
Abstract
Objective:
Interaction of advanced glycation end products (AGEs) with the receptor RAGE plays a role in diabetic nephropathy. However, effects of RAGE-aptamer on tubular damage remain unknown. We examined whether RAGE-aptamer inhibited tubular damage in KKAy/Ta mice, obese type 2 diabetic mice with insulin resistance.
Materials and Methods:
Male 8-week-old KKAy/Ta mice received continuous intraperitoneal infusion of either control-aptamer or RAGE-aptamer for 8 weeks. Blood biochemistry and blood pressure, and urinary N-acetyl-β-D-glucosaminidase (NAG) activity and albumin excretion levels were monitored. Kidney and adipose tissue samples were obtained for immunohistochemical analyses.
Results:
Although RAGE-aptamer did not affect blood glucose, blood pressure, body weight, or serum creatinine values, it significantly inhibited the increase in urinary NAG activity and HOMA-IR in diabetic mice at 12 and 16 and at 16 weeks old, respectively. Furthermore, compared with control-aptamer-treated mice, renal carboxymethyllysine, RAGE, and NADPH oxidase-driven superoxide generation were significantly decreased in RAGE-aptamer-treated mice at 12 weeks old with subsequent amelioration of histological alterations in glomerular and interstitial area, while adipose tissue adiponectin expression was increased.
Conclusion:
Our present results suggest that RAGE-aptamer could inhibit tubular injury in obese type 2 diabetic mice partly by suppressing the AGE-RAGE-oxidative stress axis and improving insulin resistance.
Introduction
Several randomized clinical trials revealed that inhibitors of sodium-glucose cotransporter 2 (SGLT2) reduced the risk of renal events in type 2 diabetic patients, regardless of the presence or absence of chronic renal disease.1–6 We have previously reported that uptake of glucose into proximal tubular cells via SGLT2 increases accumulation of advanced glycation end products (AGEs) and expression of their receptor RAGE in the kidneys of diabetic rats, thereby being involved in tubulointerstitial damage, a key mover in diabetic nephropathy.7–9 Furthermore, we recently found that albuminuria-lowering effects of SGLT inhibitors could be partly ascribed to its protective action against tubular damage in type 2 diabetic patients. 10 These observations suggest that activation of AGE-RAGE axis via SGLT2 in the tubulointerstitial areas could contribute to the progression of diabetic nephropathy, and therefore inhibition of the interaction of AGEs with RAGE may be a therapeutic target for diabetic nephropathy.
Anti-RAGE neutralizing antibodies have been shown to reduce mesangial area expansion and increase in urinary albumin excretion in both type 1 and type 2 diabetic animals.11,12 However, to date, RAGE antibodies have not been approved as a therapeutic tool for diabetic nephropathy due to safety concern and/or limited clinical utility. We have recently shown that DNA aptamer raised against RAGE (RAGE-Apt) significantly inhibits the binding of AGEs to RAGE and subsequently attenuates the AGE-induced reactive oxygen species generation and inflammatory reactions, including RAGE expression in human cultured mesangial cells. 13 Moreover, RAGE-Apt significantly blocked the development and progression of diabetic nephropathy in type 1 diabetic rats although it did not affect glycemic parameters or blood pressure. 13 However, the effects of RAGE-Apt on tubular damage in KKAy/Ta mice, obese type 2 diabetic mice with insulin resistance remain unknown. In this study, we examined whether RAGE-Apt inhibited tubular damage in KKAy/Ta mice.
Materials and methods
Preparation and selection of RAGE-Apt
Preparation and selection of RAGE-Apt and control-aptamer (Ctrl-Apt) were performed using SELEX as described previously. 13 Sequences of phosphorothioate-modified aptamers are follows: RAGE-Apt, 5′-ccTgATATggTgTcAccgccgccTTAgTATTggTgTcTAc-3′; Ctrl-Apt, 5′-ttcggCctgggGgcggcCagttcGggtccAgtcgcGggag-3′; phosphorothioate nucleotides are indicated as capital letters.
Animal experiments
Male 8-week-old KKAy/Ta mice (CLEA Japan, Inc., Tokyo, Japan) were used in the present experiments. RAGE-Apt (2.0 pmol/day/g body weight) or Ctrl-Apt was intraperitoneally administered to mice by an ALZET osmotic pump (Durect Corp., Cupertino, CA, USA). At baseline and after 4- or 8-week treatment, blood pressure was measured by a tail-cuff sphygmomanometer BP-98A (Softron Co., Ltd., Tokyo, Japan), and mice were housed in metabolic cages for 8 h to collect urine, sacrificed and then blood, kidney, and adipose tissue samples were obtained. Total cholesterol, high-density lipoprotein-cholesterol (HDL-cholesterol), triglycerides, creatinine, aspartate aminotransferase (AST), alanine aminotransferase (ALT), and blood urea nitrogen levels were measured using a chemistry analyzer FUJI DRI-CHEM 4000V (FUJIFILM Corp., Tokyo, Japan). Fasting blood glucose was measured by a STAT STRIP Xpress (Nova biomedical Corp., Waltham, MA, USA). Plasma insulin levels were measured by a LBIS Mouse Insulin enzyme-linked immunosorbent assay kit (U-type; FUJIFILM Corp.). Urinary creatinine was measured by a Serotec CRE-L (Serotec Co., Ltd, Sapporo, Japan). Urinary N-acetyl-β-D-glucosaminidase (NAG) activity was measured by a colorimetric assay kit (BioVision Inc, Milpitas, CA, USA). Serum cystatin C and adiponectin levels were measured with mouse cystatin C (Abcam, Cambridge, UK) and mouse adiponectin enzyme-linked immunosorbent assay kits (R&D Systems Inc, Minneapolis, MN, USA), respectively. Insulin resistance was estimated using the homeostasis model assessment of insulin resistance (HOMA-IR) from fasting blood glucose and insulin concentrations using the following formula: HOMA-IR = (fasting insulin [µU/mL] × fasting blood glucose [mg/dL])/405. All the animal experimental procedures were conducted in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals and were approved by the ethical committee of Kurume University School of Medicine.
Immunohistochemical analysis
Kidneys and visceral adipose tissues were obtained from KKAy/Ta mice, fixed with 4% paraformaldehyde, embedded in paraffin, cut into 4-µm intervals, and mounted on glass slides. The kidney sections were incubated overnight at 4°C with peroxidase-conjugated monoclonal antibody raised against carboxymethyllysine (CML; TransGenic Inc., Fukuoka, Japan) or anti-RAGE antibody (Santa Cruz Biotechnology Inc., Dallas, TX, USA) followed by Histofine Simple Stain MAX-PO(R) (Nichirei Bioscience Inc., Tokyo, Japan) as described previously. 13 The adipose tissue sections were incubated overnight at 4°C with antibodies raised against adiponectin (Santa Cruz Biotechnology Inc.), and then incubated with goat anti-rabbit IgG (H+L) Alexa Fluor 488 (Invitrogen Corp., Carlsbad, CA, USA). Immunoreactive areas were measured from ten randomly selected fields of kidney and adipose tissue sections per animal using the cellSens version 1.14 software (Olympus Corp., Tokyo, Japan).
Measurement of NADPH oxidase-driven superoxide generation
Renal NADPH oxidase-driven superoxide generation was measured by a luminescence assay in 50 mmol/L phosphate buffer (pH 7.0) containing 1 mmol/L ethylene glycol tetraacetic acid, 150 mmol/L sucrose, 5 µmol/L lucigenin as the electron acceptor, and 100 µmol/L NADPH as the substrate as described previously. 14
Histopathological examinations
The kidneys were fixed in 4% paraformaldehyde and embedded in paraffin, sectioned at 4-µm intervals and mounted on glass slides. The sections were stained with hematoxylin and eosin for light microscopic analysis. Glomerular area delimited by the internal edge of the Bowman’s capsule was measured using image J software as described previously. 15 Two-micrometer paraffin sections were stained with periodic acid-Schiff’s reagent. The cortical interstitial area and intra-tubular area were measured from ten randomly selected fields of tissue sample per each animal using the cellSens version 1.14 software (Olympus Corp., Tokyo, Japan).
Statistical analysis
All values were presented as mean ± standard deviation. Statistical comparisons were performed using ANOVA followed by Tukey-Kramer test (except for creatinine and cystatin C in Table 1, Figures 1(a), (c)–(e) and 2), Tukey HSD test (cystatin C at 12 weeks old), or Steel-Dwass test (creatinine, cystatin C at 16 weeks old, and Figure 1(b)); p < 0.05 was considered significant. All statistical analyses were performed with the use of JMP Pro version 14.0 (SAS Institute Inc., Cary, NC, USA).
Characteristics of animals.
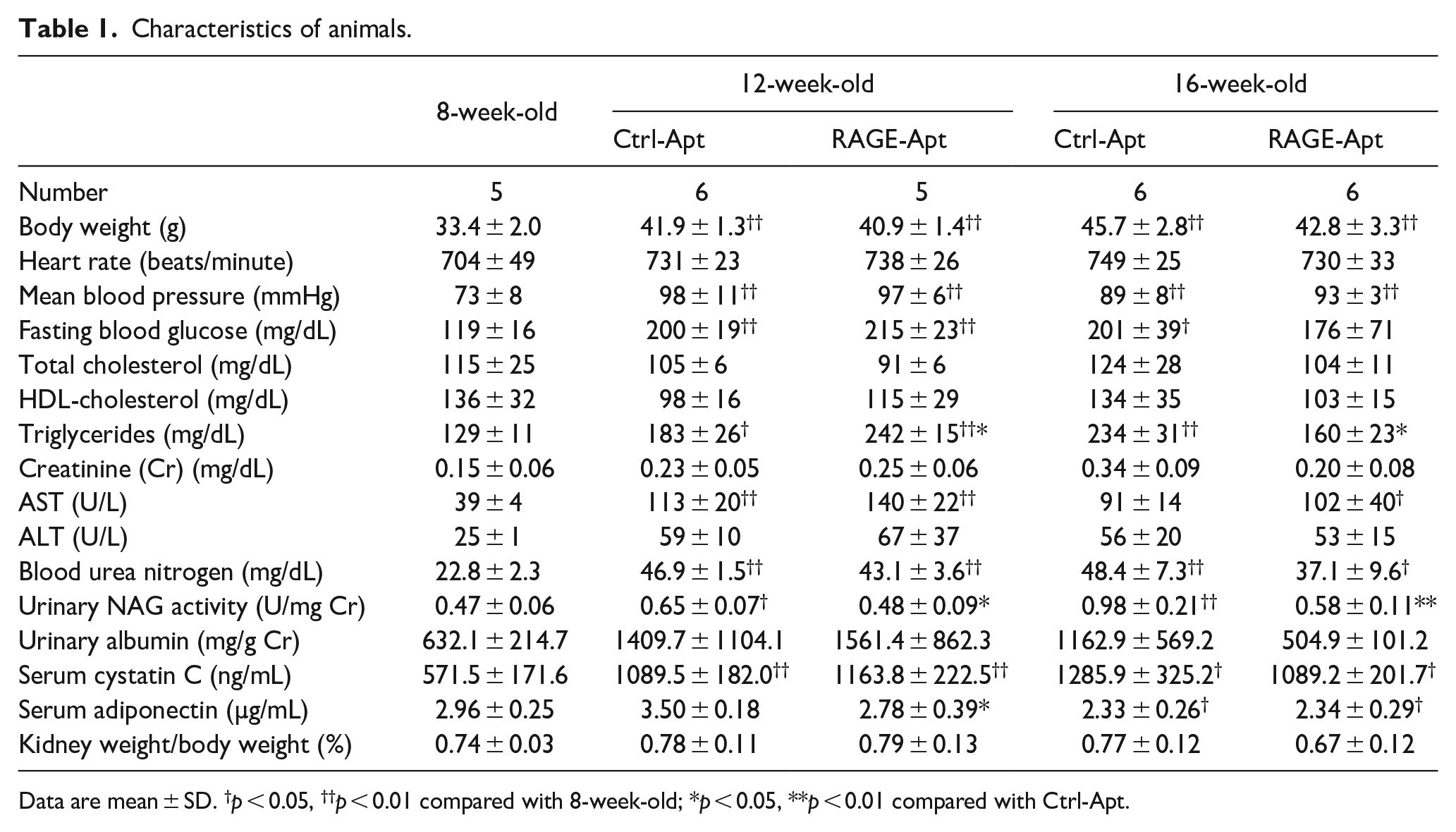
Data are mean ± SD. †p < 0.05, ††p < 0.01 compared with 8-week-old; *p < 0.05, **p < 0.01 compared with Ctrl-Apt.
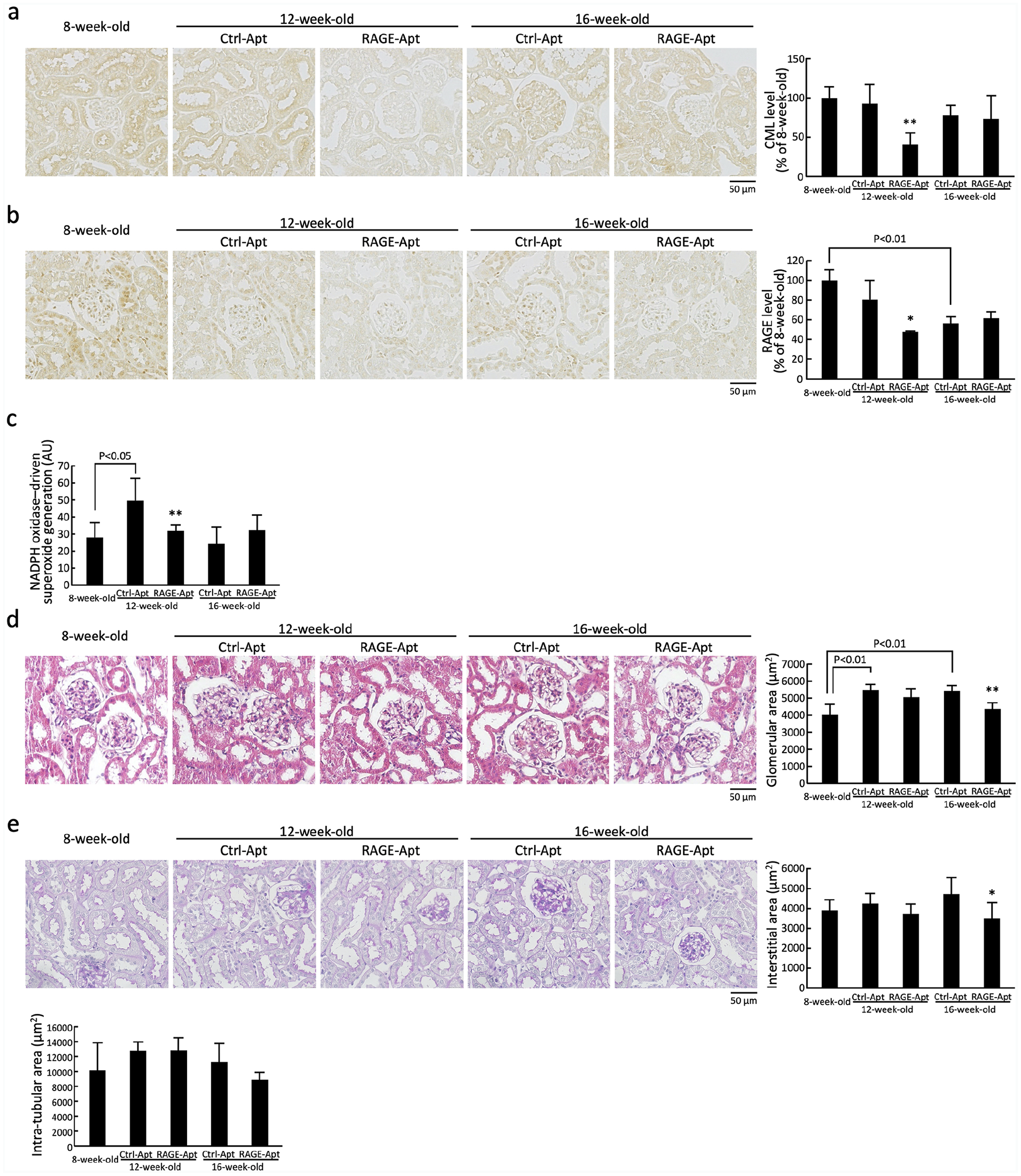
Renal CML (a), RAGE (b), NADPH oxidase-driven superoxide generation (c), glomerular area (d), and interstitial and intra-tubular area (e) in 8-week-old control mice and KKAy/Ta diabetic mice at 12 and 16 weeks old. (a and b) each left panel shows representative immunostainings of CML and RAGE in the kidneys. Each right panel shows the quantitative data, (c) NADPH oxidase-driven superoxide generation in the kidneys, and (d and e) each left panel shows representative stainings with hematoxylin and eosin (d), and with periodic acid-Schiff’s reagent (e).
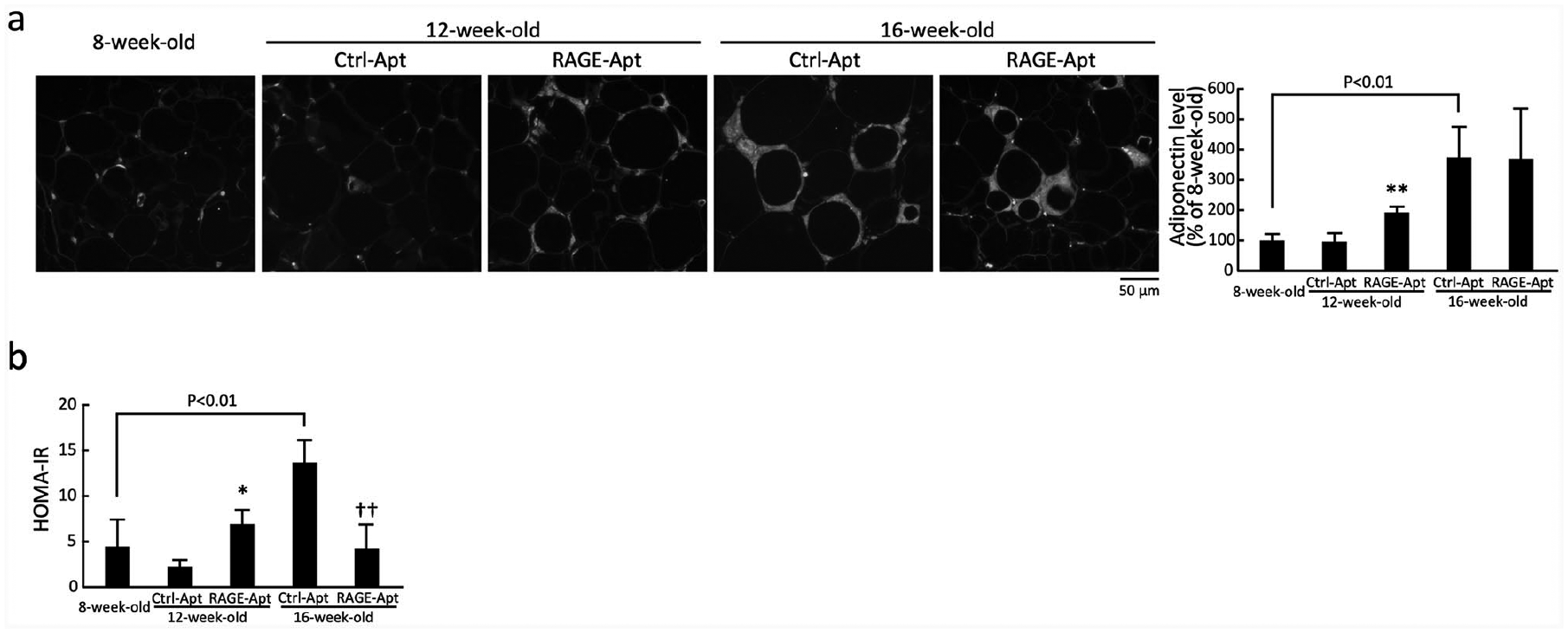
Adipose tissue adiponectin expression and HOMA-IR in 8-week-old control mice and KKAy/Ta diabetic mice at 12 and 16 weeks old: (a) left panel shows representative immunostainings of adiponectin in the adipose tissues of 8-week-old control mice and KKAy/Ta diabetic mice at 12 and 16 weeks old. Right panel shows the quantitative data and (b) HOMA-IR in 8-week-old control mice and KKAy/Ta diabetic mice at 12 and 16 weeks old.
Results
As shown in Table 1, compared with control mice at 8 weeks old, body weight, mean blood pressure, fasting blood glucose, triglycerides, AST, and blood urea nitrogen levels were significantly elevated in Ctrl-Apt-treated KKAy/Ta diabetic mice at 12 weeks old. In addition, body weight, mean blood pressure, and blood urea nitrogen levels were significantly higher in Ctrl-Apt-treated KKAy/Ta diabetic mice at 16 weeks old than those in control mice at 8 weeks old, whereas fasting blood glucose in Ctrl-Apt-treated mice and AST in RAGE-Apt-treated mice at 16 weeks old were significantly increased. There were no significant differences of these parameters between Ctrl-Apt-treated mice and RAGE-Apt-treated mice at both 12 and 16 weeks old, except for triglycerides; RAGE-Apt treatment significantly increased triglycerides in KKAy/Ta diabetic mice at 12 weeks old, whereas it decreased triglycerides in diabetic mice at 16 weeks old compared with Ctrl-Apt. Compared with control mice at 8 weeks old, serum adiponectin levels were significantly decreased in both Ctrl-Apt-treated and RAGE-Apt-treated KKAy/Ta diabetic mice at 16 weeks old, but not at 12 weeks old. Although adiponectin levels were lower in RAGE-Apt-treated KKAy/Ta mice at 12 weeks old compared with Ctrl-Apt-treated mice at the same age, there was no significant difference of circulating adiponectin levels between the two group mice at 16 weeks old (Table 1).
NAG activities in the urine were significantly elevated in Ctrl-Apt-treated KKAy/Ta diabetic mice at 12 and 16 weeks old compared with control mice at 8 weeks old, both of which were reduced by the treatment with RAGE-Apt (Table 1). Although cystatin C levels in KKAy/Ta mice at 12-week and 16-week old were significantly higher than those of control mice at 8-week-old, there was no significant difference of cystatin C levels between Ctrl-Apt- and RAGE-Apt-treated KKAy/Ta mice at 12-week and 16-week old (Table 1).
As shown in Figure 1(a) to (c), compared with Ctrl-Apt-treated mice at 12 weeks old, renal CML, RAGE, and NADPH oxidase-driven superoxide generation levels were significantly decreased in RAGE-Apt-treated mice at 12 weeks old. There were no significant differences of CML, RAGE, and NADPH oxidase-driven superoxide generation levels between Ctrl-Apt- and RAGE-Apt-treated KKAy/Ta mice at 16 weeks old.
As shown in Figure 1(d), compared with 8-week-old mice, glomerular area was significantly increased in Ctrl-Apt-treated KKAy/Ta mice at 12 and 16 weeks old. Furthermore, both glomerular and cortical interstitial areas in 16-week-old RAGE-Apt-treated mice were significantly smaller than those of Ctrl-Apt-treated mice at the same age (Figure 1(d) and (e)). In addition, glomerular and cortical interstitial areas in RAGE-Apt-treated 12-week-old diabetic were slightly, but not significantly, decreased compared with those in Ctrl-Apt-treated mice at the same age (p = 0.41 for glomerular areas and p = 0.25 for interstitial areas). Intra-tubular area did not differ among the three groups.
Adiponectin expression in adipose tissues of RAGE-Apt-treated mice at 12 weeks old were significantly higher than that in Ctrl-Apt-treated mice at the same age (Figure 2(a)). Although adipose adiponectin levels in Ctrl-Apt- and RAGE-Apt-treated KKAy/Ta mice at 16 weeks old were higher than those in 8-week-old mice, there was no significant difference of adipose adiponectin between the two groups (Figure 2(a)).
HOMA-IR was significantly increased in 16-week-old Ctrl-Apt-treated mice compared with control mice at 8 weeks old, which was reduced by the treatment with RAGE-Apt (Figure 2(b)). Although HOMA-IR was slightly higher in RAGE-Apt-treated mice at 12 weeks old compared with Ctrl-Apt-treated mice at the same age, these values did not differ from the HOMA-IR of 8-week-old mice (Figure 2(b)).
Conclusion
AGEs have been shown to induce oxidative stress generation, apoptotic cell death, inflammatory and fibrotic reactions in human cultured proximal tubular cells via the interaction with RAGE.16–19 Indeed, RAGE-Apt or antibodies raised against RAGE have been reported to block these harmful effects of AGEs on proximal tubular cells.18,19 However, the pathological role of AGE-RAGE axis in tubulointerstitial damage in type 2 diabetic nephropathy has not been fully elucidated. In order to investigate the time-course effects of RAGE-Apt on tubular damage and insulin resistance in type 2 diabetic animals, we used here 8-week-old prediabetic KKAy/Ta mice as a control group. Furthermore, to examine the effects of RAGE-Apt on AGE-RAGE axis and adiponectin and subsequent tubular damage in type 2 diabetic animals, we investigated renal CML, RAGE, NADPH oxidase activity, and adipose adiponectin expression in KKAy/Ta mice at both 12-week-old and 16-week-old. In this study, we found for the first time that RAGE-Apt treatment significantly inhibited the increase in urinary NAG activities, a marker of tubulointerstitial injury and dysfunction 20 in obese type 2 diabetic mice with insulin resistance, which was associated with reduction of CML, RAGE, and NADPH oxidase-driven superoxide generation in the kidneys of KKAy/Ta diabetic mice at 12 weeks old. Injury and apoptosis of proximal tubular cells could contribute to tubular atrophy, tubulointerstitial fibrosis and atubular glomeruli, which are prognostic histological markers to predict the progression of renal dysfunction in diabetic patients.9,21,22 Given that urinary NAG activity could also predict renal prognosis in patients at early stage of diabetic nephropathy, 23 our present study suggests that inhibition of the AGE-RAGE axis in tubulointerstitial areas by RAGE-Apt may be a novel therapeutic strategy for treatment of diabetic nephropathy.
Anti-RAGE neutralizing antibodies have already been shown to play a protective role against experimental diabetic nephropathy.11,12 However, aptamers have several advantages over antibodies for blocking the deleterious function of target protein.24,25 Compared with antibodies, aptamers can be easily selected with relatively low production costs. When used in an in vivo therapeutic application, aptamers can more efficiently penetrate into various tissues with less immunogenicity over antibodies.24,25 These findings may suggest the clinical feasibility of RAGE-Apt for treatment of diabetic nephropathy.
RAGE-Apt reduced urinary albumin excretion levels by half in KKAy/Ta diabetic mice at 16 weeks old, but its effects were not statistically significant. That meant that the present data did not provide evidence against null-hypothesis. However, in the present study, all the KKAy/Ta mice at 12-week-old were sacrificed for immunohistological analyses. Therefore, we could not evaluate the serial change of urinary albumin over time in each animal. In addition to this, small sample size may limit the statistical power of the findings. On the other hand, histological examinations revealed that compared with 8-week-old mice, glomerular areas were significantly increased in Ctrl-Apt-treated KKAy/Ta mice at 12 and 16 weeks old, the latter of which was significantly attenuated by the treatment with RAGE-Apt. Moreover, cortical interstitial area in RAGE-Apt-treated KKAy/Ta diabetic mice at 16 weeks old was significantly smaller than those in Ctrl-Apt-treated mice at the same age. Given these beneficial effects of RAGE-Apt on histological alterations in the kidneys of KKAy/Ta mice in association with the reduction of CML, RAGE, and NADPH oxidase-derived superoxide generation in both glomeruli as well as tubulointerstitial areas, it is conceivable that inhibition of glomerular AGE-RAGE axis by RAGE-Apt may also play a protective role in our animal models. It would be interesting to examine whether long-term treatment with RAGE-Apt could prevent the increase in urinary albumin excretion, glomerular sclerosis, and tubulointerstitial fibrosis in KKAy/Ta mice.
In this study, we found that CML, one of the well-characterized AGEs, 13 RAGE, and NADPH oxidase-mediated superoxide generation levels were significantly reduced by RAGE-Apt in 12-week-old KKAy/Ta mice. We have previously shown that neutralizing antibody raised against RAGE or an anti-oxidant N-acetylcysteine inhibits the AGE-induced oxidative stress generation and RAGE expression in human cultured proximal tubular cells. 18 Furthermore, renal levels of CML are decreased in RAGE-deficient diabetic mice. 26 These observations suggest that the AGE-RAGE-NADPH oxidase-evoked superoxide generation could enhance the CML formation and RAGE expression in the diabetic kidneys, whose positive feedback loop was blocked by the treatment with RAGE-Apt. Prognostic value of urinary excretion levels of AGEs for progression of diabetic nephropathy was lost after adjustment for baseline urinary activity of NAG, 27 further supporting the clinical relevance of AGE-RAGE axis activation in tubular damage and subsequent deterioration of renal function in diabetic nephropathy.
In the present study, we found that RAGE-Apt increased adipose tissue adiponectin expression at 12-week-old KKAy/Ta mice and ameliorated insulin resistance evaluated by HOMA-IR at 16-week-old KKAy/Ta mice. Although calculation of HOMA-IR is human-based and that basal concentrations of glucose and insulin are not the same in rodents, it is widely used as a proxy of insulin resistance in rodents as well and an accepted surrogate for estimating insulin resistance. 28 Since we, along with others, have found that (1) blockade of the AGE-RAGE axis improves glycemic control in fructose-fed rats, an animal model of insulin resistance and that (2) insulin sensitivity and serum adiponectin levels are increased in RAGE-deficient mice compared with wild-type mice,29,30 inhibition of the AGE-RAGE axis in the adipose tissues may also be a therapeutic target for insulin resistance in obese type 2 diabetes. A ligand of peroxisome proliferator-activated receptor-gamma has been shown to reduce triglycerides and counteract insulin resistance, thereby resulting in amelioration of tubular damage in obese type 2 diabetic animals. 31 Moreover, insulin resistance is associated with tubular damage in elderly individuals. 32 These observations suggest that beneficial effects of RAGE-Apt on triglycerides and HOMA-IR may partly contribute to the protection against tubular injury in KKAy/Ta mice.
A major limitation to this study is the lack of age-matched non-diabetic control group. Indeed, we used a spontaneous model of type 2 diabetes, and comparisons of data were performed between RAGE-Apt- and Ctrl-Apt-treated mice at 12 and 16 weeks old as well as between both groups at each time point and 8-week-old prediabetic mice. From this experimental design, it is impossible to exclude that effects of RAGE-Apt observed here may not be specific to diabetes. In other words, effects of RAGE-Apt on age-related changes may affect the present findings. Second, RAGE-Apt significantly reduced CML, RAGE, and NADPH-oxidase-derived superoxide generation in the kidneys of diabetic mice at 12 weeks old, but not at 16 weeks old. We did not know the reason for the time-varying effects of RAGE-Apt on AGE-RAGE axis. In any case, suppression of renal AGE-RAGE axis by RAGE-Apt in diabetic mice at 12 weeks old may contribute to the beneficial effects of RAGE-Apt on histological alterations in the kidneys of diabetic mice at 16 weeks old. In addition, we did not measure the effects of RAGE-Apt on tubular apoptosis in this model. It would be helpful to examine whether RAGE-Apt could attenuate tubular apoptosis in KKAy/Ta diabetic mice. Further biochemical data that could support the histological findings would strengthen the present results.
Footnotes
Author contributions
S. Yamagishi conceptualized and designed the study; acquired, analyzed, and interpreted data; and drafted the manuscript; and he takes responsibility for the integrity of the data and the accuracy of the data analysis. A. Sotokawauchi, T. Matsui, Y. Higashimoto, Y. Nishino, and Y. Koga acquired, analyzed, and interpreted data. M. Yagi critically revised the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by Grants-in-Aid for Scientific Research (Grant Number 17K08968 (SY) and 19K16387(AS)) from the Ministry of Education, Culture, Sports, Science and Technology, Japan.