
Abstract
Background:
Immunotherapy, especially immune checkpoint inhibition, is one of the most sophisticated approaches in cancer therapy. Immune checkpoint inhibition has already been successfully applied for treatment of non-small cell lung cancer and various other entities. Unfortunately, 60% of the cases show signs of therapy resistance. Additionally, a proportion of cases shows initial insensitivity to immune checkpoint inhibition. We consider a novel escape mechanism in association with deficient proteasomal epitope processing to be one prominent reason for initial insensitivity and therapy resistance. Therefore, we aim to identify mutations in association with these so-called processing escapes, in a highly diverse collective of pulmonary neuroendocrine lung tumors.
Materials and Methods:
Seventy representative tumor specimens of pulmonary neuroendocrine lung tumors were analyzed retrospectively via immunohistochemical detection of CD4, CD8, CD68, and CD20 as well as programmed cell death protein 1 and programmed cell death 1 ligand 1 for tumor immune infiltration and composition. Afterward, samples were screened for alterations in 48 genes, including 221 known mutational hotspots by massive parallel sequencing using the Illumina TruSeq Amplicon-Cancer Panel. For prediction of proteasomal cleavage probabilities, an R implementation of the machine learning tool NetChop 3.1 was utilized.
Results:
Immune cell infiltration of different compositions could be found in the majority of tumors. Deficient epitope processing was revealed to be a common event in those with steady distribution across all different subtypes. Despite immune infiltration, no significant antitumor response could be detected.
Conclusion:
Since it is widely acknowledged that tumors need to avoid the immune system to ensure their survival, processing escapes should already be present during primary tumor development. In line, processing escapes can be found in all tumors, regardless of subtype and mutational burden. Furthermore, there is solid evidence that processing escapes have a negative impact on the antitumor activity of tumor infiltrating immune cells.
Introduction
Lung cancer is the leading cause of cancer-related deaths worldwide. 1 –3 One-quarter of these belong to the group of neuroendocrine tumors (neuroendocrine lung cancer, NELC) encompassing small-cell lung carcinoma (SCLC), large-cell neuroendocrine carcinoma (LCNEC), as well as the typical carcinoid (TC) and atypical carcinoid (AC) tumors. 4,5 Five-year survival rates vary greatly depending on the particular tumor entity. Patients with TC have excellent survival rates of 87% to 100% 6 –15 ; ATs are more aggressive (survival rate 61%-88%) and show a higher frequency of nodal metastasis. 8 –14,16 The LCNEC was assigned to the non-NSCLCs but shows similar biological behavior to SCLC with a 5-year survival rate of 15% to 57%. 4,8 The SCLC is associated with the poorest survival rates with <5%. 4,5,7,8,15,17,18
Established treatments for NELCs include surgical resection (carcinoids) as well as chemotherapy (SCLC). 4,6,19,20 Due to the rarity of LCNEC, an optimal treatment is still under investigation. 4
One of the most sophisticated cancer therapies is immunotherapy. 21 –23 Due to a different antigenicity of the tumor, a physiological immune response is triggered. 21,24 –26 One prominent part of the antitumor response depends on cytotoxic T lymphocytes (CTLs), which interact with tumor cells and driving them into apoptosis via Fas/FasL interaction or the release of granzymes. 21,24,25,27
One distinct therapeutic approach involves the usage of monoclonal antibodies, especially α-cytotoxic T-lymphocyte-associated protein 4 and α-programmed cell death protein 1/programmed cell death 1 ligand 1 (α-PD-1/PD-L1), for immune checkpoint inhibition. 26,28,29 These targets are well-known immune regulators responsible for downregulation of T-cell responses. 21 The application of specific inhibitors in combination with chemotherapeutic agents has shown promising results in clinical trials on patients with SCLC. 30,31
Besides others, tumor mutational burden (TMB), representing the genetical variance of each single tumor, has been proven as a promising predictive biomarker for immunotherapy. Tumors with a high number of somatic mutations show significantly improved response rates, making immune checkpoint inhibition an emerging therapeutic strategy in certain entities with high TMB. 32,33 It is generally accepted that high TMB levels may result in numerous tumor-specific antigens. Those tumor neoantigens are further processed by the proteasome, and, like most intracellular peptides, they are presented on human leukocyte antigen (HLA) class I molecules. Some CTLs are specialized to recognize these neoantigens, leading to a tumor-specific immune response. 34 Comprehensive research revealed their potential as a diagnostic tool or biomarker, but also as a target to improve tumor immunotherapy, for example, T-cell reactivity. 35,36 Nevertheless, the specificity and sensitivity of TMB regarding response to immune checkpoint inhibition does not meet the primary expectations, and the reasons are not conclusively clarified. 37 –39 One possible explanation comprises alteration in processing and presentation of tumor-specific epitopes, hiding cancer cells from hosts immune system.
In detail, the activation of tumor-specific T cells occurs via presentation of small peptide fragments (epitopes) originating from tumor antigens. 40,41 A complex intracellular pathway is involved in the processing of those antigenic peptides. Starting with polyubiquitination of the protein, marking them for proteasomal degradation, the small fragments are further trimmed to an optimal length of 8 to 11 amino acids. These get translocated into the endoplasmic reticulum via transporter associated with antigen processing and loaded on the HLA class I complex. Finally, the complex is presented on the cell membrane. 40,41
A failure of correct epitope processing might result in an immune escape. Investigations of viral infections, in particular with the HIV 1 and hepatitis C virus (HCV), revealed a subset of mutations altering proteasomal digestion of the viral proteins. 42,43 This leads to modified epitopes with different lengths, which are less effective in activating CTLs. 42
To address the question, whether deficient proteasomal processing plays a role in tumor development and immune evasion, we aimed to correlate genetic variances to deficient proteasomal processing. Therefore, we analyzed whether deficient processing of epitopes occurs in NELC; whether the presence of deficient processing is associated with different subentities or clinicopathological parameters; and whether those altered epitopes differentially trigger immune response
We chose a cohort comprising all 4 different subentities of NELC for investigation purposes, as they display a broad range of biological behaviors (low-grade vs high-grade). Nevertheless, they probably derive from the same pulmonary neuroendocrine stem cells. However, differences in general mutational burden and the behavior toward the immune response can be observed. Consequently, the variation and influence of deficiently processed epitopes between those different subtypes may expand the insight into processing escapes as an evolutionary conserved mechanism of tumor immune evasion.
Materials and Methods
Demographic Data and Study Design
This retrospective sequencing study is based on a collective of 70 formalin-fixed, paraffin-embedded pulmonary neuroendocrine tumors (17 TC, 17 AC, 19 LCNEC, and 17 SCLC). 44 The initial diagnosis was reevaluated by 2 experienced pathologists (TH and TM). Specimens were collected at the archives of the Institute of Pathology, University Hospital Essen (Germany), from 2005 till 2012. Tumor classification is based on the World Health Organization classification of tumors guidelines. 45 Tumor node and metastasis staging is based on the Union internationale contre le cancer classification of malignant tumors. 46 The mean age at the time of diagnosis was 58.6 years (median age: 59.0 years; 95% confidence interval, 50.8-66.9 months). Survival data were available for 34 patients (high-grade tumors only) with 22 reported deaths at the time of data collection. All specimens were resected prior to (systemic) chemotherapy. The study was approved by the ethical committee of the University Hospital Essen (ID: 13-5382-BO). The investigations conform with the principles outlined in the Declaration of Helsinki.
Immunohistochemical Staining of Immunophenotype
Formalin-fixed tissues were embedded in paraffin and processed into 4-μm-thick slides for histomorphological diagnosis (hematoxylin and eosin sections) and immunohistochemistry. The slides were incubated for 2 hours at 50°C. Xylene derivates were used for deparaffinization purposes. For pretreatment, CC1 (EDTA-)-buffer was applied. Immunohistochemistry was performed according to standard protocols using an automated stainer (Ventana Discovery XT, Munich, Germany). Table 1 lists applied antibodies with the respective dilutions and incubation times used for immunological staining.
Individual Treatment of Antibodies Used for Immunological Staining.a

Abbreviations: PD-1, programmed cell death protein 1; PD-L1, programmed cell death 1 ligand 1.
a All Antibodies Were Incubated at 37°C.
Nucleic Acid Preparation
Genomic DNA was isolated on a Maxwell 16 Research (Promega Corporation, Madison) as recommended in the manufacturer’s protocol. Nucleic acid quantification was performed using Qubit (Life Technologies, Carlsbad) and Nanodrop 1000 instrument (Thermo Fisher Scientific, Waltham).
Next-Generation Sequencing
As described previously, 44 samples were prepared using the TruSeq Amplicon—Cancer Panel (Illumina Inc, San Diego, California) followed by paired-end sequencing on the MiSeq personal sequencer (Illumina) according to the protocols provided by the manufacturer. The panel covered 48 tumor-relevant genes including 221 mutation hotspots. Library construction followed the TruSeq Custom Amplicon–Library Preparation guide.
FastQ-files were aligned against the Hg19 build. For analysis of the aligned reads including variant calling, the software Avadis next-generation sequencing (NGS; Strand Scientific Intelligence, California) was used. Reads with a quality score <30 were discarded. Filtering was performed by excluding variants with <25 effective variant reads or below 10% variation frequency. Synonymous variants were removed.
Statistical and Bioinformatical Analysis
All bioinformatical, statistical, and graphical analyses were performed using the R programming environment (v. 3.4.2).
Epitope search of affected genes
Based on the number of cancer-related genes and default epitope length (9 amino acids), databases were browsed for all available epitope information of affected genes. Relevant information includes the exact epitope sequence as well as the mutated position. Moreover, the output also comprised the most important HLA information, for example, the HLA type preferably recognizing the epitope and the binding affinity of each HLA molecule (half maximal inhibitory concentration [IC50] value).
Prediction of epitope processing and proteasomal cleavage
For prediction of proteasomal cleavage of each antigen, an R implementation of the machine learning tool NetChop 3.1 was utilized. The algorithm was trained with in vitro data for proteasomal digestion (NetChop 20S) as well as in vivo data of major histocompatibility complex (MHC)/HLA class I ligand structure (NetChop Cterm). 47 –49 According to previous research, 50 –52 up to 8 amino acids upstream and downstream of a positional cut might influence the cleavage probability due to their chemical composition. So, in addition to the complete epitope, 8 amino acids for each flanking region (N- and C-terminal) were added to the NetChop input. For any given amino acid position, NetChop calculates and outputs a cleavage probability. In order to compare the differences of a proteasomal cut between wild-type and mutated sequences, NetChop was executed 2 consecutive times with the wild-type and mutated sequences, respectively. The absolute difference in cleavage probability, between wild-type and mutated positions, was calculated. Any difference >50% between wild-type and mutated position was considered a differential proteasomal cleavage.
Statistical analysis in R
Before starting the analysis, the Shapiro-Wilks test was applied to test for normal distribution of the data. 53 Based on the results, either a parametric or a nonparametric test was applied. For dichotomous variables, either the Wilcoxon Mann-Whitney rank-sum test (nonparametric) or 2-sided Student t test (parametric) was applied. 54 For ordinal variables with more than 2 groups, either the Kruskal-Wallis test (nonparametric) or the analysis of variance (parametric) was used to detect group differences. Double dichotomous contingency tables were analyzed using Fisher exact test. To test dependency of ranked parameters with more than 2 groups, Pearson χ2 test was used. Correlations between metric variables were tested using the Spearman rank correlation test as well as the Pearson product moment correlation coefficient for linear modeling. Kaplan-Meier analysis was done for the assessment of associations between gene expression and progression-free survival (PFS) or overall survival (OS). Significant differences in PFS or OS between the groups were verified by Cox proportional hazards (COXPH) model using Wald test, likelihood ratio test, and Score (log rank) test. Due to the multiple statistical tests, all P values were adjusted using the false discovery rate. The level of statistical significance was defined as P ≤ .05 after adjustment.
Results
Tumor Infiltration and Antitumor Responses
Paraffin-embedded, formalin-fixed tissue of all affected patients was stained for PD-1/PD-L1 expression. Additionally, monoclonal antibodies targeting different immune markers, including CD3 (T cells), CD4 (T-helper cells), CD8 (CTLs), CD68 (monocytes), and granzyme B, were also applied (Figure 1). Statistical analysis was performed to identify correlations between the expression of immune factors and clinicopathological covariates. Of the 198 calculations, 3 immune markers (CD3, CD8, and CD68) had significant association (P < .05) with the presence of deficiently processed tumor epitopes in general. Strikingly, low amounts of shortened/prolonged epitopes were often associated with a low infiltration rate of CD3/CD8- and/or CD68-positive cells (Figure 2). On the other hand, an increase in altered processing also correlates with immune cell infiltration into the tumor by CD3/CD8- and CD68-positive cells. All samples were negative for granzyme B expression.

(A) Example of increased intratumoral CD3-positive T lymphocytes (×40), (B) example of increased intratumoral CD5-positive T lymphocytes (×100), (C) slight increased intratumoral CD20-positive B lymphocytes (×40), (D) increased intratumoral CD68-positive macrophage reaction (×200), (E) slightly increased intratumoral CD4-positive lymphocytes (×200), (F) increased intratumoral CD8-positive lymphocytes (×100), (G) increased PD-1-positive intratumoral lymphocytes as well as a slight tumor reaction for PD-1 (clone NAT105, ×100), (H) moderate PD-L1 (clone 22C3) expression in tumor cells (TPS 1%-49%; ×400). PD-1 indicates programmed cell death protein 1; PD-L1, programmed cell death 1 ligand 1.

Associations between mutations (overall) affecting processing and immune cell infiltration into the tumor. Contingency tables were used to calculate the abundance of associations. The amount of immune markers were either quantified via H-score method (.HS) or the staining intensity in relation to tumor mass (.TU). Cases showing no processing are shown in light blue, those harboring processing escapes are highlighted in red. Y-axis show overall sample count for each feature, X-axis differentiates between samples with (“high”) or without (“low”) infiltration of certain immune cells into the tumor. The P value was calculated via Fisher exact test.
Altered Proteasomal Cleavage is a Common Event in NELCs
Based on the NetChop analysis, 97 unique cleavage sites were identified across 27 cancer-associated proteins, including TP53, PIK3CA, KRAS, and APC (Supplementary table 1). Of note, these cleavage sites affect a total of 842 epitopes associated with different HLA types. Considering all existing cleavage sites, proteasomal cuts occurred more frequently in the flanking regions (535) than inside the epitope sequence (307). Generally, 2 different variables were used for analyzing purposes: (1) mutations that influence proteasomal processing (n = 110) and (2) epitope variants caused by proteasomal cleavage (n = 858).
Altered Proteasomal Cleavage is Associated With Clinicopathological Factors
The statistical analysis revealed about 30 significant correlations between tumor-associated proteins and several clinical covariates (P < .01). A list of these correlations, including the applied tests and P values, can be observed in Table 2. Deficient processing in general (mutations across all affected proteins) seems to have a significant impact on lymph node metastasis (Figure 3). Additionally, altered processing of TP53 also affects the survival of patients, while alterations in general (mutations/epitope variances across all affected proteins) as well as those in RB1 had influence on time to tumor progression. Besides the formation of lymph node metastasis, no association between the presence of deficiently processed epitopes and lymph vessel infiltration, blood vessel infiltration, or distant metastasis could be found (Supplementary table 2). Moreover, deficient processing correlates with the expression of NCAM1 (CD56; P = .027). Deficient processing overall does not seem to impact tumor staging or grading. However, deficient processing in single proteins such as ERBB2 or TP53 has an influence on either tumor staging (P = .046) or grading (P = .048). The analysis was performed with both variables, that is, mutations with an influence on proteasomal processing and epitope variances.
Statistical Correlations of Clinical Covariates With Protein Variations.a

a These mutations had a definite influence on proteasomal processing. Only significant associations are shown (P value <.01). The P values were adjusted with the help of false discovery rate (FDR). Different tests have been applied, depending on the data structure (see “Statistical Analysis” and “Methods” sections). Most abundant clinical covariates were lymph node invasion, time to tumor progression after initial therapy and survival status (binary variable, either dead or alive).
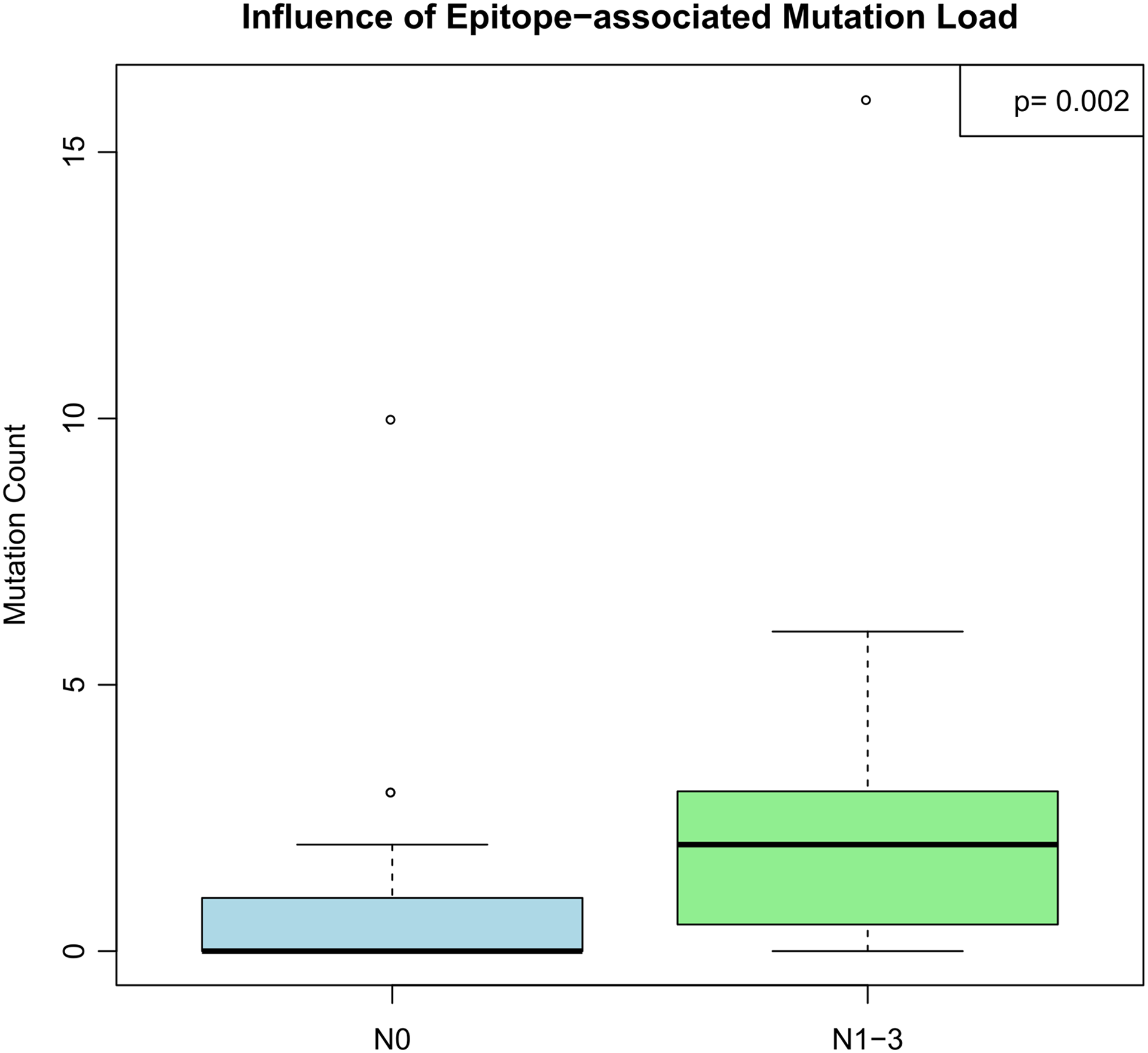
The effect of deficient processing in general on lymph node metastasis. The mutation count in dependence of the current lymph node status is displayed. The P value was calculated via Wilcoxon-Mann-Whitney rank-sum test.
Different Proteasomal Processing Affects Patient Survival
We further investigated how deficient proteasomal processing overall affects PFS. This variable was considered significant according to likelihood ratio tests and COXPH regression models. Two groups of patients diagnosed with high-grade NELCs (SCLC and LCNEC) were compared. One group harbors mutations that affect proteasomal processing, while the other group is devoid of such mutations. The PFS clearly differs between both groups, indicating a possible correlation between deficient processing and decreased PFS (Figure 4).

Effects of altered processing in general on progression-free survival (PFS). The decrease of PFS was plotted against survival in months. The P value was calculated via likelihood-ratio test.
Proteasomal Processing Does Not Significantly Differ Between Tumor Types
Additionally, it was investigated whether impaired processing varies between the different types of NELC. Aside from single counts in TC, AC, LCNEC, and SCLC, the sum of all mutations across all tumor types was calculated and visualized as overall count shown in Figure 5. Around 35% of the registered mutations had a significant influence on proteasomal processing. Around 35% of all mutations in AC, LCNEC, and SCLC were associated with deficient processing (Figure 5A). Only TC samples were shown to have fewer mutations correlating with deficient processing (22%). In comparison, the relative number of general mutations per tumor type was also displayed (Figure 5B). Low-grade NELCs (TC and AC) had fewer mutations than high-grade variants (LCNEC and SCLC). The highest number of mutations could be observed in SCLC. The results, shown in Figure 5B, served for comparison purposes only and were already described. 44

Mutation analysis in dependence of tumor type. The analysis was performed with data for deficient processing across all affected proteins (A) and general mutation count across all proteins (B). A, For every protein, the relative number of mutations causing deficient processing were plotted and splitted into 5 categories. One for each tumor type: TC (green), AC (yellow), LCNEC (red), and SCLC (dark red). In addition, the number of mutations was averaged across all tumor types (overall, blue). B, Same procedure as in (A), but with a different data set including general mutation data. TC indicates typical carcinoid; AC, atypical carcinoid; LCNEC, large-cell neuroendocrine carcinoma; SCLC, small-cell lung carcinoma.
Furthermore, we attempted to visualize the general occurrence of deficient processing across all tumor types via general unsupervised clustering (Figure 6). This clustering method was used to display the differences in affected proteins, per tumor type. Three clusters could be observed, with each tumor type represented in all clusters. In at least one-third of all observed cases (middle cluster, Figure 6), mutations affecting proteasomal cleavage were almost absent.

Associations between tumor type and mutations or epitope variances. By scaling each event (mutations/epitope variances), a normalized z score was calculated. In the heatmap, the z score is visualized via color code. The code displays the number of events that affect proteasomal processing for each protein per tumor type: more events than usual (yellow), average amount (black), and fewer than average (blue). Based on the distribution of events, the map can be divided into 3 clusters (white lines).
Tumor Progression and Immune Responses
It is generally accepted that the recognition of antigens is dependent on the structure of the MHC/HLA class I molecule or the HLA type. 55 Radar chart visualization was used to display, which HLA-types bind and present alternatively processed epitopes with higher binding affinity and thereby more frequently (Figure 7). HLA A*02 and HLA B*15 are the most important HLA types for recognition of alternatively processed epitopes.

HLA binding of mutated epitopes. The amplitude shows the HLA type with the highest affinity for mutated epitopes. The HLA types with the highest affinity are marked in red. HLA indicates human leukocyte antigen.
Discussion
Immune escapes involving deficient proteasomal processing were described first in viral infections. 42,43 In a recent review article, we introduced processing escapes as a possible explanation for different immunogenicity of cancers. 56
Generally, we consider deficient processing to a part of 2 possible mechanisms to subvert the immune response against the tumor. First, tumors develop mutations that cause deficient processing during its primary development to overcome immune surveillance. Furthermore, if immunotherapy is applied, natural evolution could drive the tumor population to favor tumors with deficient processing within the selection process. However, for homogeneity purposes, the patients in our collective were all treated adjuvant.
Our hypothesis of tumoral immune evasion is supported by the fact of different immunogenicity of tumors according to their number of processing escapes. Tumor tissue was stained for PD-1 and PD-L1 and other immune markers such as CD8 and granzyme B. High amounts of mutations associated with deficient processing correlate with a high rate of infiltrating immune cells (Figure 2). The expression for granzyme B was negative in all samples, although immune cells migrate to the tumor site. Granzyme B is a proapoptotic protein expressed by CTL and natural killer cells. It serves as one of the main mechanisms to destroy infected or neoplastic cells. 25,26 The association between a high amount of deficient processing and the infiltration of monocytic immune cells (CD68 positive) could also lead to the assumption that the tumor attracts immune cells for its own purposes, while simultaneously evading the antitumor response. Several studies proved that an inflammatory process, especially caused by cells of the innate immune response, at the tumor side will have severe tumor-promoting effects. 57 –59 This might indeed explain why a high number of mutations are associated with higher immune infiltration.
Methodical inaccuracies aside, patients not affected by mutations that lead to deficient proteasomal processing (45%), are still evading the immune response. However, in these specific cases, processing escapes are not responsible for causing the immune escape. Since immune evasion was added into the hallmarks of cancer, 59 it is generally accepted that potential neoplasms need to overcome permanent surveillance from the immune system in order to develop into full-blown malignancies.
As 54% of the patients carry mutations that correlate with deficient proteasomal processing, processing escapes might indeed be one of the most prominent mechanisms to initiate an immune escape. However, in 45% of the samples, no mutations leading to deficient proteasomal processing could be found, indicating that other mechanisms are also responsible for an immune escape. Other evasion mechanisms, not involving deficient processing, are dysregulation of HLA class I molecules, 60,61 which lowers the chance of being detected significantly or the development of an immune suppressive tumor microenvironment. 60,62,63 This also includes the recruitment of FOXP3-positive regulatory T cells. Especially, HLA downregulation was frequently observed (20%-60%) in melanoma, breast, renal, prostate, bladder, and lung cancer. 55,60 Conclusively, immune cells, including T cells and monocytes, are attracted to the tumor side. However, a significant antitumor response could not be detected. From this observation, it can be assumed that the tumor is actively evading the immune response via processing escapes and other mechanisms, while using local inflammation for progression.
Deficient processing in general was significantly correlated with increase in lymph node metastasis (Spearman rank correlation test, P < .003) as well as tumor progression (Wilcox rank-sum test, P < .001). Furthermore, these mutations had a negative impact on PFS of patients (Figure 4). Therefore, a correlation between progressive tumor development and the occurrence of processing escapes is presumed. This is further supported by the mutations in RB1 and TP53, which both influence time to tumor progression and PFS, respectively.
The NELCs are a diverse group of tumors, encompassing low-grade (TC and AC) as well as high-grade variants (LCNEC and SCLC). Due to this, we investigated the distribution of processing-altering mutations in general and the mutational burden (per tumor entity). Mutations leading to deficient processing (in general) were nearly evenly distributed throughout all tumor types, with exception of TCs (Figure 5A). This is a strong contrast to the distribution of the mutational burden per sample (Figure 5B). Additionally, the distribution of general processing-altering mutations is better visualized by general unsupervised clustering (heatmap; Figure 6). Because all NELC subtypes can be found in each heatmap cluster, even in a cluster with almost no associated mutations, deficient processing and subsequently processing escapes could be a general phenomenon in NELCs, defining 2 different groups with different tumor immunogenicity. That means, mutations affecting proteasomal processing and subsequently also processing escapes generally occur in NELCs, regardless of tumor grading and mutational burden. It is possible that the occurrence of deficient processing rises with the tumor grading due to the fact that some proteins like TP53 and RB1 carry more mutations in high-grade NELCs. However, taken all mutations affecting proteasomal processing together, it was shown that the difference between low grade and high grade is not that significant at all (Figure 5A).
Our results highlighted that more cleavage sites exist (537 vs 307) in the flanking regions of epitopes. Depending on the location of cleavage sites, the structure of alternate epitopes might differ. Cleavage sites in the flanking region might result in a C-terminal or N-terminal extended epitope, while cleavage sites within the epitope lead to shortened epitopes. Both claims are supported by the research of deficient processing in viruses. 42,43
Most genetic variants responsible for tumor immunogenicity occur from passenger and not driver mutations. Nevertheless, even when drivers are overrepresented within such a targeted, amplicon-based sequencing panel as we used in the present study, the majority of mutations detected and analyzed can be categorized as passenger mutations. As shown in a previous study, 64 classic drivers represent only a minor part of all found variants. Our data indicate that processing escapes are either present, originating from evolutionary selection pressure by the hosts immune system, or completely absent, commending it to alternative mechanisms of immune evasion. Taking all these aspects, together with the different immune cell infiltration and activity, into account, we are capable to generate the required information out of small, routinely in diagnostics used NGS panels.
In regard to how processing escapes relate to TMB or tumor neoantigens, we hypothesize that processing escapes are associated with a subset of differentially processed tumor-associated antigens or neoantigens. Some CTLs are specialized to recognize tumor neoantigens presented on HLA class I molecules, leading to a tumor-specific immune response (Figure 8A). 34 In contrast, a subset of tumor neoantigens result in alternatively processed epitopes. Upon presentation, those epitopes exhibit a weakened or reduced ability to trigger T-cell–mediated immune response compared to their wild-type homologous. A model describing the underlying mechanism of processing escapes was introduced previously 56 and is schematically illustrated in Figure 8. Chemical properties of proteins may change due to nonsynonymous genetic variants, resulting in different proteasomal cleavage properties of the antigens. In general, the proteasome contains multiple subunits with different cleavage preferences for acidic, basic, or hydrophobic amino acids. 64 Moreover, the immune proteasome induced by interferon γ secretion during an active immune response further specializes toward the cleavage of hydrophobic side chains. This indicates a preference of HLA class I molecules for hydrophobic peptides. 64 –66 This may lead to altered cleavage of mutated peptides, resulting in epitopes varying in length. 42,43 Despite this, deficient or incorrectly processed epitopes can show affinity to HLA class I molecules and thereby get presented on the cell surface. However, for most of these shortened or prolonged epitopes, the efficiency to activate T-cell receptor (TCR) signaling is reduced. Models for T-cell activation describe first a convergence between the target cell and the T cell, which is stabilized by different adhesion molecules such as CD2, CD48, intercellular adhesion molecule 1 (ICAM-1), and lymphocyte function-associated antigen (LFA-1). This allows the interaction between the HLA complex and the TCR, resulting in a conformational change of the TCR, which triggers intracellular signaling (Figure 8B). A weak or nonspecific interaction, however, leads to quick dissociation of the complex and a nonactivation of the T cell 67,68 (Figure 8D). Conclusively, the application of specific monoclonal antibodies would not impact the immune response against the tumor.

Mechanism behind processing escapes. The general mechanism for an HLA class I-driven immune response is shown (A, B). Mutations in tumor-associated antigens (neoantigens) are leading to alternatively processed epitopes. The peptide fragments are still presented via HLA class I molecule (C). The fragment is still recognized by the TCR. However, due to a weak activation signals, it results in nonactivation of the T cell. At the same time, antibody therapy of PD-1 or PD-L1 is rendered ineffective due to spatial separation between the cells (D). HLA indicates human leukocyte antigen; TCR, T-cell receptor; PD-1, programmed cell death protein 1; PD-L1, programmed cell death 1 ligand 1.
To find a mechanical explanation for processing escapes, we compared the binding affinity of natural and mutated epitopes and looked especially for changes of the IC50 values. A higher affinity of mutated epitopes increases the probability of a weak immune response. 34% of all mutations affecting proteasomal processing result in fragments with a higher affinity for the MHC-I/HLA-I molecule compared to the wild type. However, this applies only for mutations inside the epitopes. Since this explanation will not suffice, it will be necessary to use experimental in vitro approaches using mutated and nonmutated cleavage fragments in cell activity assays. 48,69,70 As a result, our understanding of the fundamental mechanics behind processing escapes and immune checkpoint inhibition will be significantly expanded. Nevertheless, we have already shown that some (34%) alternate epitopes are bound with a higher affinity than wild-type fragments. Therefore, mutations related to this mechanism might indicate the occurrence of actual processing escapes.
Conclusion
Mutations affecting proteasomal processing and subsequently also processing escapes can be found in all subtypes of pulmonary neuroendocrine tumors. Most important, they occur regardless of subtype and mutational burden. Furthermore, it is possible that processing escapes have a negative impact on the antitumor activity of tumor-infiltrating immune cells.
Outlook
Our long-term goal is to improve the workflow introduced in this article. We hope to provide a prediction tool that identifies patients affected by processing escape. We are convinced that with the help of such a prediction tool, the clinical management of patients will be improved significantly.
Supplemental Material
supplemental_table_1 - Processing Escape Mechanisms Through Altered Proteasomal Cleavage of Epitopes Affect Immune Response in Pulmonary Neuroendocrine Tumors
supplemental_table_1 for Processing Escape Mechanisms Through Altered Proteasomal Cleavage of Epitopes Affect Immune Response in Pulmonary Neuroendocrine Tumors by Michael Wessolly, Robert F. H. Walter, Claudia Vollbrecht, Robert Werner, Sabrina Borchert, Jan Schmeller, Elena Mairinger, Thomas Herold, Anna Streubel, Daniel C. Christoph, Wilfried E. E. Eberhardt, Jens Kollmeier, Thomas Mairinger, Kurt W. Schmid, Jeremias Wohlschlaeger, Thomas Hager, and Fabian D. Mairinger in Technology in Cancer Research & Treatment
Supplemental Material
supplemental_table_2 - Processing Escape Mechanisms Through Altered Proteasomal Cleavage of Epitopes Affect Immune Response in Pulmonary Neuroendocrine Tumors
supplemental_table_2 for Processing Escape Mechanisms Through Altered Proteasomal Cleavage of Epitopes Affect Immune Response in Pulmonary Neuroendocrine Tumors by Michael Wessolly, Robert F. H. Walter, Claudia Vollbrecht, Robert Werner, Sabrina Borchert, Jan Schmeller, Elena Mairinger, Thomas Herold, Anna Streubel, Daniel C. Christoph, Wilfried E. E. Eberhardt, Jens Kollmeier, Thomas Mairinger, Kurt W. Schmid, Jeremias Wohlschlaeger, Thomas Hager, and Fabian D. Mairinger in Technology in Cancer Research & Treatment
Footnotes
Authors’ Note
Thomas Hager, MD, and Fabian D. Mairinger, PhD, equally contributed to this work. All authors have read and approved the manuscript. All authors disclose any affiliations that are considered to be relevant and important with any organization that to our knowledge has any direct interest in the subject matter discussed. Patients were informed during their registration at the Hospital and signed a formal consent. A separate patient consent for research purposes has not been obtained exist; however, our study was approved by the ethical committee of the University Hospital Essen (ID: 13-5382-BO). The investigations conform with the principles outlined in the Declaration of Helsinki.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: No relevant funding was obtained. All resources were provided by the Department of Pathology, University Hospital Essen.
Supplemental Material
Supplemental material for this article is available online.
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.