
Abstract
Chronic diseases account for approximately 45% of all deaths in developed countries and are particularly prevalent in countries with the most sophisticated and robust public health systems. Chronic metabolic diseases, specifically lifestyle-related diseases pertaining to diet and exercise, continue to be difficult to treat clinically. The most prevalent of these chronic metabolic diseases include obesity, diabetes, non-alcoholic fatty liver disease, chronic kidney disease and cardiovascular disease and will be the focus of this review. Wnt proteins are highly conserved glycoproteins best known for their role in development and homeostasis of tissues. Given the importance of Wnt signalling in homeostasis, aberrant Wnt signalling likely regulates metabolic processes and may contribute to the development of chronic metabolic diseases. Expression of Wnt proteins and dysfunctional Wnt signalling has been reported in multiple chronic diseases. It is interesting to speculate about an interrelationship between the Wnt signalling pathways as a potential pathological mechanism in chronic metabolic diseases. The aim of this review is to summarize reported findings on the contrasting roles of Wnt signalling in lifestyle-related chronic metabolic diseases; specifically, the contribution of Wnt signalling to lipid accumulation, fibrosis and chronic low-grade inflammation.
Keywords
Introduction
The term ‘Wnt’ originated as a merging of homologous proteins in Drosophila (Wg) and mice (Int-1), described in the early 1980s1,2 and the term was finally coined by Nusse et al. in 1991. 3 Although Wnt signalling plays a role in ‘normal’ physiological functions such as embryology, publications on the Wnt family related to its functions in disease processes is rapidly increasing, making the Wnt signalling pathways particularly interesting to study. Wnt signalling consists of two major pathways: a canonical pathway involving β-catenin (β-catenin-dependent) and a non-canonical Wnt pathway that is β-catenin-independent. The canonical Wnt signalling pathway is disproportionately studied compared to the non-canonical Wnt signalling pathway; this may be due to the greater complexity of the non-canonical signalling pathway. Wnts are best known for their association with a number of embryonic processes and homeostasis of tissues. 4 Given the importance of homeostasis to metabolism, it stands to reason that Wnts also regulate metabolic activities.
The leading cause of death since the early 1900s has shifted from acute infectious disease to chronic disease; this shift is attributed to improved public health interventions as countries transition from developing to developed.5,6 Chronic diseases account for approximately 45% of all deaths in developed countries. 7 Chronic metabolic diseases (CMDs) are difficult to prevent and treat due to long latency periods, numerous risk factors, co-morbidities, increasing average age of older adults and so on. The most prevalent of these CMDs include obesity, diabetes, non-alcoholic fatty liver disease (NAFLD), chronic kidney disease (CKD) and cardiovascular disease (CVD). In particular, CMDs, such as CVD and type II diabetes mellitus, are particularly prevalent in countries with the most sophisticated and robust public health systems. 5 Wnts have also been implicated in a variety of other contexts including bone metabolism, respiratory diseases, the urogenital tract, cardiovascular/endocrine systems, and inflammation and fibrosis.8–12 Additionally, Wnts have been implicated in obesity, type II diabetes, NAFLD, CKD and CVD.10,13–16 This review will summarize the emerging evidence implicating a role for Wnt signalling in lifestyle-related CMDs, obesity, diabetes, NAFLD, CKD and CVD.
Classification of Wnt signalling pathways
Wnt signalling consists of two major pathways: a canonical pathway (Wnt/β-catenin) and a non-canonical pathway that is subdivided into the Wnt/Ca2+ and planar cell polarity (PCP) pathways (Figure 1). 17 Wnt3a is one of the most highly studied canonical members. With regard to canonical Wnt signalling, when Wnt is not bound to its receptor(s), the ‘destruction complex’ is constitutively active.18,19 The destruction complex is composed of axin, adenomatous polyposis coli (APC), glycogen synthase kinase 3 beta (GSK3β), casein kinase-1 (CK-1) and β-transducin repeat containing protein (β-TrCP).18,19 The active destruction complex phosphorylates β-catenin in a GSK3β-dependent manner resulting in ubiquitination and proteosomal degradation of β-catenin.18,19 When a canonical Wnt binds to the frizzled receptor (Fz) and its co-receptor lipoprotein receptor-related protein (LRP) 5/6, dishevelled (Dsh) is recruited and the destruction complex is inhibited, thus promoting the accumulation of non-phosphorylated β-catenin in the cytosol. As non-phosphorylated β-catenin accumulates intracellularly, it is translocated to the nucleus where it activates the T-cell factor (TCF)/lymphocyte enhancer factor (LEF) transcription factor families to regulate gene transcription. 18
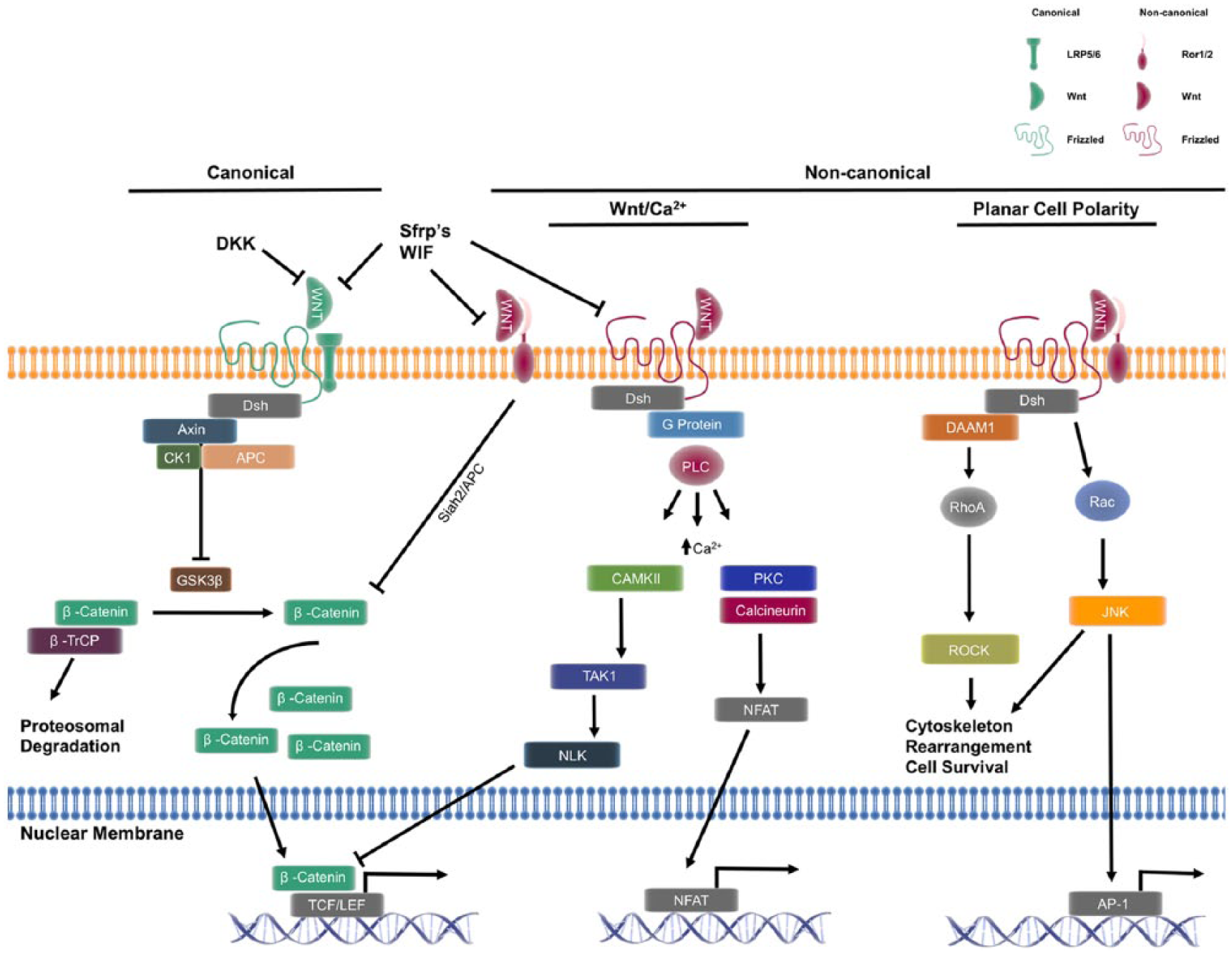
Wnt signalling pathways and their interactions. The canonical pathway is defined by the intracellular accumulation of β-catenin, and its resulting translocation to the nucleus where it regulates expression of target genes. The non-canonical pathway is defined by its β-catenin-independent actions ranging from intracellular signalling and expression of target genes. The non-canonical pathway can inhibit the canonical pathway at multiple levels. Activation of the Wnt/Ca2+ pathway has been demonstrated to affect gene transcription through NFAT, but can also inhibit β-catenin signalling through NLK. Downstream of the PCP pathway, RhoA regulates cytoskeletal rearrangement and cell survival; in addition, Rac and subsequent c-JNK activation positively regulates AP-1-dependent gene transcription. The complexity of Wnt signalling is compounded by the multiple extracellular factors, and co-receptors capable of influencing both canonical and non-canonical signalling.
The hallmark of the non-canonical signalling pathway is its β-catenin-independent actions. 17 The diversity of receptors and co-receptors makes non-canonical Wnt signalling highly context specific, depending on cell type and receptor availability, ultimately making evaluation of non-canonical signalling complicated. Of the 19 members of the Wnt family of glycoproteins, Wnt5a is one of the most highly studied non-canonical members. Non-canonical Wnts bind to Fz as well as a variety of co-receptors all of which contain a cysteine-rich binding domain. Such co-receptors include receptor tyrosine kinase (Ryk), tyrosine-protein kinase transmembrane receptor 1 or 2 (Ror1/2) and collagen triple helix repeat containing protein 1 (CTHRC1). 17 Unique combinations of these receptor/co-receptor/ligand complexes have been suggested to play a role in the downstream effects by a given Wnt ligand. 20 For example, CTHRC1 is a co-receptor that has been demonstrated to selectively activate the PCP pathway by stabilizing the Wnt5a/Ror2 receptor complex. 21
Beyond the receptor/co-receptor/ligand interactions at the cell surface, there is increasing evidence that non-canonical Wnt signalling interacts with other signalling pathways. Park et al. 22 recently elucidated a novel interaction of non-canonical PCP signalling and the Hippo-YAP/TAZ signalling pathway, demonstrating that YAP/TAZ signalling is, in fact, part of an ‘alternate Wnt-YAP/TAZ signalling axis’ that is responsible for many of the biological functions of non-canonical Wnt signalling. Furthermore, non-canonical signalling has been shown to inhibit canonical signalling by multiple mechanisms.22,23 One such mechanism involves increased destruction of β-catenin through a GSK3β-independent mechanism. 23 Another mechanism involves interaction with the Hippo-YAP/TAZ signalling pathway, increasing secretion of canonical signalling inhibitors (such as DKK1, WNT5A/B and others). 22
Compared to the canonical pathway, much less is known about the non-canonical pathway. Interestingly, Van Amerongen and Nusse 24 have proposed to do away with the binary classification of Wnt signalling in favour of an integrated Wnt signalling pathway, in part due to the complexity of canonical and non-canonical pathway cross-talk. Also, in general, the prototypical representation of signal transduction pathways as cause and effect is out dated. 24 For this review, the general classifications of canonical and non-canonical will be presented to be consistent with past literature and the apparent common effects of over activated canonical or non-canonical signalling and each with the ability to inhibit the other.
Characterization of Wnt proteins
Wnt proteins are approximately 350–400 amino acids in length with a conserved cysteine-rich binding domain consisting of 23–24 cysteine residues. 4 This cysteine-rich binding domain permits Wnt proteins to bind many different receptors described above. As mentioned, Wnt proteins have been classified into canonical (Wnt1, 2, 3, 8a, 8b, 10a and 10b) and non-canonical (Wnt4, 5a, 5b, 6, 7a, 7b and 11) based upon downstream signalling effects (β-catenin dependent or independent, respectively). Wnt proteins are highly insoluble, hydrophobic and classically described as acting in an autocrine/paracrine manner.18,25,26 Accounting for their hydrophobic properties is a high degree of post-translational palmitoylation and glycosylation, which are essential for secretion and function, respectively.4,27 These post-translational modifications are important for docking to extracellular lipoprotein particles allowing them to travel systemically.28,29
Although Wnts are described to act in both autocrine and paracrine manners, Wnt proteins are detectable in serum and levels correlate with chronic diseases such as atherosclerosis, obesity and rheumatoid arthritis.14,16,30 Furthermore, endogenous Wnt sequesters, such as secreted frizzled-related proteins (sFRP), which bind to Wnts and prevent their actions, correlate negatively with the aforementioned diseases.31–33 Altogether, it is interesting to suspect that Wnts have distinct endocrine functions, as well as autocrine and paracrine functions. Additional research in this area is warranted, particularly in the context of CMDs.
Wnts as regulators of metabolism
The perpetuation of chronic low-grade inflammation is a common denominator in CMD.34–36 Wnts are implicated in inflammatory responses;37–39 non-canonical Wnt actions are prototypically inflammatory, whereas canonical Wnt actions are anti-inflammatory; however, there are exceptions.34,40 Un-checked or overwhelming non-canonical Wnt signalling may perpetuate chronic low-grade inflammation and contribute to CMD. Wnts are important regulators of proliferation, migration and differentiation; these processes require tightly controlled changes in energy supply and demand. Thus, dysregulation of Wnt signalling likely influences metabolism.
Canonical and non-canonical Wnt ligands regulate key metabolic signalling pathways such as mTOR and insulin signalling.30,41–43 For example, canonical signalling can downregulate mTOR signalling, a key activator of aerobic glycolysis. 43 On the other hand, non-canonical signalling has been shown to have an opposing effect and activate mTOR.43,44 Canonical Wnt signalling has also been shown to regulate a number of metabolic enzymes such as aldolase and cytidine deaminase in carbohydrate and amino acid metabolism, respectively, and dihydrolipoamide S-succinyltransferase, in the tricarboxylic acid cycle. 42 Additionally, identification of putative TCF/LEF binding sites has yielded at least eight additional metabolic enzymes regulated by canonical signalling.45,46 Wnt signalling appears to be a metabolic rheostat that, when chronically dysregulated, may contribute to the pathogenesis of CMDs. An in-depth review of Wnt signalling in the control of cellular metabolism (carbohydrate, protein, lipid) has been reviewed in great detail by Sethi and Vidal-Puig. 46 Importantly for this review, the Wnt signalling pathways have been strongly implicated in metabolic dysfunction in adipose tissue,30,47 vasculature13,48–50 and co-morbidities associated with metabolic syndrome. So, the implication of Wnt signalling in homeostatic metabolism of carbohydrates, proteins and lipids, as well as metabolic dysfunction, supports a role of canonical and non-canonical signalling in metabolism. However, more work is necessary to fully understand Wnt signalling in these contexts.
Wnt signalling in CMDs
Obesity and diabetes
Adipogenesis
Approximately two-thirds of adult Americans are considered to be obese, and obesity is a risk factor for diabetes, fatty liver disease, CVD and cancer. 51 Wnt signalling has been implicated as a key regulator of adipose tissue development with canonical Wnt signalling generally regarded as anti-adipogenic and non-canonical Wnt signalling as promoting adipogenesis and lipid accumulation. 52 Non-canonical Wnts, Wnt5a and Wnt5b, have been shown to promote adipogenesis by increasing PPAR-γ expression which is critical for adipose tissue development. 53 Furthermore, Wnt5a and Wnt5b have been demonstrated to activate Hippo-YAP/TAZ signalling to inhibit canonical Wnt signalling and promote adipogenesis in 3T3 cells. 22 Non-canonical Wnt5a may work indirectly, and/or in concert with Wnt5b, to promote adipogenesis and lipid accumulation in adipocytes.22,53 The non-canonical Wnt receptor, Ror1, has also been suggested to play an important role in adipogenesis, 54 further supporting the role of non-canonical Wnt signalling in obesity.
In contrast to the effects of non-canonical signalling, induction of canonical signalling has been shown to prevent adipogenesis and blockade of canonical signalling at multiple levels of the signalling cascade promotes adipogenesis.11,55 Importantly, canonical Wnt signalling is an important negative regulator of PPAR-γ, thus retarding adipogenesis. 52 Lu et al. 56 further support the role of canonical signalling as anti-adipogenic by demonstrating endogenous antagonism of canonical Wnt signalling by dickkopf-1 (DKK-1) promotes differentiation and adipokine secretion in pre-adipocytes. Furthermore, DKK-1 exposure is able to accelerate the differentiation of pre-adipocytes by downregulating canonical Wnt signalling independent of activated non-canonical Wnt signalling. 56
There are some reports of non-canonical Wnt5a having differential effects on adipogenesis under in vitro and in vivo conditions.57,58 These discrepancies may be due to differences in the developmental processes of adipocytes when comparing human/mouse mesenchymal stem cells and committed pre-adipocytes.57,58 However, majority of reports in the literature support the cannon that canonical Wnt signalling functions as anti-adipogenic and non-canonical Wnt signalling functioning as pro-adipogenic and promoting fat development.
Diabetes
Non-canonical Wnt signalling has been shown to negatively regulate insulin signalling in white adipose tissue. 30 Adipose tissue insulin resistance is a hallmark of type 2 diabetes. 59 Mice and humans with obesity-induced insulin resistance have higher levels of circulating non-canonical Wnt5a compared to healthy, lean controls. 30 The non-canonical receptor, Ror1, has also been suggested to play an important role in glucose homeostasis. 54 Furthermore, Wnt5a, and Jun N-terminal kinase (JNK), are elevated in diabetic vascular endothelial cells, implicating the non-canonical signalling pathway in diabetes.13,30 Thus, blocking non-canonical signalling may be a useful target to restore insulin sensitivity in type 2 diabetes. 30
The canonical Wnt signalling pathway is also implicated in diabetes and indirect promotion of the canonical pathway may be beneficial. For example, inhibition of the canonical Wnt antagonist, DKK-2, improves glucose tolerance in type 2 diabetic mice. 60 Improvement in glucose tolerance has also been demonstrated using the Wnt5a sequester, sFRP5. 33 Antagonism of non-canonical signalling in adipose tissue with sFRP5 has been demonstrated to be a strong anti-inflammatory adipokine.31,33 sFRP5-deficient mice on a high-calorie diet developed signs of metabolic dysfunction (glucose intolerance and hepatic steatosis) but were able to be rescued through adenoviral delivery of sFRP5. 33 As mentioned previously, Wnt proteins are detectable in serum and levels correlate with diabetes and CVD. Interestingly, Farb et al. 47 have tackled the complex condition of the metabolically healthy obese patient. The authors demonstrated the quality (inflamed or non-inflamed) of adipose tissue may, in part, explain why some obese patients are metabolically healthy. 47 sFRP5 has been described as an anti-inflammatory adipokine and supports that antagonism of the non-canonical signalling pathway may be a way to promote a metabolically healthy phenotype. Specifically, Ouchi et al. 33 have suggested that the sFRP5 and the Wnt signalling pathway may be a prime therapeutic target to combat obesity and obesity-induced insulin resistance. Serum levels of other Wnts and Wnt agonists/antagonists in the context of metabolically healthy/unhealthy obesity are not known and warrant further investigation.
Overall, activated canonical signalling inhibits adipocyte differentiation and lipid accumulation; in contrast, non-canonical signalling promotes lipid accumulation and insulin resistance in white adipose tissue. Additionally, activated non-canonical signalling contributes to the pathogenesis of diabetes while canonical signalling appearing to be protective. Imbalanced canonical and non-canonical Wnt signalling appears to be an important homeostatic mechanism regulating not only accumulation of lipids in adipocytes but also insulin resistance in diabetes.
NAFLD
NAFLD progresses to non-alcoholic steatohepatitis (NASH) through a largely unknown mechanism. Serum levels of non-canonical Wnt5a correlate with hepatic steatosis, NAFLD and NASH and it is known that activation of JNK signalling and subsequent insulin resistance contributes to hepatic steatosis. 61 As Wnt5a activates JNK, it is reasonable that Wnt5a signalling may contribute to hepatic steatosis, NAFLD and NASH. In fact, sFRP5 (endogenous Wnt5a quencher) is able to reduce Wnt5a-mediated JNK activation in mice resulting in reduced levels of triglyceride within the liver. 33 Roux-en-Y gastric bypass (RYGB) is a procedure to treat hepatic steatosis, NAFLD and NASH associated with obesity. 62 Interestingly, patients who underwent RYGB for weight loss saw expected improvements in measures of obesity, but also a significant reduction in serum Wnt5a. 15 Importantly, there was no observable change in sFRP5 levels in these patients. 15 This suggests that Wnt5a could be a potential biomarker post-RYGB 15 or perhaps indicator of a patient’s adherence to lifestyle modification.
Non-canonical signalling is generally regarded as pro-inflammatory and likely contributes to the inflammatory component of NASH. Tian et al. 63 suggested that inhibition of cyclooxygenase-2 (COX-2) with celecoxib could reduce NASH via suppression of Wnt5a signalling components. A decrease in Wnt5a signalling components in the liver in response to an anti-inflammatory drug supports Wnt5a’s importance in prolonged inflammation.
Inactivating mutations of LRP6, a key co-receptor in canonical Wnt signalling, in mice is sufficient to induce steatohepatitis, fibrosis and accumulation of lipid by 9 months of age, 10 suggesting that reduction of canonical signalling is sufficient to induce lipid accumulation in the liver. Interestingly, activation of canonical Wnt signalling through recombinant Wnt3a administration is able to prevent the inflammation and lipid accumulation associated with disease progression from NAFLD to NASH.10,64 In agreement with this, knocking out β-catenin in the liver mimics inactivation of canonical Wnt signalling and perpetuates a liver pathology similar to NASH. 65 Behari et al. 65 also evaluated the lipid profile of steatohepatitis in β-catenin knock-out mice and found increased cholesterol and triglyceride accumulation as well as an overall increase in steatosis, inflammation and hepatocyte ballooning. Thus, activating canonical Wnt signalling may be protective in NAFLD and NASH. Interestingly, a family of agonists for the canonical Wnt pathway, roof plate-specific spondins (RSPOs), increase hepatic stellate cell activation and induce liver fibrosis in mice through canonical signalling activation. 66 RSPOs were found to be elevated in human fibrotic liver and an inhibitor of the canonical pathway, DKK1, is able to prevent RSPO-Wnt induced liver fibrosis. 66 On the other hand, Wnt5a has been suggested to have profibrotic activities and is under the control of TGFβ in liver fibroblasts. 67 These contrasting results add a level of complexity and suggest that an intricate balancing of Wnt signalling may be necessary when considering its usefulness as a therapeutic target.
In summary, the ability of Wnt5a-JNK signalling to interfere with insulin signalling in the liver, the correlation of serum Wnt5a with NAFLD/NASH and the ability of anti-inflammatory drugs to reduce non-canonical signalling suggest that non-canonical signalling contributes to hepatic steatosis, NAFLD and NASH. The ability of canonical signalling to rescue from NAFLD, but if over activated can also induce liver fibrosis, makes manipulation of the Wnt signalling pathways particularly complex.
CKD
CKD is defined as progressive loss of kidney function over time, most commonly because of diabetes, hypertension and other chronic diseases. CKD is a major global public health concern resulting in 956,000 deaths in 2013 and presents treatment challenges in societal interventions and medical practice.7,68 Wnt signalling is implicated in the embryological development of the kidney, but the role of Wnt signalling in adult kidney pathology is currently debated. 4 Some reports suggest no involvement of any Wnt signalling, while others suggest canonical Wnt signalling contributes to adult kidney fibrosis.69–71 Still others suggest canonical Wnt signalling contributes to end-stage renal disease by promoting podocyte dysfunction. 72
Unilateral ureteral obstruction (UUO) mouse models of obstructive nephropathy are most commonly used to investigate mechanisms of CKD.73,74 Reports of elevated Wnt signalling in obstructive nephropathy are evident.70,75 Furthermore, blocking of canonical Wnt signalling with DKK1 decreases expression of markers of fibrosis (e.g. alpha-smooth muscle actin, collagen type 1 and fibroblast specific protein-1). 70 Inhibition of canonical signalling also reduced total renal fibrosis and matrix deposition, 70 suggesting that aberrant canonical Wnt signalling contributes to renal fibrosis. In another study, inhibition of canonical signalling was able to prevent fibrosis, inhibit inflammation and prevent macrophage infiltration in mouse kidneys. 8 Furthermore, exogenous administration of sFRP4 was able to rescue mice from the fibrotic changes associated with elevated canonical Wnt signalling in the kidney. 75 Thus, elevated canonical Wnt signalling contributes to kidney fibrosis and therapeutic inhibition of canonical Wnt signalling may be an effective way to combat CKD.
The role of non-canonical signalling in the context of CKD is not well understood relative to canonical signalling; however, the integrated relationships of these pathways suggest that normalizing both Wnt signalling pathways may be an attractive direction for future research and therapeutic development. Specifically, excessive canonical signalling appears to promote the fibrosis and decline in kidney function in CKD and increasing non-canonical signalling to inhibit canonical signalling may be protective in this context.
Activation of the renin-angiotensin system (RAS) is a key component of hypertension, CVD and CKD. However, effective pharmacological control of RAS has been difficult given no overall reduction in CVD risk; this is likely due to complex compensatory regulation of RAS. 76 Using bioinformatics techniques, canonical Wnt signalling transcription factor binding sites for TCF/LEF were found to be present in the promoter regions of RAS-specific genes, 8 suggesting that canonical signalling can directly regulate RAS. Additionally, a small-molecule inhibitor of β-catenin was able to prevent RAS induction in mice. 8 The novel finding that Wnt/β-catenin signalling can regulate the RAS axis may be particularly relevant for the treatment of CVD and normalizing Wnt signalling may provide multiple beneficial effects in these diseases. 77 Thus, effective treatment likely should focus on multiple RAS targets, including Wnt signalling, for optimum therapy and outcomes.
CVD
CVD is highly prevalent in developed countries such as the United States where 86 million people have at least one type of CVD. 78 Atherosclerosis is the root cause of heart attack and stroke, which together comprise nearly 18% of those with at least one CVD. 78 The earliest recognizable lesion is the fatty streak, which is ubiquitously present in the vasculature, evidenced by autopsies showing fatty streaks in the aorta and coronary arteries of 10- to 14-year-old children. 79 Non-canonical Wnt5a was the first Wnt associated with atherosclerosis. 80 Additionally, Wnt5a RNA correlates with atherosclerotic plaque severity and serum levels of Wnt5a are elevated in atherosclerotic patients. 16 The roles of canonical and non-canonical Wnt signalling have gained considerable attention in many cell types associated with atherosclerosis, in particular endothelial cells, macrophages and vascular smooth muscle cells (VSMCs).50,81 For this reason, Wnts have been intensely investigated in recent years as playing an important role in the pathogenesis of CVD.
Endothelial cells
Progression of atherosclerosis begins with an initial endothelial injury and subsequent endothelial dysfunction. 82 Wnt5a has been shown to reduce insulin-dependent eNOS activity, resulting in impaired vasodilation, and illustrates Wnt5a’s role in diabetic endothelial dysfunction, which may help explain the increased incidence of atherosclerosis in diabetic patients.13,48 Importantly, Wnt5a-induced endothelial dysfunction is independent of reactive oxygen species production. 13 Wnt signalling has been shown to regulate proteins involved in cell-to-cell junctions (e.g. adherens, cadherins and catenins) and, as such, may play a role in atherogenesis and progression of the disease. In the context of atherosclerosis, vascular cell adhesion molecule 1 (VCAM-1) is critical for recruitment of inflammatory cells to the atheroma. 83 Importantly, non-canonical Wnt5a signalling has been demonstrated to upregulate intercellular adhesion molecule 1 (ICAM-1) and VCAM-1 expression in haematopoietic progenitors, 84 supporting the role of non-canonical Wnt signalling in the initiation of the inflammatory response and atheroma development.
Canonical Wnt3a signalling has been shown to reduce VCAM-1 expression and can limit export of myeloid cells from the bone marrow. 85 Inhibition of GSK3β was also able to prevent nuclear factor kappa-light-chain-enhancer of activated B cells (NFkB) activation. 86 VCAM-1 is important for leukocyte transmigration and it is reasonable to suspect that Wnt signalling also plays a role in macrophage recruitment to diseased vasculature.
Macrophages
Non-canonical Wnt5a is an inflammatory stimulus that has been shown to regulate macrophage phenotypic shifts in the atheroma.87–90 Critical to plaque development is fatty streak formation and accumulation of lipid, forming the plaque core. Our group has recently demonstrated non-canonical Wnt5a signalling components correlate with plaque severity and, furthermore, lead to increased lipid accumulation in macrophages and foam cell formation (unpublished). Furthermore, silencing Wnt5a expression results in inhibition of progression of atherosclerosis and a reduction in the size and severity of plaques without affecting blood lipid levels. 91 Given Wnt5a’s lipid-specific actions are independent of systemic lipid levels is important because the goal of current therapies are to normalize lipid levels, yet CVD continues to be the number one killer in the United States. 78 Targeting non-canonical Wnt signalling, in addition to systemic lipids, may provide a novel way to combat atherosclerosis.
VSMCs
Proliferation, migration and differentiation of smooth muscle cells into a myofibroblast phenotype are important for plaque stability.82,92 Canonical and non-canonical Wnt signalling have been demonstrated to dictate adipogenic and myofibrotic phenotypic switches in pericytes and VSMCs.91,93 Thus, Wnt signalling may regulate plaque stability and severity in the context of fibrous cap formation by VSMCs. Specifically, canonical Wnts (Wnt2 and Wnt4) have been demonstrated to promote VSMC proliferation and migration.94–96 Increased VSMC proliferation and migration results in intimal thickening, but also may be athero-protective by increasing fibrous cap formation by myofibroblasts resulting in a more stable plaque. Inhibition of canonical Wnt signalling may be particularly beneficial in preventing restenosis and vein graft failure.95,96 However, the precise role of canonical versus non-canonical signalling in the context of fibrous cap/plaque stability is still unclear.97–99
Recently, Pandey and Chandravati 100 have suggested that Wnt signalling can complement public health initiatives in predicting occlusive vascular disease (restenosis after coronary artery bypass graft surgery) which is a major public health problem in the United States. Identification of new molecular mechanisms in coordination with clinical data is crucial in the development of cost-effective biomarkers for this at-risk population. 100 Wnt signalling contributes to multiple levels of atherosclerosis pathogenesis: initial endothelial dysfunction, plaque instability and size of the lipid core; however, the full contribution of non-canonical and canonical Wnts in the development of atherosclerosis and CVD has yet to be elucidated in detail. Thus, therapeutics targeting Wnt signalling may provide a novel way to combat atherosclerosis.
Conclusion
In conclusion, Wnts have broad implications in obesity, diabetes, NAFLD, CKD and atherosclerosis (CVD) through promoting lipid accumulation, inflammation and fibrosis. These features make the Wnt signalling pathways an attractive therapeutic target. Although evidences presented in this review demonstrate some functions of Wnt signalling pathways in CMDs through homeostatic and inflammatory mechanisms (Figure 2), the precise role of Wnts in a patient with multiple disease co-morbidities remains unknown.
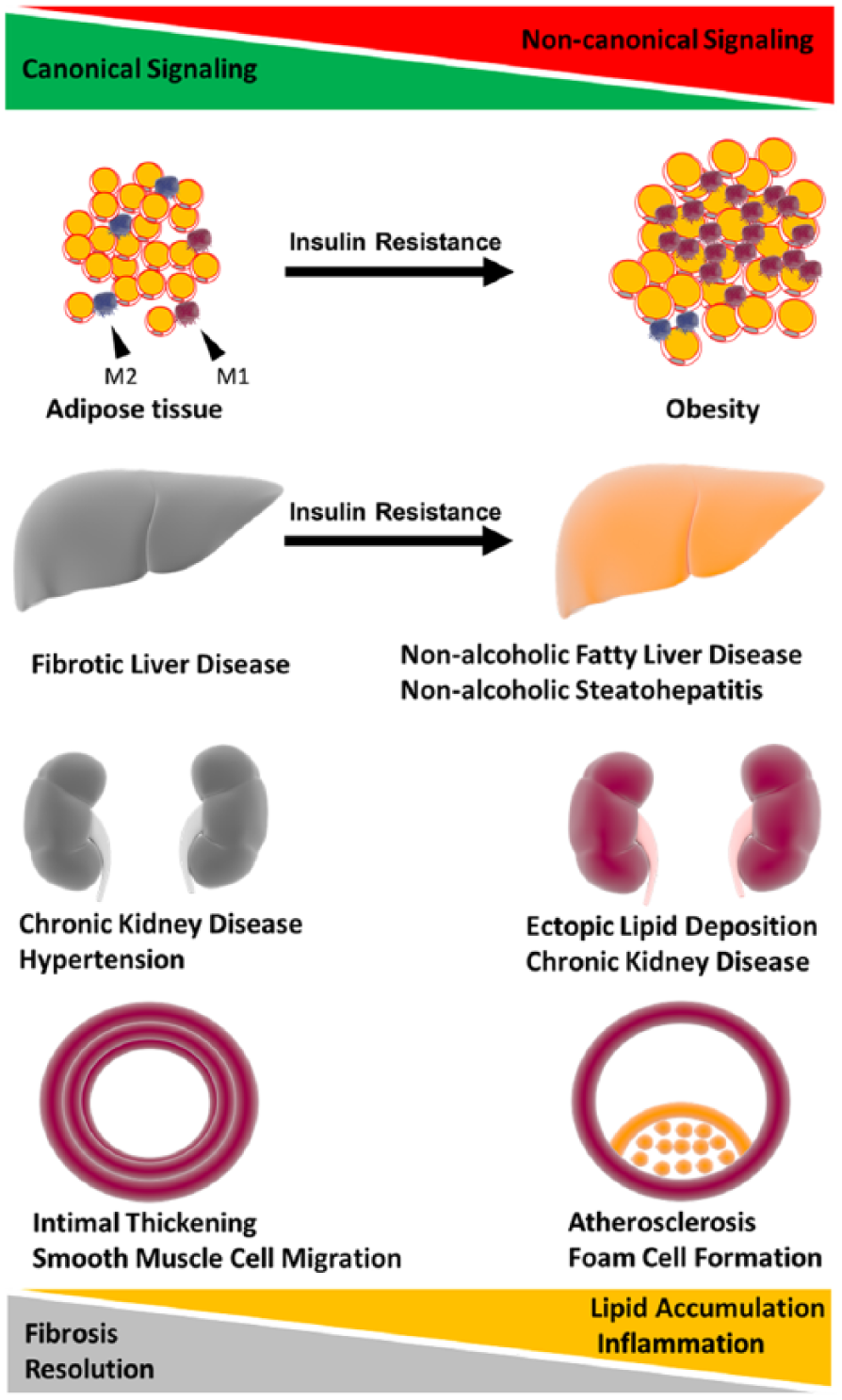
Overarching hypothesis. Chronic metabolic disease involving fibrosis and extracellular matrix deposition have dysregulated Wnt signalling and a tendency to have excessive activation of the canonical Wnt signalling pathway. Chronic metabolic diseases involving inflammation and lipid accumulation have dysregulated Wnt signalling and a tendency towards excessive activation of the non-canonical Wnt signalling pathway. Excessive non-canonical signalling has also been shown to drive insulin resistance in adipose tissue and liver. The role of canonical and non-canonical Wnt signalling in kidney disease is less clear. However, it appears to be contingent on over/under activation of either pathway both contributing to chronic kidney disease. Wnts are implicated in obesity, type II diabetes (insulin resistance), NAFLD, CKD and cardiovascular disease through lipid accumulation and fibrosis. The intricate balance of both arms of the Wnt signalling pathways with fibrosis, inflammation and lipid accumulation may contribute to the development of these chronic metabolic diseases.
The pervasiveness of aberrant Wnt signalling in CMDs makes it an interesting target as a therapeutic as well as a potential biomarker. Targeting the Wnt signalling pathways may be particularly useful in identification of the metabolically healthy/unhealthy obese. The unique aspects of canonical and non-canonical Wnt signalling are additional challenges when considering the development of therapeutic strategies such as avoiding potential off target effects of Wnt signalling modulation.
Discrepancies in the literature concerning overall effects of canonical and non-canonical signalling appear to depend on the affected tissue, reiterating the importance of avoiding off target effects. Additionally, there are mixed reports of the role of Wnt signalling depending on in vitro and in vivo conditions and echoes that the Wnt signalling pathways are highly cell and context dependent. A concept to highlight is the complexity of Wnt signalling at the systemic, receptor and intracellular levels. Also, the actions of Wnts are implicated in multiple organ systems. The Wnt signalling pathway may also play distinct roles at various stages of each disease over time. These additional levels of complexity represent additional barriers when developing a therapeutic intervention. In conclusion, the literature supports an interrelationship between the Wnt signalling pathways as a potential pathological mechanism in CMDs, and a greater understanding of the complexity of Wnt signalling will increase the opportunity to develop effective treatments for these burdensome diseases.
Wnt signalling is implicated in lipid accumulation, fibrosis and chronic low-grade inflammation in lifestyle-related chronic metabolic diseases.
Targeting the Wnt signalling pathways may be novel approach to treating lifestyle-related chronic metabolic diseases.
Given complexity of the Wnt signalling pathways and their role in multiple organ systems, it is important to avoid potential off target effects of Wnt signalling inhibition.
Footnotes
Acknowledgements
The authors would like to thank Dr Karen Coschigano and Dr Kelly McCall for critical review of the manuscript. Stipend and Fellowship support to Ian Ackers generously provided by the Osteopathic Heritage Foundations, Dual Degree Program, Ohio University Heritage College of Osteopathic Medicine, Athens, Ohio. The authors would also like to thank the Kopchick MCB/TBS Research Fellowship awarded to Ian Ackers. Clip art for figures: Library of Science and Medical Illustrations. Creative Commons license (CC BY-NC-SA 4.0). ![]() .
.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.