accordingly, the compound A77636, a DRD1 selective antagonist, inhibits proliferation and motility, and triggers apoptosis and autophagy in TN breast cancer cells, but lacks effect in normal epithelial cells. A77636 also reduces osteolytic metastasis in vivo, indicating that the D1-mediated dopaminergic signaling is essential for establishing bone metastasis. 41
However, other authors report that DRD1-signaling impairs breast cancer progression. Dopamine or D1-like agonists reduce the viability and promote apoptosis in TN breast cancer cells, but not in the luminal MCF-7 cells. 37 Apoptosis induction is dependent on DRD1 since it is blocked by the DRD1 silencing or by the addition of the antagonist SCH39166. 37 Furthermore, the DRD1 agonist fenoldopam (which has only peripheral effects due to its absence of brain penetration) inhibits the growth of TN xenotransplants by activating the DRD1/cyclic guanosine 3,’ 5’-monophosphate (cGMP)/protein kinase G (PKG) pathway. 37
In luminal breast cancer cells, the use of fenoldopam or l-stepholidine reduces migration and invasion in a DRD1-dependent manner. 42 The same study demonstrated that fenoldopam or l-stepholidine administration to mice isotransplanted with 4T1 breast cancer cells blocks lung metastasis but does not change tumor growth. 42 On the other hand, administration of the DRD1 antagonist SCH-23390 to mice xenotransplanted with doxorubicin-resistant luminal breast cancer cells promotes tumor growth and blocks the synergic inhibitory effect of dopamine and sunitinib (a multi-targeted receptor tyrosine kinase inhibitor) on tumor growth. 36 These results suggest that the role of DRD1 may be different in TN than in luminal breast cancer cells.
Besides, effects elicited by DRD1 activation may vary in different subsets of cancer cells. Dopamine decreases the fraction of cells with the CSC-associated immunophenotype CD44+/CD24- in MCF/Adr cells, 36 whereas the DRD1 antagonist SCH-23390 increases such fraction. In agreement, activation of the DRD1 with fenoldopam reduces the CD44+/CD24−, ALDH+, and mammosphere-forming fractions in 4T1 cells. 42 Thus, it has been suggested that the population of breast cancer stem cells within luminal cell lines is particularly sensitive to the DRD1 activation, an idea supported by the fact that the large majority of CD44+/CD24− cells express DRD1. 36 However, additional studies are still required to fully clarify the role of DRD1 in luminal breast CSCs. Such studies must evaluate the effect of DRD1 activation on transcriptional and functional responses displayed only by breast CSCs, using proper methodologies beyond immunophenotype. 76 Additionally, the role of DRD1 in CSCs from TN or HER2+ tumors is still to be clarified.
DRD2 also plays an important role in breast cancer progression. DRD2 is overexpressed in breast cancer human samples, as well as in cell lines with different ER, PR, and HER2 expression. 13 However, the TN cells displayed the highest DRD2 levels. 13 The inhibition of DRD2 expression with a microRNA (miR-4301) suppresses the proliferation and induces apoptosis in diverse human breast cancer cell lines, suggesting that DRD2 favors breast cancer progression. 13 Yet, the administration of cabergoline, a DRD2 agonist, lacks effect in xenotransplants of TN breast cancer cells. 37
Additional evidence suggests that DRD2 activation is essential for maintaining the CSC pool, especially in TN cell lines. Quinpirole, a DRD2/DRD3 agonist, increases the mammosphere-forming efficiency in SUM-149 cells. 21 Congruously, DRD2 silencing or the exposure to different D2-like antagonists, including thioridazine, inhibits mammosphere formation by impairing activation of STAT3/IL-6 pathway. 21 Interestingly, thioridazine, an antipsychotic drug employed before 2005, is active against leukemia 23 and colorectal cancer stem cells, 77 suggesting that DRD2 may play similar roles in CSC maintenance in other tumor types. Thus, multiple authors have proposed the use of thioridazine or other DRD2-blocking drugs as adjuvant therapy for breast cancer. Nonetheless, it is unclear if those therapies would equally benefit all different subtypes of breast cancer. For example, in the luminal MCF-7/Adr cells, only 5%-15% of the CD44+/CD24− cells express DRD2, 36 suggesting that only a small fraction of cells within the CSC pool would respond to DRD2 inhibition.
DRs in Lung Cancer
Lung cancer is the most common cancer, and it causes more deaths than any other cancer worldwide. 78 Approximately 85% of the human lung tumors are non-small cell lung cancers (NSCLC); for those patients, the 5-year survival rate is below 20%. 79 Lung tumors express DRD2 and DRD4, 49,80 but DRD2 expression is reduced in NSCLC compared to normal lung tissu. 49 Furthermore, low DRD2 expression correlates with increased risk for larger tumors and more advanced TNM stages. 49 In vitro, DRD2 silencing by shRNA promotes cell proliferation and colony formation, whereas DRD2 overexpression has the opposite effect. 49 In agreement, DRD2-activation by apomorphine 50 or nanoparticled bromocriptine 47 decreases NSCLC cell proliferation. In vivo, activation of DRD2-signaling in cancer cells, either by DRD2 forced overexpression or by the administration of agonists, reduces tumor growth 38,49 and inhibits brain metastasis. 50 Similar effects have been reported in the subpopulation of lung CSCs. DRD2 is expressed in the majority of CD133+ CSCs from NSCLC human tumor samples and human cell lines, and the activation of the receptor reduces their viability, clonogenicity, and invasiveness, as well as in vivo tumor growth. 30
DRD2-signaling can also be triggered in non-cancer tumor cells, affecting NSCLC progression. DRD2 is overexpressed in the endothelium of NSCLC tumors, and such expression correlates with tumor stage and smoking history. 38 Administration of dopamine or DRD2 agonists (quinpirole or cabergoline) impairs the in vivo tumor growth of NSCLC iso- and xenotransplants. 38 Such effect is associated with reduced tumor angiogenesis and decreased infiltration of myeloid-derived suppressor cells (MDSCs). 38 So, it seems that endothelial DRD2 plays an anti-tumoral role in NSCLC.
The evidence above demonstrates that DRD2 activation may be beneficial for NSCLC patients since it reduces proliferation and stemness in cancer cells and may impair the activation of endothelial cells required for angiogenesis.
However, other reports have found favorable effects of DRD2 blockage, particularly with the drug thioridazine. Thioridazine reduces viability and clonogenicity, induces cell cycle arrest, and promotes apoptosis in NSCLC cell lines, and impairs tumor initiation and growth in vivo. 25,52 The drug also increases the sensitivity of NSCLC cells to non-targeted chemotherapy. 25 However, those studies did not analyze whether thioridazine effects are caused only by DRD2 blockage or by additional DRD2-independent mechanisms, as previously suggested. 37 Clarification of the thioridazine mechanism of action will promote the identification of drugs with similar activities.
DRs in Hepatocellular Carcinoma (HCC)
HCC is the most frequent hepatic cancer and is the fourth cause of cancer-related deaths worldwide. 81 HCC patients usually have a poor prognosis, with the exemption of those diagnosed early, for whom the 5-year survival rate is above 70%. 82 Only a few studies have analyzed the role of DRs in HCC progression. Normal hepatic cells and HCC cells express all dopamine receptors. 39 DRD5 is overexpressed in HCC tumor samples compared to normal adjacent tissue, whereas DRD1 expression is decreased. 43 However, the functional roles of such changes are still unclear.
On the other hand, there is enough evidence to support the anti-tumoral role of DRD2 in HCC. DRD2 activation inhibits proliferation and invasion, 39,44 promotes apoptosis, and blocks migration of HCC cells. 44 In vivo, dopamine reduces tumor growth and lung metastasis in a DRD2-dependent fashion since the effects are reverted by the simultaneous administration of dopamine and the DRD2 antagonist domperidone. 39 In agreement, the DRD2 agonist bromocriptine reduces tumor mass and increases overall survival of HHC-bearing mice. 39
As for other tumor types, it has been shown that thioridazine impairs tumorsphere formation and reduces the expression of CSC-associated genes, 43 suggesting that the DRD2 signaling may be different in CSCs and tumor-bulk HCC cells.
DRs in Acute Myeloid Leukemia (AML)
AML is the most common (62%) leukemia and generally has a poor prognosis. 83 It has been reported that human AML cell lines overexpress DRD2 and DRD4 and have low DRD1 and DRD3 expression. 45 Similar results have been found in animal models of myelodysplastic syndrome, 45 a condition associated with an increased risk of developing AML. Chlorprothixene, a wide-spectrum antagonist of DRs, reduces growth and induces apoptosis of AML cells from different subtypes and impairs tumor progression in vivo. 40
The available evidence points to DRD2 as a regulator of cell proliferation and cell death in AML, particularly in leukemia CSCs. Sachlos and collaborators identified that the DRD2 antagonist thioridazine induces differentiation in neoplastic pluripotent stem cells and AML blasts, but lacks effect on normal pluripotent or hematopoietic stem cells. 23 Further analysis showed that thioridazine reduces the engraftment of AML cells but not that of hematopoietic stem cells; such differential effect is caused by the absence of DRs expression in normal cells and the expression of multiple DRs in AML blast. 23 This landmark study showed that: i) leukemic stem cells overexpress DRs, ii) DRs expression in leukemic cells could be a prognosis marker, and iii) altering the dopaminergic signaling could be helpful in a subset of AML patients. 23
Subsequent reports analyzed the efficacy and safety of thioridazine, combined with cytarabine, in patients with recurrent or refractory AML. Oral administration of thioridazine gives rise to plasma concentrations similar to those employed in vitro by Sachlos et al, inducing partial responses in the fraction of patients with DRD2-positive disease at the beginning of the trial. 24 Even when thioridazine causes multiple toxic effects that limit its use, the study demonstrates that DR-mediated signaling is important for AML clinical progression. Nevertheless, as discussed above, thioridazine may not elicit its effects only through DRs. Therefore, these data should not be considered definitive proof that DRD2 (or any other DR) is a universal target in leukemia. For example, the treatment of cultures of AML cells with the DRD2 agonist bromocriptine induces apoptosis, reducing cell viability and colony formation. 45 Likewise, the combined treatment with bromocriptine and cytarabine has a synergic cytotoxic effect in AML cells. 45 Thus, the specific role of DRD2 in AML is still to be clarified. Further studies should analyze the participation of DRD2 using different models of AML with special focus on the CSC population.
DRs in Gastric Cancer
Normal gastric cells and gastric cancer cells express all dopamine receptors. 19 DRD2 is overexpressed in tumor samples compared to adjacent healthy tissues, correlating with a shorter survival of patients. 26 In contrast, gastric cancer cells in culture display reduced levels of DRD2. 19,84 In vitro activation of DRD2 with quinpirole, a specific agonist, inhibits the insulin-like growth factor 1 receptor (IGF-1 R)/Protein kinase B (PKB) pathway reducing cell proliferation. 19 Likewise, exogenous administration of dopamine to tumor-bearing mice inhibits tumor growth by activating DRD2 on endothelial cells, leading to activity suppression in the vascular endothelial growth factor-A receptor-2 (VGEFR2). 84 However, inhibition of DRD2 with thioridazine, a DRD2 antagonist, has also been reported to decrease gastric cancer cell growth. 26
In addition, DRD5 has been associated with cell growth of gastric cancer cells. The activation of DRD5 by the agonist SKF83959 suppresses cell growth by inhibiting mTOR functions and inducing autophagy, followed by cell death. 73 Thus, is it possible that DRD5 become a therapeutic target for gastric cancer in the future.
DRs in Colorectal Cancer (CRC)
In CRC, DRD2 is overexpressed in tumor samples. 18 In vitro, DRD2 activation with dopamine or the D2-like agonist sumanirole increase CRC cell number, whereas transient knockdown of DRD2 or DRD2 inhibition with antagonists (L-741626 or PG01037) generate the opposite effect. 18 ONC201, another selective antagonist of DRD2 and DRD3, also reduces cell viability. However, the effect of ONC201 is not altered by DRD2 knockout, indicating a DRD2-independent mechanism of action. 18
In a subgroup of CRC patients, the existence of DRD2 polymorphisms reduces protein expression 85 and is associated with an increased risk of CRC. 85,86 Furthermore, the dopamine content in malignant human colon tissue is reduced 3- to 10-fold compared to normal tissue, depending on the tumor stage. 85 The progressive reduction of dopamine and DRD2 levels caused by the polymorphisms leads to a reduction of intracellular cyclic AMP, an inhibitor of cell growth, promoting CRC progression. 85,86
DRs in Pancreatic Cancer
In pancreatic ductal adenocarcinoma (PDAC), DRD2 protein levels are increased in tumor samples compared with normal tissue. 22 DRD2 silencing by shRNA inhibits pancreatic cancer cell growth in vitro and in vivo. Similarly, treatment of pancreatic cancer cell lines with pimozide, an FDA-approved DRD2 antagonist, induces cell-cycle arrest and apoptosis. 22 L-741626, another DRD2 antagonist, generates similar effects, suggesting that pharmacologic blockade of DRD2 may be a promising therapeutic strategy for pancreatic cancer. A third DRD2 antagonist, olanzapine, sensitizes pancreatic CSCs to chemotherapeutic agents in vitro, but it is unclear if such effect is mediated exclusively by DRD2 inhibition. 87 Thus, independent studies report that DRD2-targeting could be beneficial for pancreatic cancer patients. On the contrary, no reports analyze the role of other DRs in this tumor type, suggesting that future research should focus on receptors beyond DRD2.
Conclusions and Final Considerations
We conclude that certain DRs can be considered therapeutic targets for specific tumor types. For example, DRD2/DRD4 inhibition in GBM or DRD1 activation in luminal breast cancer produces antitumoral effects. In such cases, future efforts should focus on: i) analyzing the efficacy of specific DR-targeting drugs in relevant models of disease; ii) identifying pertinent pharmacodynamic variables for the evaluation of therapeutic and adverse effects in future clinical trials; and iii) developing new drugs with increased specificity for the target.
On the other hand, the evidence is either inconclusive or contradictory for other receptor/disease combinations. For example, the roles of DRD5 in GBM or DRD1 in TN breast cancer need further clarification. Additional studies should define: i) whether DRs expression is associated with tumor stage and/or clinical outcomes; ii) the differences in DRs expression between tumor-bulk cells and CSCs; and iii) the functional consequences of genetic or pharmacological modulation of DRs in cancer cells. We recommend that when performing those studies, researchers take into consideration that: a) the same receptor can generate opposite responses in different subpopulations of cancer cells; b) the receptor expression levels may be crucial for the induction of specific responses; and c) available DRs-modulating drugs generate off-target effects and, thus, additional controls are required to demonstrate their specificity in the model of study. These considerations will increase the relevance of the data generated and facilitate the development of specific therapies for cancer patients.
Footnotes
Authors’ Note
All authors made substantial contributions to conception and design, compilation of published information, or interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by UNAM-PAPIIT IN219719 (M.A.V-V.) and UNAM-PAPIIT IA205421 (N.S-J.). A.R-C. is recipient of a M.Sc. fellowship from CONACYT (1003227).