
Abstract
Chondrosarcomas are malignant tumors of the cartilage that are chemoresistant and radioresistant to X-rays. This restricts the treatment options essential to surgery. In this study, we investigated the sensitivity of chondrosarcoma to X-rays and C-ions in vitro. The sensitivity of 4 chondrosarcoma cell lines (SW1353, CH2879, OUMS27, and L835) was determined by clonogenic survival assays and cell cycle progression. In addition, biomarkers of DNA damage responses were analyzed in the SW1353 cell line. Chondrosarcoma cells showed a heterogeneous sensitivity toward irradiation. Chondrosarcoma cell lines were more sensitive to C-ions exposure compared to X-rays. Using D10 values, the relative biological effectiveness of C-ions was higher (relative biological effectiveness = 5.5) with cells resistant to X-rays (CH2879) and lower (relative biological effectiveness = 3.7) with sensitive cells (L835). C-ions induced more G2 phase blockage and micronuclei in SW1353 cells as compared to X-rays with the same doses. Persistent unrepaired DNA damage was also higher following C-ions irradiation. These results indicate that chondrosarcoma cell lines displayed a heterogeneous response to conventional radiation treatment; however, treatment with C-ions irradiation was more efficient in killing chondrosarcoma cells, compared to X-rays.
Keywords
Introduction
Chondrosarcomas are defined as malignant tumors of the cartilage accounting for about 10% of all bone tumors with different malignant potential depending on histological grading. 1,2 Since chondrosarcomas are considered to be resistant to conventional chemo- and radiotherapy, 3 the first line of treatment is complete surgical resection of the tumor. Nevertheless, patients with inoperable lesions (eg, due to location) are treated with high-dose radiotherapy (68-74 Gy). 4 -8 However, it is still unclear why the efficacy of conventional radiotherapy is still weak for high-grade chondrosarcoma. Radiotherapy technologies were constantly improved over several decades, with the emergence of intensity-modulated radiotherapy or stereotactic radiotherapy, 9,10 leading to better control of localization and an increased dose to the tumor, while sparing the healthy tissue of risk organs, with promising results, particularly in the treatment of nasopharyngeal carcinoma. 11 Nevertheless, conventional radiotherapy (X-rays/photon) is not the preferred standard therapy for inoperable chondrosarcomas. Indeed, whenever possible, irradiation with accelerated ions is performed. Hadron therapy, using protons and C-ions, allows a single-dose deposit, specifically within the tumor volume, preserving surrounding radiosensitive organs thanks to distribution of the irradiation particles in matter within the Bragg peak. 12 -15 At equivalent physical doses, C-ions present higher biological efficacy against tumor cells than photons (X-rays) and protons. 16 -20 Such efficacy was observed by comparing clonogenic survival of cancer cells irradiated with different radiation qualities. The “relative biological effectiveness” (RBE) can be calculated with the ratio of the dose of each radiation (ions vs photons) inducing 10% of clonogenic survival (D10). The RBE of C-ions was estimated between 1.5 and 7 depending on the cells and culture conditions used and the linear energy transfer (LET) of the native C-ions beam. 16,17,19,21 According to this promising biological efficacy, several studies used C-ions irradiations against different X-ray-resistant cancer cells, in vitro, in preclinical and clinical studies. Delivery of irradiation with this quality requires the development of special infrastructures, comprising large ion accelerators (synchrotron), thus limiting the access to such treatment to a small number of patients and reducing the possibilities of large-scale clinical studies. Nevertheless, several countries are now able to study the benefits of ion radiotherapy applied to X-ray-resistant tumors mostly in Japan, Italy, and Germany. Several installations are planned to be developed in the near future all over the world. A new center dedicated to research with accelerated light ions will be erected in Caen, France (ARCHADE, Advanced Resource Center for HADrontherapy in Europe) and will contribute to the progress of the scientific knowledge on hadronbiology and hadrontherapy. The goal of this study was to analyze the responses of different chondrosarcoma cell lines to C-ions irradiations and estimate the advantage of this irradiation procedure compared to X-rays in targeting malignancy. Different C-ion beams were used for this study, including the C-ions beam from Grand Accelérateur National d’Ions Lourds (GANIL, Caen, France) and from Heavy Ion Medical Accelerator in Chiba (HIMAC) facility of QST (Chiba, Japan). The different technical approaches provided data converging to the same conclusion, namely that C-ions irradiation is more efficient than X-rays at killing chondrosarcoma cells, and demonstrate a high RBE for X-ray-resistant cell lines.
Materials and Methods
Cell Culture
The chondrosarcoma cell line SW1353 (CLS Cell Lines Service GmbH, Eppelheim, Germany) initiated from a primary grade II chondrosarcoma of the right humerus from a 72-year-old female Caucasian was cultured in “chondrocyte growth medium” as previously described 22 using culture medium from CellSystems (#411PR-500; Troisdorf, Germany). The chondrosarcoma cell line CH2879 23,24 was initiated from a primary grade III chondrosarcoma of the chest wall from a 35-year-old female and was cultured in RPMI 1640 (Sigma-Aldrich, Saint-Quentin Fallavier, France), supplemented with 10% fetal calf serum (Sigma-Aldrich). The chondrosarcoma cell line OUMS27 25 was initiated from a primary grade III chondrosarcoma of the humerus from a 65-year-old male and was cultured in minimum essential medium Eagle (MEM, M5650; Sigma-Aldrich), supplemented with 5% fetal calf serum (Sigma-Aldrich). The chondrosarcoma cell line L835 26 was initiated from a primary grade III chondrosarcoma of the humerus from a 55-year-old male and was cultured in MEM (M5650; Sigma-Aldrich), supplemented with 10% fetal calf serum (Sigma-Aldrich). The SW1353 cell line was commercially available; the CH2879, OUMS27, and L835 were obtained from the Department of Pathology, Leiden University Medical Center (Leiden, the Netherlands). All cell lines were cultured in humidified atmosphere with 5% CO2 and cartilage physioxia condition with 2% O2 at 37°C, in a Heracell 150i Tri-Gas incubator. All experiments (passages, etc) were performed outside of the incubator, as fast as possible, to limit the impact of a higher oxygen tension on cell metabolism. To keep 2% O2 during irradiation, flasks without “cap filter” were used and the cap was closed during the short period of irradiation. This study does not report research involving human participants.
Irradiations
X-rays were applied at doses between 1 and 8 Gy. The photon beam was delivered at room temperature (20°C) with a tube tension of 225 kV, a copper filter, and an intensity of 10 mA corresponding to a dose rate of 2 Gy/min on the Pxi XradSmart 225cX irradiator, dedicated to preclinical research, with a lower energy compared to clinical radiotherapy (4-18 MV). The flasks were irradiated horizontally and the medium was changed immediately after irradiation. The dose rate was measured inside flasks with thermoluminescent dosimeters in the irradiation conditions (same distance to the source, same scattering volume below the flask) at 225 kV and 13 mA with a 0.3 mm filtration of copper. Thermoluminescent dosimeters were preliminary calibrated on a 15-cm-thick virtual water phantom thanks to reference dose measurements performed with a calibrated ionization chamber following the “American Association of Physicists in Medicine protocol,” developed by the Radiation Therapy Committee Task Group 61, for reference dosimetry of low- and medium-energy X-rays for radiotherapy and radiobiology. Dose rate was finally corrected from the used tube current.
For C-ions exposure, irradiations were delivered at room temperature (20°C) on 2 different irradiation platforms. Flasks were irradiated vertically, completely filled with medium. The medium was changed immediately after irradiation. Due to the low availability of beam time with such facilities, not all experiments and conditions were performed with all systems.
Clonogenic assays, cell cycle analysis, and Western blotting were performed with the GANIL facility, using the IRABAT beam line. 27 Two LETs were compared to a native 12 C C-ions beam of 95 MeV/A using a PMMA (poly-methyl methacrylate) device inserted between the exit of the beam and the sample holder. Without PMMA (native beam), the LET was 28 keV/µm (2 Gy = 4.46 × 107 particles/cm2); and using a 16.9 mm thickness PMMA (degraded beam), the LET was 73 keV/µm (2 Gy = 1.71 × 107 particles/cm2). In the present study, LET is considered as high when equal or superior to 28 keV/µm.
Micronuclei assays with C-ions were performed at the HIMAC facility. The cells were exposed to C-ions beam at the HIMAC of the National Institute of Radiological Sciences (Chiba, Japan), at room temperature. C-ions were accelerated with an initial energy of 290 MeV/n and the cells were irradiated at the center of a 6 cm spread-out Bragg peak region, with an average LET of 50 keV/µm. In the procedure of dose calibration, the physical dose is measured with a traceable ionization chamber by the standard method authorized by Japan Society of Medical Physics according to the International Atomic Energy Agency (IAEA) protocol. 28,29 The measurement value is converted to the absorbed dose to water and evaluated as Gy/the monitor counts.
Clonogenic Assays
This method is used to screen the sensitivity of cells to irradiation or combined treatment. For this approach, cells at 90% confluency are directly irradiated in 25 cm2 flasks. A sham control is done to evaluate the plating efficiency; this represents the 0 Gy control condition. The medium in the irradiated flasks was changed immediately after irradiation and then the flasks are returned to the incubator for at least 18 hours. Next, the cells are harvested and replated with appropriate dilutions in the multiwell plates. Six-well plates are used with 2 plating densities, in order to obtain about 100 and 1000 colonies per well in control samples. Each well contains 2 mL of medium, necessary for the cellular growth and division. After an incubation period of at least 8 days, the colonies are simultaneously fixed and stained with a crystal violet solution (0.3% wt/vol crystal violet in 20% vol/vol ethanol). Only colonies containing more than 50 cells were counted visually with a stereomicroscope. The results are expressed as a percentage of control unirradiated cells. A linear quadratic model of cell survival is used to fit the results obtained with X-rays according to the equation:
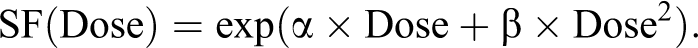
A linear model of cell survival is used to fit the results obtained with C-ions irradiation according to the equation:

All curves were fitted with a dedicated tool for Clonogenic Survival Calculation, the CS-cal software (www.oncoexpress.de), and the corresponding parameters (SF2 = surviving fraction at 2 Gy; D10 = irradiation dose giving 10% survival and D37 = irradiation dose giving 37% survival) were calculated.
Colony Size Distribution
Colony size distributions (CSDs) were determined by allowing single cells to form colonies over the course of 11 days, followed by crystal violet staining, and the number of cells in each colony was counted visually with a stereomicroscope. Colony size distribution data are reported as a cumulative probability distribution of the percentage of cells able to form colonies of greater than or equal to n cells after a defined growth interval. 30,31 Nonparametric Wilcoxon test was used as the data did not meet the assumptions of normal distribution (Supplementary Data 2). The analysis was performed using the computer software R, version 3.1.2 (2014-10-31).
Cell Cycle Analysis
Following irradiation, the cells were plated at subconfluency in 25 cm2 culture flasks, then placed in the incubator from 6 to 48 hours. The cells were incubated with 100 µM of 5-ethynyl-2′-deoxyuridine (EdU) during 2 hours before harvest. The cells were then harvested and centrifuged at 135g for 5 minutes. The cell pellet was washed in phosphate buffered saline (PBS), fixed in ethanol 75%, then stored at 4°C prior analysis. Finally, the cells were centrifuged at 845g for 5 minutes and the cell pellet was resuspended in PBS + BSA (1%) before staining. Fixed cells were stained with Click-iT EdU assay kits for flow cytometry from Thermo Fisher Scientific (Waltham, Massachusetts). Briefly, after permeabilization with 0.1% saponin (15 minutes), the cells were stained with Click-iT reaction mix for 30 minutes (dark room). The cells and all pellets of washing steps were suspended in 500 µL DNA Prep Stain (containing propidium iodide) and 50 µL DNA Prep LPR (DNA Prep Reagent Kit; Beckman Coulter, Pasadena, California). Samples were incubated in the dark for 15 minutes and a minimum of 1 × 104 cells per sample were analyzed using GALLIOS flow cytometer (Beckman Coulter). FlowJo analyzing software (Ashland, Oregon) was used. Experiments were repeated 4 times and data expressed as mean ± standard error of the mean (SEM). A one-way analysis of variance (ANOVA) statistic test was applied for each time point (red for low LET and blue for high LET; *P < .05, **P < .01, and ***P < .001).
Western Blotting Analysis
Following irradiation, cells were detached from the flasks at different time points, centrifuged at 845g for 5 minutes, and the cell pellets were mixed with T-PER lysis buffer supplemented with a protease and phosphatase inhibitors cocktail (ref 78440; Thermo Fisher). This cell lysis step was followed by addition of Laemmli buffer and a denaturation at 100°C. The extracted sample was then separated by SDS-PAGE and transferred to a nitrocellulose membrane according to Hamdi et al. 22 Membranes were analyzed against anti-H2AX phospho-serine 139 (clone JBW301; Merck, Fontenay-sous-Bois, France), anti-GAPDH (MA5-15738; Fisher, Illkirch, France), and anti-p21 (2947; Cell Signaling, Denver, Colorado). Membranes were then incubated with HRP-conjugated secondary antibody (mouse or rabbit; 1:10000; GE Healthcare). The membranes were treated with electrochemiluminescence reagent (Merck KGaA, Darmstadt, Germany) before exposure to hyperfilms (VWR, Fontenay-sous-Bois, France). The films were developed and scanned as JPEGs using a GS 700 Bio-Rad scanner (Bio-Rad, Hercules, CA).
In Vitro Micronucleus Test
The cells were plated on 10-mm-diameter glass coverslips placed in 24-well plates so that they reach subconfluence at the time of analysis. About 22 hours before harvest and 4 hours after irradiation, cytochalasin B (Sigma-Aldrich) was added at a concentration of 3 µg/mL in culture medium. For the analysis of micronuclei, the cells were washed with PBS and fixed in cold acid acetic (10% vol/vol) in methanol solution for 20 minutes. The coverslips were mounted on glass slides with Prolong Gold Anti-Fade reagent with DAPI (Invitrogen, Paris, France) which allowed staining of the DNA. For each experimental point, 500 binucleated cells were analyzed per slide, for at least 3 slides. The micronuclei were scored only in binucleated cells where the 2 nuclei had similar size and staining intensity and did not present nuclear condensation or any other morphology abnormalities. The micronuclei were considered when they were about 1/3 to 1/16 of the size of nucleus and presented similar staining intensity. The experiments were repeated at least 3 times and data expressed as mean ± SEM. A one-way ANOVA test was applied to assess significance at the .05 level.
Results
Clonogenic Survival Is Reduced With C-Ions as Compared to X-Ray Radiation
The clonogenic survival was calculated for the 4 chondrosarcoma cell lines with increasing doses of X-rays or C-ions. Eighteen hours following irradiations at culture confluency, the cells were seeded in culture flasks at low density and the plating efficiencies of SW1353, CH2879, OUMS27, and L835 cells were 0.17 ± 0.02, 0.51 ± 0.08, 0.32 ± 0.05, and 0.34 ± 0.04, respectively. The cells were kept in a humidified incubator at 5% CO2 and 2% O2 for at least 8 days, until large clones could be observed but without cells merging from different clones. Clones with more than 50 cells were counted and survival curves were fitted by Linear-Quadratic (LQ) equation in case of X-rays and linear model in case of C-ions irradiations (Figure 1).

Comparison of clonogenic survival of 4 chondrosarcoma cell lines irradiated with different radiation qualities. The surviving fractions of chondrosarcoma cells irradiated with 225 kV X-rays (blue squares), 28 keV/µm C-ions (red squares), and 73 keV/µm C-ions (green squares). Four chondrosarcoma cell lines were plotted with the same irradiation conditions: (A) (SW1353), (B) (CH2879), (C) (OUMS27), and (D) (L835). The symbols and the bars corresponded respectively to the means and standard errors from at least 3 independent experiments. The data were fitted with the linear quadratic equation in case of X-rays irradiations, and with a linear equation for C-ions irradiations, as explained in the corresponding paragraph of the “Materials and Methods.” The plots were obtained from the CS-cal software, which allowed the calculation of survival and biological effectiveness parameters (Table 1).
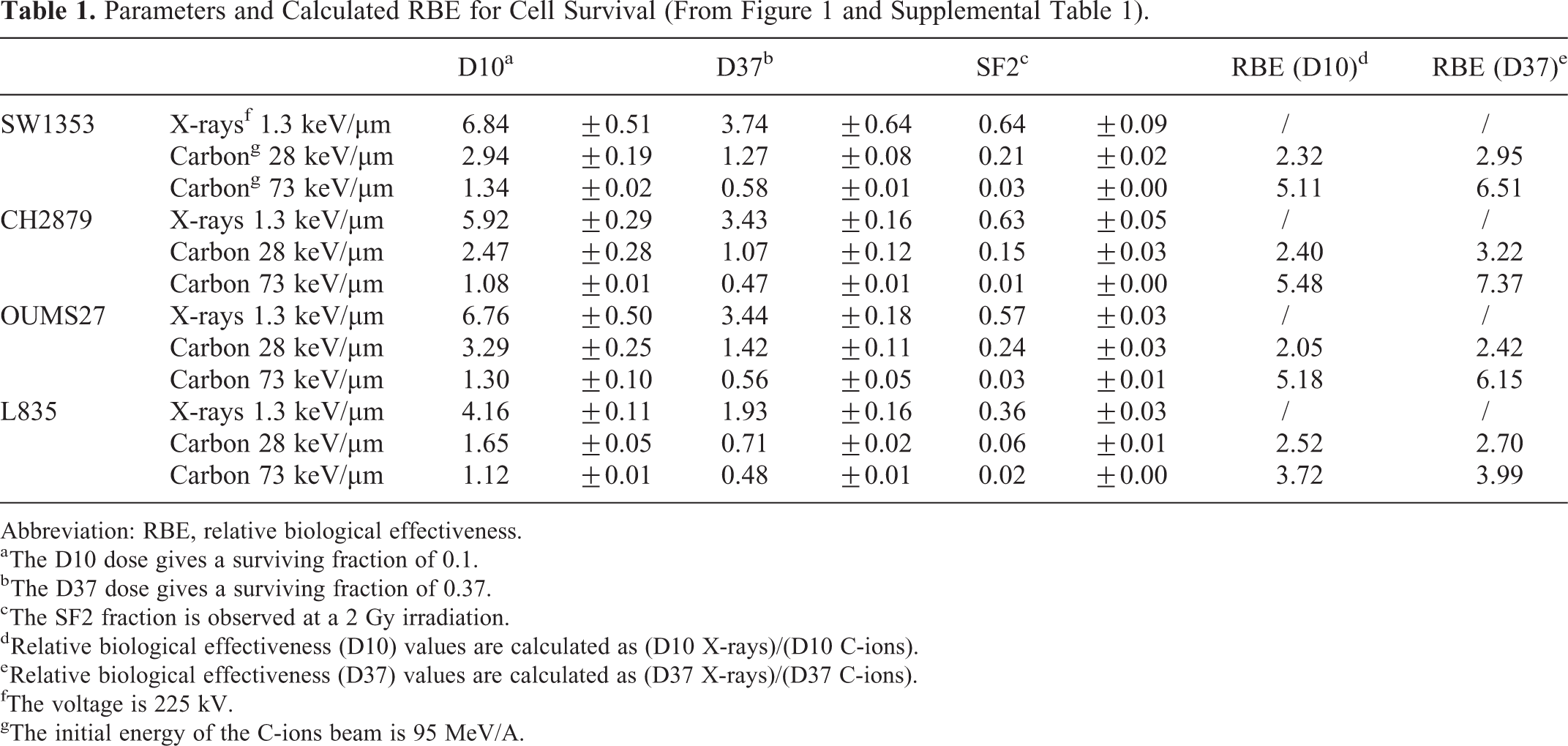
Considering X-rays, the L835 cell line was observed as the most sensitive with a D10 of 4.16 ± 0.11 Gy; and the OUMS27 and SW1353 cell lines were the most resistant with a D10 of about 6.7 Gy (6.76 ± 0.5 and 6.84 ± 0.51, respectively); the CH2879 cell line displayed an intermediate resistance with a D10 of 5.92 ± 0.29 Gy, but not significantly different with the OUMS27 and SW1353 cell lines (Table 1). The same radioresistance was observed when considering the surviving fraction at 2 Gy (SF2). The factors D10, D37, and SF2 illustrated that the 3 cell lines SW1353, CH2879, and OUMS27 presented a similar response to X-rays, while the L835 cell line demonstrated the highest radiosensitivity.
Parameters and Calculated RBE for Cell Survival (From Figure 1 and Supplemental Table 1).
Abbreviation: RBE, relative biological effectiveness.
a The D10 dose gives a surviving fraction of 0.1.
b The D37 dose gives a surviving fraction of 0.37.
c The SF2 fraction is observed at a 2 Gy irradiation.
d Relative biological effectiveness (D10) values are calculated as (D10 X-rays)/(D10 C-ions).
e Relative biological effectiveness (D37) values are calculated as (D37 X-rays)/(D37 C-ions).
fThe voltage is 225 kV.
gThe initial energy of the C-ions beam is 95 MeV/A.
Higher LET irradiations induced a decrease in the surviving fraction, observed for all cell lines, intermediate with C-ions at 28 keV/µm and higher with C-ions at 73 keV/µm (Figure 1). In case of irradiation with C-ions at 28 keV/µm LET, the highest RBE, calculated with the ratio of D10 (X-rays/C-ions), was obtained with CH2879 and L835 cells, ranging between 2.40 and 2.52, respectively (Table 1). The lowest RBE (2.05) was observed with OUMS27 cells; the RBE of the cell lines SW1353 was intermediate (2.32). When considering the RBE with the ratio of D37, the lowest RBE (2.42) was still observed with OUMS27 cells and the highest (3.22) with CH2879 cells, the 2 other cell lines (SW1353 and L835) showed an intermediate RBE of about 2.8 (Table 1). In case of irradiation with C-ions at high LET (73 keV/µm), an increased RBE was observed for all cell lines, but CH2879 cell line displayed the highest RBE (D10) with a factor of 5.48, when compared to X-rays. With the cell lines SW1353 and OUMS27, an RBE higher than 5 was observed (5.11 and 5.18, respectively), and the lowest RBE was obtained with the L835 cell line (3.72), possibly related to the higher sensitivity to X-rays of this last cell line. When considering the SF2 after C-ions irradiation at 73 keV/µm, CH2879 and L835 cell lines were the most sensitive (0.014 ± 0.001 and 0.016 ± 0.001, respectively), and SW1353 and OUMS27 cell lines were the most resistant (0.029 ± 0.001 and 0.030 ± 0.002, respectively).
Each cell line was characterized for the CSD following exposure to irradiation in order to assess the intrinsic differences in growth and division potential of chondrosarcoma cell lines (Figure 2). Both X-ray and C-ion exposures at a single dose of 2 Gy reduced significantly the growth and division potential of chondrosarcoma cell lines. However, the respective impact of X-rays and C-ions was very different from one line to another. Indeed, the SW1353 line showed a light but significant effect of X-rays compared to sham, while C-ions presented a strong impact (Figure 2A). Both CH2879 and OUMS27 cell lines (Figure 2B and C) showed a fair intermediate effect of X-rays on division potential in between sham and C-ions irradiations. Finally, the L835 cell line (Figure 2D) displayed a major impact of both radiation modalities, although a significant difference remains measurable between photons and C-ions.

Colony size distribution of 4 chondrosarcoma cell lines irradiated with different radiation qualities. Four chondrosarcoma cell lines were plotted with the same irradiation conditions: A (SW1353), B (CH2879), C (OUMS27), and D (L835). The symbols corresponded to 3 experiments; blue diamond (control), red square (2 Gy X-rays), and green triangle (2 Gy 28 keV/µm C-ions). Control, X-rays, and C-ions plots were significantly different, excepted in case of the L835 cell line (X-rays vs C-ions), Supplementary Data 2.
The cell line SW1353 displayed an average RBE and radioresistant responses to X-rays and C-ions. The following experiments presented in this study were focused on this cell line, reducing the number of samples to be analyzed, and in relation to the reduced access to C-ions beams to perform these experiments.
C-Ion Irradiation Induced a G2 Blockage and Major DNA Damages
SW1353 cells were analyzed by flow cytometry to estimate the percentage of cells in each cycle phase and the capacity of irradiation to block cells on specific phases. Following 2 Gy X-rays and C-ions at 28 keV/µm, cells were fixed at different time points and analyzed by flow cytometry (Figure 3).

Cell cycle progression of chondrosarcoma cells after exposure to different radiation qualities at a dose of 2 Gy. The cell cycle distribution of SW1353 cells unirradiated (empty circle), irradiated with X-rays (filled circle), and 28 keV/µm C-ions (filled square) is compared between G0/G1 (top panel), S (middle panel), and G2/M phases (bottom panel) at different time points following irradiation (from 6 to 48 hours). Time points significantly different from control were reported with 1 star (P < .05), 2 stars (P < .01), and 3 stars (P < .001), in red in case of X-rays irradiation and in blue in case of C-ions.
According to the (sham) unirradiated control, 22.7% (±0.6) of SW1353 cells were in G2/M phase at 6 hours (Figure 3, open circle, bottom) and 43.9% (±1.3) of cells were in G1 phase with a maximum (54.4% ± 4.3) after 15 hours (Figure 3 open circle, top). When considering X-irradiated cells (Figure 3, filled circle), a significant increase in the G2/M phase population was observed, with a maximum (36.9% ± 2.3) 9 hours after irradiation. The level of this population (Figure 3, filled circle, bottom) went back to the level of the unirradiated control 18 hours after irradiation. At the opposite, the level of the G2/M phase population stayed significantly higher to the sham irradiated control when cells were irradiated with 2 Gy C-ions. For C-ion irradiation, the blockage in G2 reached the maximum after 12 hours (45.9% ± 2.7) and then decreased to a plateau (30.7% ± 5.3) but without reaching the level of the control. Indeed, C-ions induced a longer G2/M phase arrest as compared to X-rays. No sub-G1 fraction could be observed following 2 Gy irradiations (X-rays and C-ions).
In addition, a higher amount of micronuclei was observed following C-ions irradiation, as compared with X-rays (Figure 4). SW1353 cells irradiated with 2 Gy C-ions at 50 keV/µm displayed about 500 micronuclei per 1000 binucleated cells (499 ± 30), while about 375 micronuclei per 1000 binucleated cells (374 ± 20) were induced by 2 Gy X-rays in our conditions.
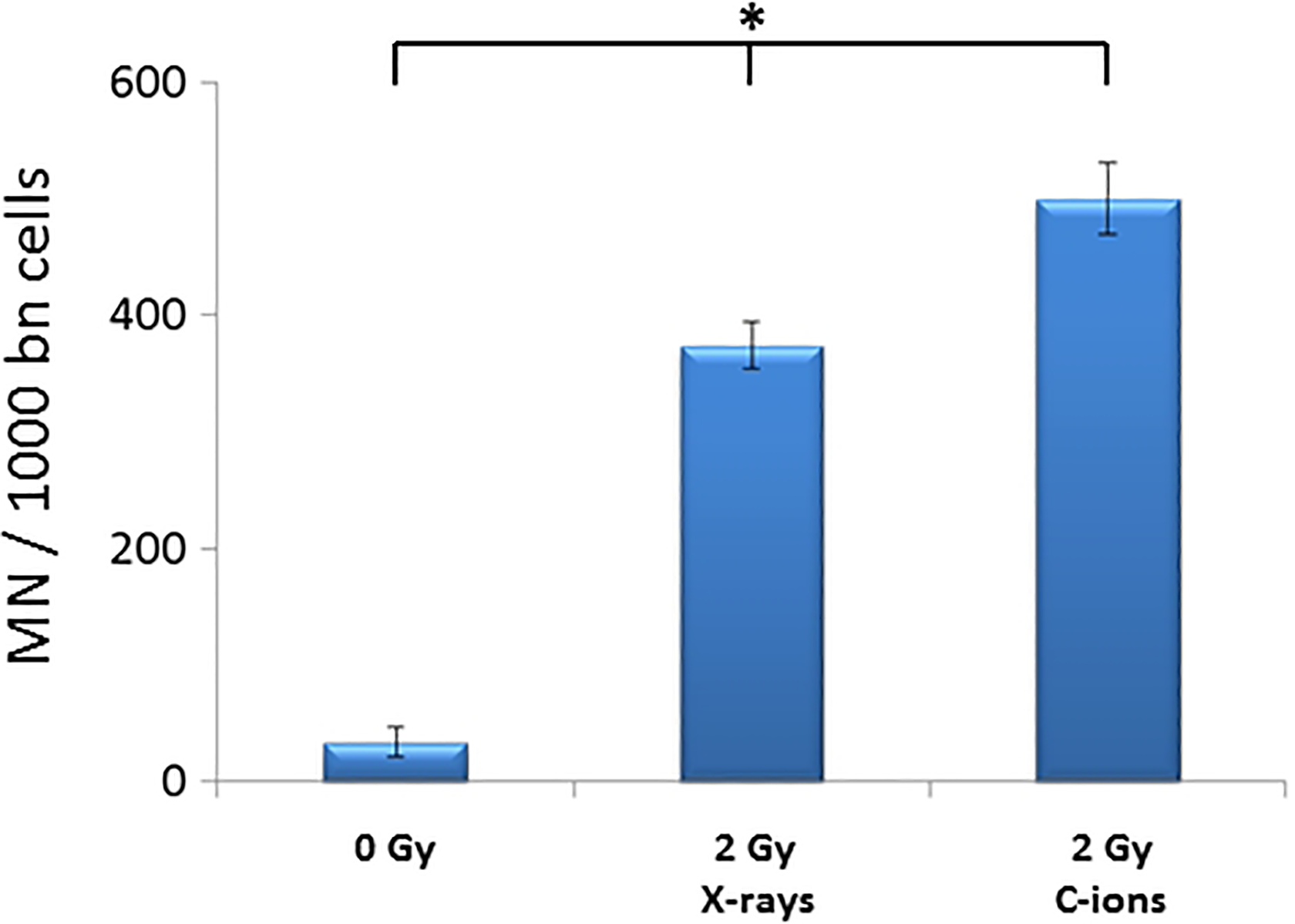
Micronucleus induction in sw1353 chondrosarcoma cells irradiated with X-rays and 50 keV/µm C-ions. Results from X-rays experiments are means of 2 independent experiments. Results from the C-ions experiment are from 1 irradiation experiment. The data presented are a mean (with standard errors) of at least 3 technical repetitions. One-way ANOVA on raw data was performed at the 0.05 level; the mean values were significantly different (*), P = 2.07 E-8. ANOVA indicates analysis of variance.
C-Ion Irradiation Delayed DNA Damage Repair
Following irradiation (X-rays and C-ions 28 keV/µm), SW1353 cells were analyzed by Western blotting to estimate the relative amount of specific proteins markers (γ-H2AX and p21) of irradiation responses (Figure 5). With both irradiation qualities, γ-H2AX level was maximum at 1 hours after irradiation and then decreased until 96 hours following irradiation. The maximum level of γ-H2AX was higher with X-rays, compared to C-ions, but a tendency to a plateau occurred with C-ions from 12 hours after irradiation, while a constant decrease was observed with X-rays (Figure 5A and B, top), in agreement with a study in human fibroblasts. 32
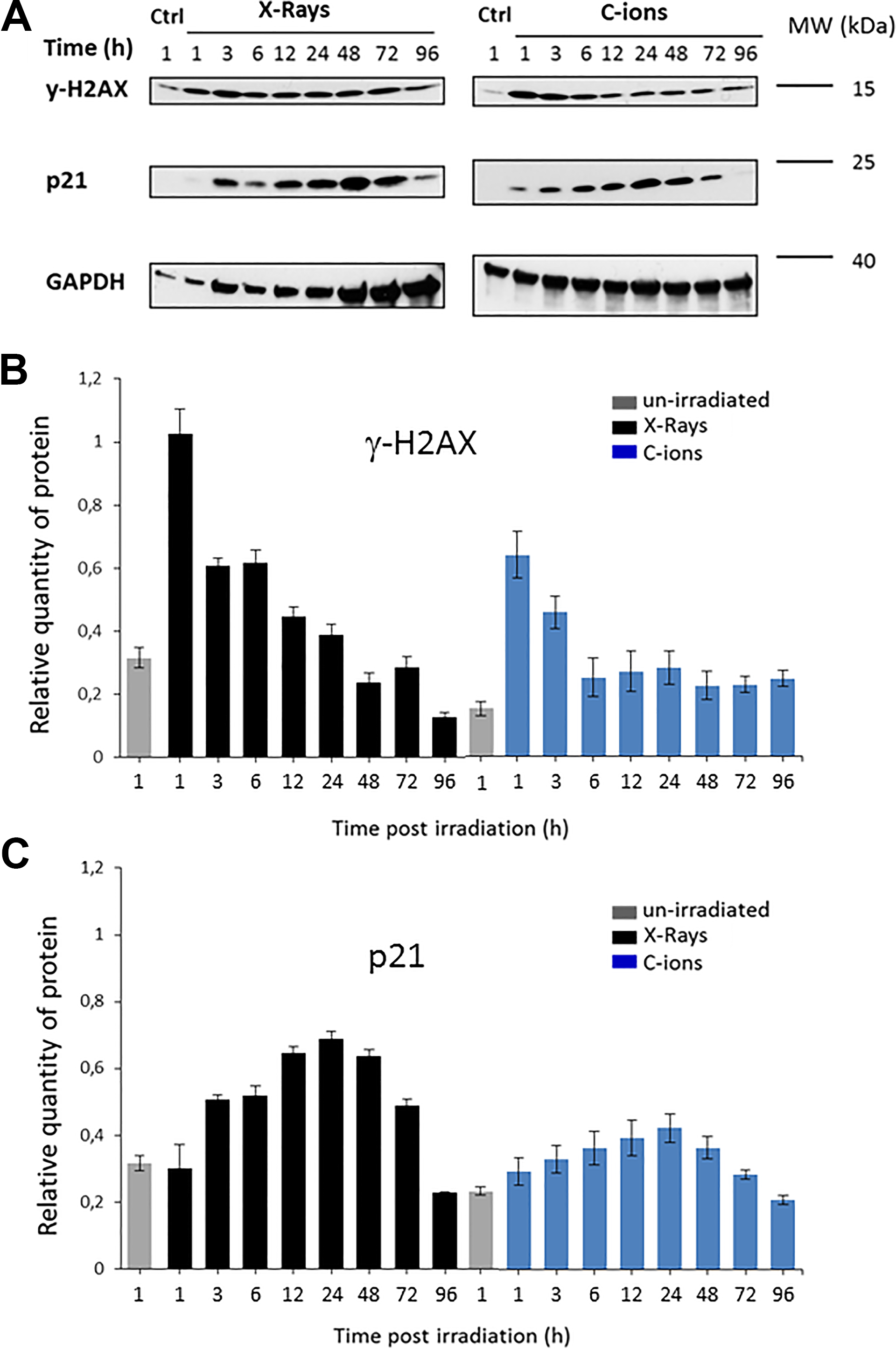
Western blotting analysis of Sw1353 cells at different time points following irradiation (X-rays and C-ions 28 keV/µm). SW1353 cells were analyzed by Western blotting to estimate the variation kinetics of specific proteins markers to irradiation responses. A control (0 Gy) at 1 hour and irradiated samples (2 Gy) were compared from 1 to 96 hours following treatment. Different antibodies were used, γ-H2AX, p21, and GAPDH were analyzed by Western blotting (A) and the corresponding signal was quantified with the Quantity One software (Bio-Rad, Hercules, California) and normalized against the signal of the GAPDH loading control (B). Gamma-H2AX and X-rays partially from Hamdi et al. 22 .
The level of the cell cycle inhibitor p21 increased from 1 hour after irradiation, with a maximum at 24 hours (Figure 5B, middle). Both irradiation types increased p21 expression in the DNA damage response, but with a lower level after C-ion irradiation.
Discussion
Chondrosarcomas are mainly considered as resistant to conventional radiotherapy (X-rays). But in case of a nonresectable localized disease and the ineffectiveness of chemotherapy, radiotherapy is the only available treatment. 7 Owing to the close proximity of risk organs (brain, cochlea, brain stem), a dose increase is difficult with X-rays in case of commonly observed localization of chondrosarcoma (skull base). Consequently, the efficiency of radiotherapy against such X-ray-resistant tumor needs to be improved, and the use of accelerated ions can be a solution. 33 Indeed, such nonconventional radiotherapy (with protons and/or C-ions) has proven its capacity to improve treatment against X-ray-resistant cancers. 34 The present study indicated that heavy ions may be a superior method at identical physical doses to induce chondrosarcoma cell killing in vitro compared to X-rays, based on using 4 chondrosarcoma cell lines with various radiosensitivities. Numerous studies have already shown that high LET irradiation, involving accelerated heavy ions, are more effective for cell killing than low LET irradiations. 17,21,35,36 High LET radiation qualities are considered to generate a large number of complex clusters of DNA damage which would be more difficult to repair, resulting in more severe biological damages than that induced by low LET radiation. 37,38
Moreover, the clonogenic survival data confirmed the trend previously shown in prostate cancer in an in vivo study, that there is much larger variation in radiosensitivity with photons than with high LET radiation. 39
In the present study, the RBE values for the 4 chondrosarcoma cell lines ranged from 2.04 to 7.30 relative to X-rays, depending on the LET of C-ions beams and the calculation of the factor, with D10 or D37 ratios. We observed that RBE was lower using the X-ray-sensitive cell line (L835). With the 3 X-ray-resistant chondrosarcoma cell lines, the RBE was about 2.2 using D10 ratio and an LET of 28 keV/µm LET, and about 5.2 at 73 keV/µm LET. No correlation could be done between cell response to radiation and p53 status, since only SW1353 cells appeared as p53 mutated (V203L), CH2879, OUMS27, and L835 were p53 wild type. 23,40
Taken together, all these results clearly show that the analysis of one parameter was not enough to understand the radiosensitivity of a cell line. The RBE of C-ions was indeed related to the increased cell killing effect of high LET irradiation, while it was related to the relative radiosensitivity of cell of the X-rays too (Table 1).
These clonogenic survival assays allowed us to classify the cell lines (from the less resistant to the most resistant) according to their radiosensitivity to: X-rays: L835 < CH2879-SW1353-OUMS27 (P < .05) C-ions at 28 keV/µm: L835 < CH2879 < OUMS27-SW1353 (P < .05) C-ions at 73 keV/µm: CH2879 < L835 < SW1353-OUMS27 (P <.05)
These RBE values of chondrosarcoma cells were in the range of the previously published studies on various tumors. 16,17,21
These values were of particular interest in relation to the use of RBE-corrected dose (Cobalt Gray equivalent) in particle therapy. But in order to follow the recommendation about harmonization of radiobiological studies, 41 we kept for this whole article physical dose in Gy.
The present study proposes for the first time a comparison between C-ions with different LET and X-rays on 4 chondrosarcoma cell lines. Since these types of cell lines were irradiated for the first time in those conditions, and in vitro, it is difficult to extrapolate these results to medical treatments. Nevertheless, the RBE obtained with high LET C-ions on X-ray-resistant cells were higher than previously reported RBE on osteosarcoma, 21 showing the promising potential of such irradiation on the control of chondrosarcomas.
In the present study, we confirmed the difficulty of cells to repair high LET irradiation damages, with different technical approaches on a single X-ray-resistant chondrosarcoma cell line. The SW1353 chondrosarcoma cell line was selected for further and deeper analysis of DNA repair mechanism following high LET irradiation. Indeed, C-ions irradiation induced a prolonged blockage of SW1353 cells in G2 phase and a concomitant high level of γ-H2AX protein, in agreement with previous studies in prostate and colon cancer cells. 42 Compared to X-rays, C-ions irradiation induced 30% of additional micronuclei. This result is coherent with another study showing an increase in the MN frequency as a function of LET after 2 Gy irradiation with the SW1353 cell line. They observed from 35% of binucleate cells with micronuclei with C-ions (LET = 13 keV/µm), to 80% of binucleate cells with micronuclei with iron ions (LET = 151 keV/µm). 43
In conclusion, the results of the present study provide a basis and a radiobiological confirmation into the possibility of application of heavy ions in medical treatment of chondrosarcoma. 7,33 With the use of several X-ray-resistant chondrosarcoma cell lines, this study clearly pointed out the significant effectiveness of high LET C-ions in killing chondrosarcoma cells, with physical doses comparable with X-rays. X-ray-resistant cells exhibited higher RBE values for C-ions. These findings support the capacity of accelerated ions in the treatment of X-ray-resistant tumor. Nevertheless, many radiobiological studies are still needed to understand all the mechanisms involved in the biological activity of heavy ions, including the role of the isocitrate dehydrogenase enzyme mutation status, 44,45 the possibility of neighborhood effect, that is, bystander effect, 46,47 and the accessibility to radiosensitizing chemicals, for example, Parp inhibitors. 33,48 All these areas of study are of great interest for radiobiologists, in order to improve the treatment of resistant and aggressive tumor and to preserve surrounding healthy tissues.
Supplemental Material
Supplementary_data_1 - High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines
Supplementary_data_1 for High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines by Francois Chevalier, Dounia Houria Hamdi, Charlotte Lepleux, Mihaela Temelie, Anaïs Nicol, Jean Baptiste Austry, Paul Lesueur, Guillaume Vares, Diana Savu, Tetsuo Nakajima and Yannick Saintigny in Technology in Cancer Research & Treatment
Supplemental Material
Supplementary_data_2 - High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines
Supplementary_data_2 for High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines by Francois Chevalier, Dounia Houria Hamdi, Charlotte Lepleux, Mihaela Temelie, Anaïs Nicol, Jean Baptiste Austry, Paul Lesueur, Guillaume Vares, Diana Savu, Tetsuo Nakajima and Yannick Saintigny in Technology in Cancer Research & Treatment
Supplemental Material
Supplementary_data_3 - High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines
Supplementary_data_3 for High LET Radiation Overcomes In Vitro Resistance to X-Rays of Chondrosarcoma Cell Lines by Francois Chevalier, Dounia Houria Hamdi, Charlotte Lepleux, Mihaela Temelie, Anaïs Nicol, Jean Baptiste Austry, Paul Lesueur, Guillaume Vares, Diana Savu, Tetsuo Nakajima and Yannick Saintigny in Technology in Cancer Research & Treatment
Footnotes
Acknowledgments
The authors thank Prof. Judith V.M.G. Bovée and Ms. Yvonne de Jong from the Department of Pathology (Leiden University Medical Centre, Leiden, the Netherlands) and Dr. Nicoleta Moisoi from the Leicester School of Pharmacy (De Montfort University, United Kingdom), for valuable discussions and helpful suggestions and comments.The cell line L835 was kindly given by Prof. Judith V.M.G. Bovée (Leiden, the Netherlands); the cell line CH2879 by Prof. Antonio Llombart-Bosch (University of Valencia, Spain) and the cell line OUMS27 by Prof. Masayoshi Namba (Okayama University, Japan).The authors thank Florent Durantel from CIRIL platform (Cimap Caen, France) for C-ions Dosimetry, Maryline Guillamin (University of Caen) for flow Cytometry support and the GANIL (Caen, France) and NIRS (HIMAC, Chiba, Japan) as C-ions irradiation facilities staff.
Author Contributions
FC and YS conceived the study and its design. FC, DHH, CL, MT, JBA and PL performed the experiments. AN performed the statistical analysis. FC prepared the figures and analyzed the data. FC, DS, GV and TN drafted the manuscript. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by “Agence Nationale de la Recherche”, in the frame of the “Investments for the Future” under the references France HADRON (ANR-11-INBS-0007) and Equipex Rec-Hadron (ANR-10-EQPX-1401) ; Région Normandie (IRHEMME project 2015-2018) ; Electricité de France funding from Life Sciences group of the four-way national agreement CEA-EDF-IRSN-Areva ; French Institute of Romania (MT); French Ministry of Foreign Affairs – Partenariat Hubert Curien Sakura (35973SB) ; CEA, Life Science Division funding for Radiation biology and Irtellis funding for PhD (CL).
Supplemental Material
Supplemental material for this article is available online.
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.