
Abstract
Obesity-related euglycaemic insulin resistance clusters with cardiometabolic risk factors, contributing to the development of both type 2 diabetes and cardiovascular disease. An increased thrombotic tendency in diabetes stems from platelet hyperactivity, enhanced activity of prothrombotic coagulation factors and impaired fibrinolysis. Furthermore, a low-grade inflammatory response and increased oxidative stress accelerate the atherosclerotic process and, together with an enhanced thrombotic environment, result in premature and more severe cardiovascular disease. The disruption of circadian cycles in man secondary to chronic obesity and loss of circadian cues is implicated in the increased risk of developing diabetes and cardiovascular disease. Levels of melatonin, the endogenous synchronizer of circadian rhythm, are reduced in individuals with vascular disease and those with deranged glucose metabolism. The anti-inflammatory, antihypertensive, antioxidative and antithrombotic activities of melatonin make it a potential therapeutic agent to reduce the risk of vascular occlusive disease in diabetes. The mechanisms behind melatonin-associated reduction in procoagulant response are not fully known. Current evidence suggests that melatonin inhibits platelet aggregation and might affect the coagulation cascade, altering fibrin clot structure and/or resistance to fibrinolysis. Large-scale clinical trials are warranted to investigate the effects of modulating the circadian clock on insulin resistance, glycaemia and cardiovascular outcome.
Keywords
Introduction
Diabetes mellitus is a major health problem with almost 400 million adults with type 2 diabetes mellitus (T2DM) worldwide.1–3 A physiological phenomenon of insulin resistance results from circannual weight gain, which confers resistance to hypothermia and infections and is thus protective in overwintering and migrating animals.4–8 However, in man, the loss of regulation of body weight by the seasonal needs and artificial lighting leads to a loss of seasonal adjustment in adiposity, transforming what is a circannual phenomenon of insulin resistance in overwintering animals to a circadian pattern in humans.9–12 Fat-loaded adipose tissue provokes insulin resistance and contributes to metabolic syndrome through the dysregulated production of free fatty acids and pro-inflammatory adipokines affecting metabolic tissues (skeletal muscle and liver) and modifying inflammatory responses.12–14 The net result is a hyperglycaemic and pro-inflammatory environment that in turn accelerates vascular pathology, explaining the close association between diabetes and vascular ischaemic events.6,8
Cardiovascular disease (CVD) remains the main cause of morbidity and mortality in patients with T2DM, with around three-quarter of patients dying of CVD complications. 15 In addition to increased risk of first vascular events, T2DM subjects have worse outcomes following vascular ischaemia compared to subjects with normal glucose metabolism. 16 One of the key links between T2DM and CVD is insulin resistance, which is associated with clustering of vascular risk factors including deranged glucose metabolism, increased very-low-density lipoprotein triglyceride, decreased high-density lipoprotein cholesterol and hypertension.17–26
While insulin resistance has been attributed to lifestyle changes and the increase in obesity worldwide, understanding the mechanistic pathways underpinning insulin resistance will help to develop more effective management strategies. In particular, the production of the endogenous synchronizer of circadian rhythms, melatonin, is reduced in several cardiovascular diseases and in insulin-resistant subjects.27–35 The anti-inflammatory, antihypertensive, antioxidative activity of melatonin is suggested to favourably influence the development of atherothrombotic disease in diabetes.36–38 This review will use a foundation of circadian cardiometabolic physiology to summarize mechanisms for increased atherothrombotic events and evaluate the potential therapeutic opportunities for melatonin as an anti-atherothrombotic agent, particularly in the context of obesity-related insulin resistance.
A search of PubMed (MEDLINE) was performed starting from the year 1980 until the present, using the following terms either singly or in combination: ‘insulin resistance’, ‘type 2 diabetes’, ‘prediabetic state’, ‘cardiovascular disease’, ‘atherosclerosis’, ‘atherosclerotic plaque’, ‘inflammation’, ‘platelet aggregation’, ‘endothelial dysfunction’, ‘circadian rhythm’, ‘circadian misalignment’, ‘melatonin’, ‘thrombosis’, ‘coagulation’, ‘atherothrombosis’, ‘fibrinogen’, ‘plasminogen activator inhibitor - 1’. Research papers were considered on the basis of their relevance to diabetic atherothrombosis and potential atheroprotective effect of melatonin. Articles that covered circadian rhythm and diabetes, dysglycaemia or cardiovascular disease were selected for inclusion in this review.
Mechanisms for increased atherothrombotic events in T2DM
Insulin resistance is associated with endothelial dysfunction, a pro-inflammatory and a prothrombotic environment, which are all key to the development of atherothrombotic disease in T2DM (Figure 1).39–44
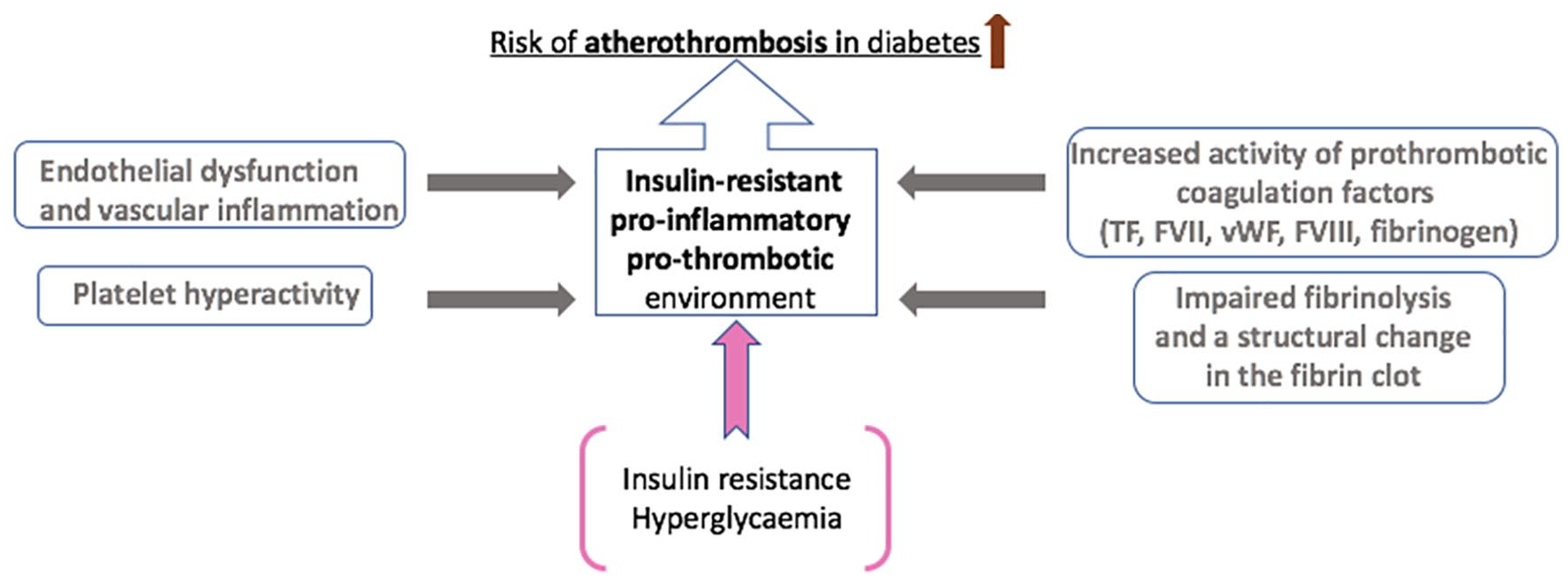
Mechanisms for increased atherothrombotic events in diabetes. The underlying mechanisms for increased atherothrombotic risk in diabetes are complex and multifactorial. In addition to platelet hyperactivity following endothelial damage as a result of a low-grade chronic inflammatory response, increased thrombotic tendency primarily stems from enhanced activity or raised plasma levels of prothrombotic coagulation factors, including TF (tissue factor), FVII (Factor VII), vWF (von Willebrand factor), FVIII (Factor VIII) and fibrinogen. This results in fibrin networks, which form the backbone of the blood clot, that are compact with increased resistance to fibrinolysis. Insulin resistance and hyperglycaemia enable these underlying mechanisms and directly contribute to an insulin-resistant, pro-inflammatory and prothrombotic environment in diabetes, increasing the risk of atherothrombosis.
Endothelial dysfunction and vascular inflammation
Insulin resistance is associated with a cluster of risk factors that result in endothelial dysfunction, a key abnormality that contributes to the atherothrombotic process (reviewed in Sena et al. 71 ). Briefly, endothelial dysfunction causes endothelial cells to express adhesion molecules, thus attracting inflammatory cells, and also increases inter-cellular permeability resulting in accumulation of lipoproteins and inflammatory cells within the vessel wall. Lipid particles migrating to vessel wall are oxidized into highly atherogenic molecules that are taken up by inflammatory cells to form foam cells. An accumulation of foam cells forms the fatty streak, the earliest abnormality in the atherosclerotic process. This is followed by deposition of collagen, gradually resulting in the development of the mature atherosclerotic plaque. Enhanced production of cytokines and growth factors by these inflammatory cells leads to smooth muscle migration and proliferation in the intima, which further contributes to the inflammatory process.42,49,54,55
Increased inflammatory cytokines, including tumour necrosis factor α (TNFα) and interleukin (IL)-6, impairs production of a vasodilatory molecule NO (nitric oxide) and also modulates activation of PI3K/Akt pathways further accelerating atherothrombosis in diabetes.72–79 In addition, the activation of the pro-inflammatory nuclear transcription factor nuclear factor-kappa B is permitted by the loss of NO, increasing the expression of adhesion molecules, cytokines and chemokines, creating a vicious cycle.50,52 Finally, increased production of advanced glycation end product (AGE) and formation of reactive oxygen species (ROS) diminishes endothelial-derived NO and leads to enhanced synthesis of vasoconstrictor prostanoids and endothelin.49,80–82
Role of circadian rhythm in T2DM and increased thrombotic environment
The underlying mechanisms for increased thrombosis risk in diabetes subjects are complex and multifactorial. Both the cellular and protein arms of coagulation in diabetes are affected leading to a prothrombotic environment. Platelet activation is increased in diabetes and a key mechanism appears to be reduced NO production, an important inhibitor of platelet activation, secondary to endothelial dysfunction.46,83–96 Furthermore, under conditions of hyperglycaemia and insulin resistance, chronic low-grade systemic inflammation drives the haemostatic system towards a prothrombotic state.39,40–44 Plasma levels of a number of procoagulant factors are elevated in diabetes patients, including tissue factor, factor VII, von Willebrand Factor, factor VIII and fibrinogen, increasing thrombotic tendency.45,49,59–65,80 In addition, diabetes is associated with reduced concentration of antithrombotic factors, such as antithrombin III and protein C, 83 further adding the prothrombotic milieu. Moreover, the fibrinolytic system is directly affected in diabetes secondary to elevated levels of plasminogen activator inhibitor (PAI)-1 and impaired plasminogen to plasmin conversion, together with reduced enzyme activity. 16 Interestingly, the EuroClot study involving healthy subjects reported an association between clustering of cardiometabolic risk factors and suppression of fibrinolytic activity, making modulation of hypofibrinolysis in diabetes a possible target to reduce atherothrombotic disease. 97 Hypofibrinolysis has been recently reported to be an independent predictor of cardiovascular mortality in individuals with acute coronary syndrome treated with modern antithrombotic therapies. 98 In summary, inflammation-mediated endothelial dysfunction, platelet hyperactivity, increased activation of pro-coagulation factors and impaired fibrinolysis represent a wide spectrum of biological targets with most of them exhibiting circadian variation, thus implicating loss of circadian rhythmicity in the increased thrombotic tendency in diabetes.99–103
The constant exposure to food and the loss of the dark/night stimulus translate into loss of environmental control of circadian rhythmicity, which is implicated in the above-mentioned processes and thus increased risk of developing both diabetes and cardiovascular disease.104–106 The master circadian pacemaker in the suprachiasmatic nucleus generates diurnal circadian rhythms secondary to cyclical expression of clock-controlled genes in response to light and food intake. 107 Clock genes, the Clock and Bmal1, encode transcriptionally activating proteins of the clock system in response to light, whereas Per 1-3 and Cry 1 and 2 genes encode transcriptionally inhibiting proteins of the clock system. 35 Rudic et al. 108 reported that an adequate insulin response depends on normal function of the circadian clock. Indeed, Clock mutant Bmal1−/− mice develop a diabetes phenotype with metabolic abnormalities, including impaired gluconeogenesis, hyperglycaemia and hypercholesterolaemia. 108 Mutations in the Clock gene have also been associated with the metabolic syndrome and vascular dysfunction in man, implicating circadian disruption in the development of both diabetes and associated cardiovascular disease.109–111
In human subjects, data linking circadian misalignment with development of T2DM and adverse cardiovascular events comes either from epidemiological studies in shift workers or from short-term sleep deprivation studies (Table 1). Factors opposing aligned circadian rhythmicity are proposed to underlie higher rates of cardiovascular disease events in shift workers when compared to day workers.112–117 Women of the Nurses’ Health Study cohort with 6 years or more of rotating shift work experience were 1.51 [95% confidence interval (CI): 1.12–2.03] more likely to develop coronary heart disease compared to women who had never been shift workers. 118 A systematic review conducted in 2012 reported that shift work was associated with 41% increased relative risk of coronary events. 119 Furthermore, circadian misalignment has been shown to have a negative impact on the predictors of cardiovascular disease risk, including pro-inflammatory markers, such as serum IL-6 and C-reactive protein (CRP), and was also associated with hypercoagulability.120,121 There is, however, evidence of metabolic and cardiovascular disease markers being improved when shift work schedules are adapted to biological rhythmicity. 122
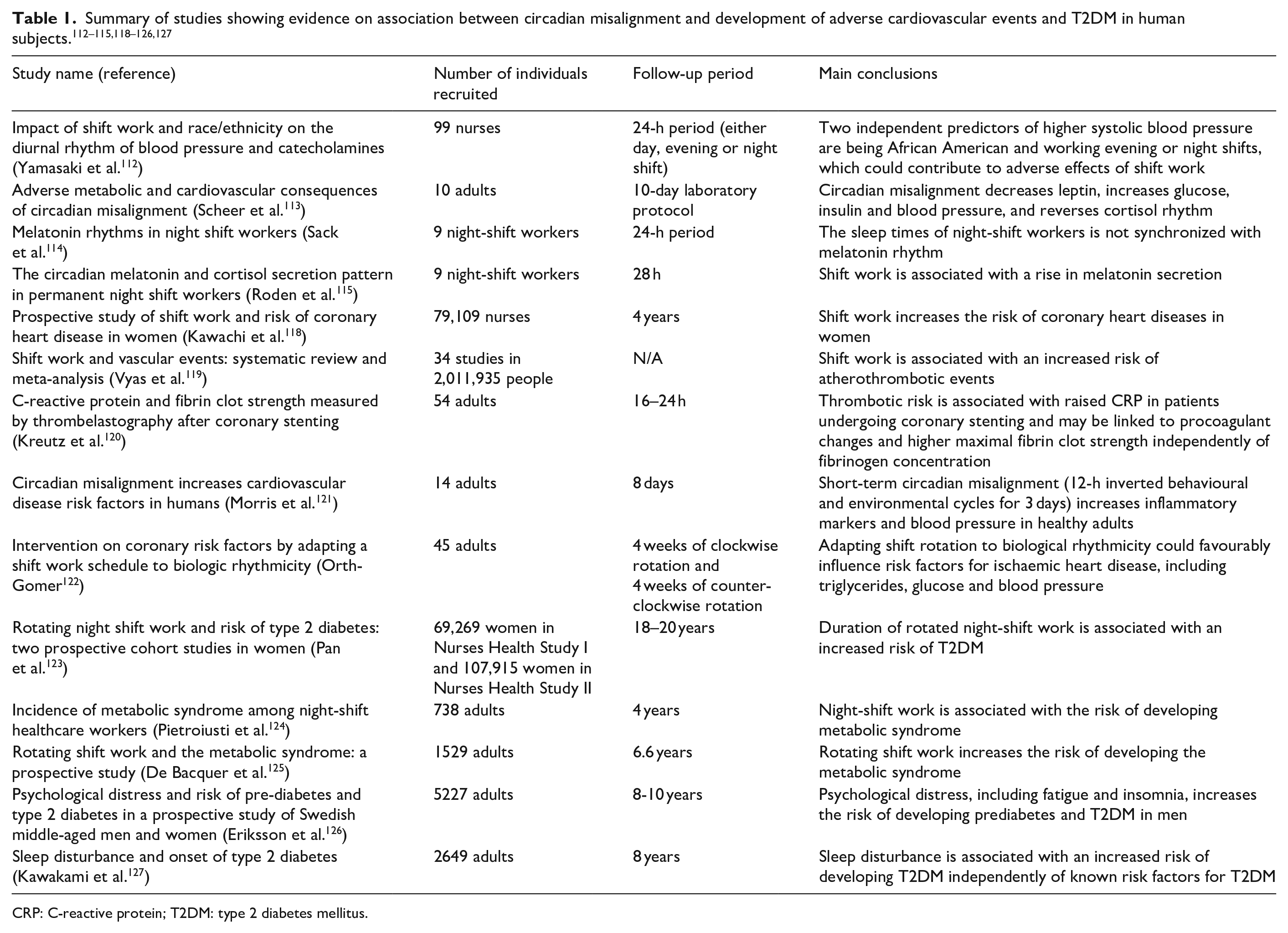
CRP: C-reactive protein; T2DM: type 2 diabetes mellitus.
Epidemiological evidence also indicates associations between shift work and development of T2DM.27,123 An important prospective clinical study, including 402 night-shift workers and 336 daytime workers followed up for a median of 4 years, identified a five-fold increased risk of developing T2DM/obesity in night-shift workers. 124 In addition, a prospective study on a cohort of 1529 workers reported a 77% higher incidence of the metabolic syndrome in shift workers. 125 Misalignment of circadian rhythm and sleep disorders have been associated with the development of metabolic syndrome and prediabetes, correlated with an elevated risk of T2DM.27,126 There is also mounting evidence to demonstrate a close association between sleep deprivation, insomnia, restless leg syndrome and T2DM. 128 Poor sleep quality is associated with increased sympathetic activity, decreased leptin levels and elevated ghrelin levels among other disturbances, leading to a number of metabolic abnormalities, including glucose intolerance.127,129,130 Moreover, loss of circadian clock synchronization has been suggested as one mechanism for increased thrombotic tendency in diabetes. 131 That view is supported by in vitro studies demonstrating that the Clock/Bmal heterodimers regulate PAI-1 gene expression. 132 The involvement of Clock in the regulation of PAI-1 was also reported in the setting of obesity, supporting the link between obesity, diabetes and CVD through circadian regulation and PAI-1 gene expression. 133
The above evidence raises the possibility of using circadian rhythm modulators to ‘reset’ normal circadian physiology, in turn reducing metabolic abnormality and vascular risk. 134
Melatonin and the circadian rhythm
The pineal gland is essential as a rhythmic synchronizer, connecting the nervous and the endocrine systems.135,136 The retinohypothalamic-pineal system becomes activated by norepinephrine with the onset of darkness, leading to melatonin release (Figure 2).36,135,136 Activity of the central pacemaker, being regulated by the humoral system, triggers peripheral circadian oscillators, controlling their rhythmicity to coordinate metabolic activity throughout the body.136–138 These peripheral pacemakers share similar organization at the molecular level and regulate circadian variation in local transcription clusters.138,139 Metabolically active organs including skeletal muscles, liver and adipose tissue display circadian variability in the expression of clock genes and genes that are involved in hormone synthesis and encode proteins participating in key metabolic events.139,140 Although animal studies demonstrate that environmental stimuli, such as light exposure, regulate the phase of the master clock, experiments of restricted feeding show that the phase of circadian expression in mouse liver could be inverted without affecting central clock gene expression.141–143 Such desynchrony, whereby the peripheral pacemakers are uncoupled from the master clock in the suprachiasmatic nucleus, may worsen circadian-related metabolic disorders. 144
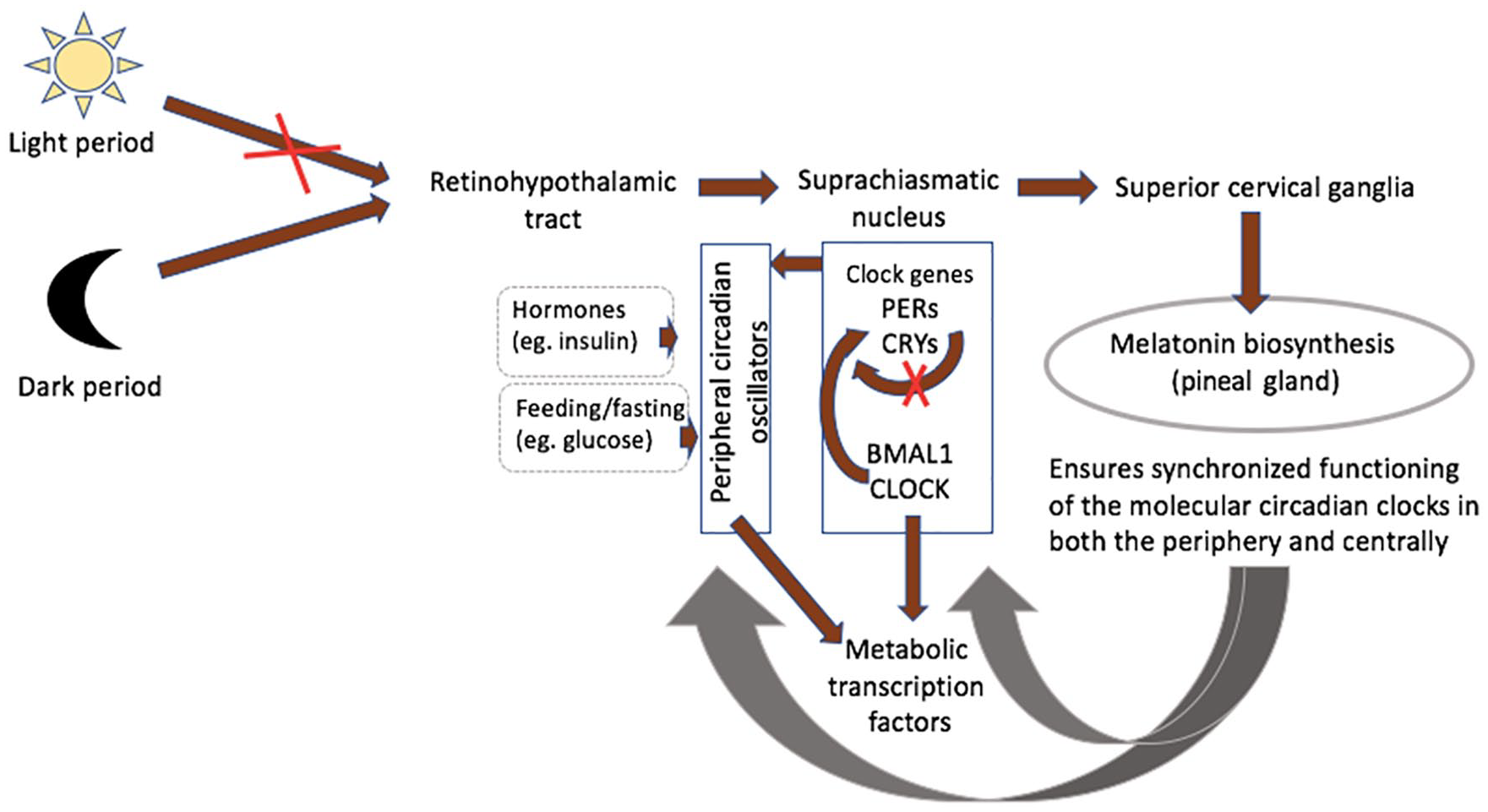
Melatonin an internal synchronizer of circadian biological rhythmicity. The retinohypothalamic-pineal system becomes activated with the onset of darkness, leading to the transmission of neural signals from the suprachiasmatic nucleus (central pacemaker) to the superior cervical ganglia. Noradrenalin release from postganglionic fibres stimulates synthesis of melatonin by pinealocytes. Clock genes encoding transcriptionally activating or inhibiting proteins of the clock system in response to variation in light exposure are the Clock and Bmal1, PERs and CRYs, respectively. Activity of the central pacemaker triggers peripheral circadian oscillators, which are also influenced by local factors, including hormones and glucose. Metabolically active organs including skeletal muscles, liver and adipose tissue display circadian variability in the expression of metabolic transcription factors. Melatonin prevents desynchrony, whereby the peripheral pacemakers are uncoupled from the master clock in the suprachiasmatic nucleus, which may worsen circadian-related metabolic disorders.
Melatonin plays an essential role in the synchronized functioning of the molecular circadian clocks in both the periphery and centrally in the suprachiasmatic nucleus.36,137,146,147 Its circadian rhythm-entraining effects are mediated via melatonin receptors MT1 and MT2, which have been implicated in jet lag phenomenon and in the context of sleeping disorders.148–150 Animal studies report reductions in visceral adiposity, glucose, insulin and triglycerides following melatonin administration.36,137,151–154 Although the evidence on the effect of melatonin is limited in human individuals and in the context of insulin resistance, decreased melatonin production found in patients with T2DM, prediabetes and cardiovascular disease suggests a protective role for this hormone in insulin resistance and atherosclerosis.27–35
Role of melatonin in vascular complications in T2DM
Evidence gathered in the last 20 years indicates that melatonin influences multiple aspects of cardiometabolic function.36,137,151–154 Strong evidence comes from studies demonstrating that genetic variants in the melatonin receptor as a result of single nucleotide polymorphisms are related to obesity and are associated with atherosclerosis and the risk of myocardial infarction (MI). Genome-wide association studies indicate a causal relationship between one of the single nucleotide polymorphisms of melatonin receptor MTNR1B and reduced glucose-regulating ability. Furthermore, studies on hypercholesterolemic rats indicated that melatonin leads to lowering of total cholesterol, low-density lipoprotein (LDL) and very-low-density lipoprotein cholesterol levels, suggesting it may protect from atherosclerosis. 155 In addition, animal studies provide evidence on the antioxidant and anti-inflammatory actions of melatonin, which attenuates thermal-induced activation of blood clot formation.156,157 These observations together with studies demonstrating low melatonin levels in individuals with coronary artery disease, insulin resistance and T2DM led to the speculation that melatonin has a role in vascular pathology.27–35 Therefore, melatonin treatment may prove to be an effective strategy in the management of atherothrombotic disease, particularly in high-risk subjects with deranged circadian rhythm such shift workers. Experimental work provided evidence that melatonin supplementation improves insulin resistance induced by disruption of internal circadian rhythms. 158 In addition to preclinical evidence suggesting exogenous melatonin could also reduce leptin resistance, hyperinsulinaemia and hyperglycaemia in animal models of obesity and metabolic syndrome, melatonin supplementation has also been shown to be associated with significant reduction in HbA1c in a small group of patients with diabetes and insomnia.159–161 More recently, studies show that the low levels of melatonin secretion predicts the onset of T2DM in women and that adequate melatonin secretion is thought to reduce the incidence of T2DM.34,162
Antihypertensive and antioxidant properties of melatonin: potential impact on the atherosclerotic process
Studies have shown associations between melatonin secretion and reduction in night-time systolic blood pressure.36,163 Obayashi et al., 163 however, did not find a significant relationship between melatonin excretion and diastolic blood pressure, suggesting the relationship is mainly with systolic blood pressure by mechanisms that are not entirely clear.
Oxidative stress plays a central role in the atherosclerotic process, particularly in diabetes. 44 The antioxidant properties of melatonin stem from its detoxifying characteristics, effective scavenging of reactive oxygen and nitrogen species and its ability to enhance antioxidative enzymes. 36 Studies have demonstrated that the rhythm of antioxidant defence is obliterated in pinealectomized rats and by light in humans. 36 In prothrombotic stroke models of ischaemia-induced oxidative damage, in vivo melatonin administered post-ischaemia exhibited a concentration-dependent protection mediated through a reduction in ROS production, associated with decreased infarct size. 164 Similarly, melatonin was shown to have a cardioprotective effect against oxidative injury in the ischaemic/reperfused heart.155,165,166 Decreased serum melatonin and an increase in oxidative stress in patients with MI may indirectly suggest a protective effect of melatonin against cardiovascular disease in man.167–169 Finally, subjects with elevated levels of LDL, oxidation of which is implicated in the progression of atherosclerosis, have been reported to have low levels of melatonin. 153 Some studies, but not all, support prevention of LDL oxidation by melatonin, providing another potential mechanism of protection from vascular pathology.153,170
Melatonin deficiency augments sympathetic activity and influences atherosclerosis and thrombus formation
Nocturnal secretion of melatonin is reduced in subjects with coronary artery disease, and especially those with acute coronary syndrome (ACS), compared to healthy individuals, and in patients with unstable compared to stable angina.29,30,168,171–173 Impaired circadian biological rhythmicity and the lack of the blunting effect of melatonin on sympathetic activity lead to sympathetic activation, contributing to endothelial injury, platelet activation and predisposing vulnerable plaques to rupture.29,113 Increased sympathetic activity and activation of the coagulation cascade in the early morning might contribute to the well-described morning peaks in cardiovascular events in patients with coronary artery disease (CAD).28,173,174 In addition, increased sympathetic activity might affect production of a key inhibitor of fibrinolysis, PAI-1, thus contributing to hypofibrinolysis and increasing the risk of vascular events. 175
Urinary 6-sulfatoxymelatonin excretion, a urinary metabolite of melatonin serving as an index of its secretion, is inversely associated with arterial stiffness after adjusting for diabetes and hypertension. 176 While this does not prove causality, the association warrants further investigation. Previous work has also shown that melatonin has antithrombotic properties when administered at low doses before or during coronary occlusion, although the exact mechanisms remain to be elucidated. 177
Melatonin affects activated coagulation and platelet aggregation
A few animal studies have investigated the effect of melatonin on heat-induced inflammatory and coagulation responses. Platelet morphology and elevated levels of fibrinogen, fibrin degradation products, prothrombin activity and CRP following thermal injury were normalized after melatonin administration.156,157 Antithrombotic effects of melatonin were also investigated in randomized-controlled trials in healthy volunteers under acute psychosocial stress conditions. Emotionally triggered catecholamine-induced hypercoagulable state promotes thrombus formation following plaque rupture.178–181 Stress-induced increases in FVII:c and fibrinogen contribute to a hypercoagulable state; however, no statistically significant reductions in these factors were observed after melatonin administration, arguing against a beneficial role for melatonin during thrombotic vascular occlusion.181–183 In healthy young men, the administration of melatonin was associated with a significant reduction in FVIII:c and fibrinogen levels, whereas levels of FVII:c and D-dimer showed no change.
173
On the contrary, melatonin could attenuate elevations in
In addition to the protein arm of coagulation, melatonin is thought to influence circadian variation in platelet activity and several studies have suggested melatonin having direct effects on platelet function. Melatonin has been implicated in the inhibition of platelet aggregation, both spontaneous and induced, mainly through the cyclooxygenase-dependent pathway.184–189 In addition, some studies, but not all, suggest that melatonin elevates the apoptotic events in platelets.190,191 Although both of these effects of melatonin on platelets could potentially limit the process of thrombogenesis, the evidence behind them is not consistent and therefore warrants further exploration.
Conclusion and future directions
There is convincing evidence stemming from genetic and epidemiological studies to implicate deranged circadian rhythm in the development of both diabetes and cardiovascular disease. Although the evidence for melatonin as an agent that restores the deranged rhythm is limited in man, indirect evidence suggests a central role for melatonin in abnormal glucose metabolism and/or vascular pathology.
An increased thrombotic tendency in the setting of low-grade chronic inflammation primarily stems from platelet hyperactivity, enhanced activity of prothrombotic coagulation factors and impaired fibrinolysis. Anti-inflammatory and antioxidative properties of melatonin may favourably modulate the prothrombotic and inflammatory environment typically found in atherothrombotic disease.36–38 In particular, the antioxidant activity of melatonin may be atheroprotective in diabetes since hyperglycaemia-induced free radical formation increases oxidative stress, which predisposes to vascular pathology.49,192,193 A cardioprotective and antithrombotic effect of melatonin might also stem from a selective effect on the plasma levels of coagulation measures, including fibrinogen and FVIII:c, both of which are found at increased concentrations in diabetes and are associated with an elevated risk of coronary thrombotic events.84,173 In addition, the atheroprotective potential of melatonin comes from evidence of melatonin deficiency increasing the risk of cardiovascular events by augmenting sympathetic activity, which contributes to platelet activation and hypertension-induced injury to the endothelial cell layer. 29
The potential mechanisms of melatonin-mediated atheroprotection are summarized in Table 2. Studies to date suggest that melatonin should be explored as an agent to reduce insulin resistance and prevent diabetes and/or vascular disease, particularly in individuals with deranged circadian rhythm. An advantage of melatonin supplementation is the favourable side effect profile, potentially giving it an advantage over established therapies. 145 However, the effects of melatonin to prevent desynchrony are not one-sided and melatonin supplementation could affect endogenous serotonin concentrations and functions of pineal and hypothalamic systems, thus influencing appetitive, emotional and cognitive processes. 194 In addition, persistent exposure to melatonin could lead to continuous activation of both melatonin receptors, generating off-target as well as on-target effects. For example, activation of MT1 receptor could generate continuous effects on downstream pathways, such as PLC/DAG/PKC pathway. This in turn could impact neuronal firing and immune system response via a continuous inhibition of leukocyte rolling in the microvasculature. 195 In addition, the effects of melatonin metabolites have to be explored and evaluated, in particular, 6-hydroxymelatonin-induced oxidative DNA damage. 196 Therefore, more mechanistic studies are needed in different population of patients to not only further understand the role of melatonin in modifying the inflammatory and thrombotic environment that predisposes to vascular disease, but also consider the effect of melatonin and its metabolites on the variety of biological systems.
Mechanisms responsible for the cardioprotective effect of melatonin in atherothrombotic disease.

An increased thrombotic tendency in diabetes stems from platelet hyperactivity, enhanced activity of prothrombotic coagulation factors and impaired fibrinolysis.
Levels of melatonin, the endogenous synchronizer of circadian rhythm, are reduced in individuals with vascular disease and those with deranged glucose metabolism.
Anti-inflammatory, antioxidative, antihypertensive and antithrombotic activities of melatonin may favourably modulate the prothrombotic and inflammatory environment typically found in atherothrombotic disease, which make it a potential therapeutic agent to reduce the risk of vascular occlusive disease in diabetes.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.