
Abstract
Background:
Sodium glucose transporter type 2 inhibitors may reduce cardiovascular events in type 2 diabetes. Our study aimed to determine the effect of the sodium glucose transporter type 2 inhibitor dapagliflozin on endothelial cell activation, vasoreactivity and atherogenesis using in vitro and in vivo models and identify associated molecular mechanisms.
Methods:
In vitro studies utilised human vascular endothelial cells stimulated with tumour necrosis factor α or hyperglycaemic conditions. In vivo studies were performed in C57Bl/6J mice to evaluate direct vasorelaxation responses evoked by acute dapagliflozin administration and acute vaso-protective effects of dapagliflozin on hyperglycaemia-induced endothelial dysfunction. Adult and aged Apolipoprotein E–deficient mice maintained on a high-fat diet were used to investigate endothelial-dependent vascular reactivity and atherogenesis. Dapagliflozin treatment (1.0 mg/kg/day) was administered for 4 weeks.
Results:
In vitro studies demonstrated dapagliflozin-mediated attenuation of tumour necrosis factor α- and hyperglycaemia-induced increases in intercellular adhesion molecule-1, vascular cell adhesion molecule-1, plasminogen activator inhibitor type 1 and NFκB expression. Acute dapagliflozin administration dose-dependently induced endothelium-independent vasorelaxation. Chronic dapagliflozin treatment improved endothelial function and significantly reduced in vivo vascular adhesion molecule and phospho-IκB expression together with macrophage vessel wall infiltration.
Conclusion:
These observations identify a potential role for dapagliflozin in the attenuation of atherogenesis and identify anti-inflammatory molecular mechanisms associated with these effects.
Keywords
Introduction
The sodium glucose transporter type 2 (SGLT-2) inhibitors represent a novel class of anti-diabetic agents used for the treatment of type 2 diabetes. 1 SGLT-2 inhibitors limit renal glucose reabsorption promoting urinary excretion of glucose, thereby reducing plasma glucose. 1 Cardiovascular safety studies suggest a potential benefit from treatment with the SGLT-2 inhibitor empagliflozin. 2 Dapagliflozin (DAPA) was one of the first SGLT-2 inhibitors to be made available worldwide. 3 Pre-clinical studies suggest potential cardiovascular benefits from DAPA treatment,4,5 while preliminary clinical studies have indicated no significant cardiovascular risk with DAPA treatment. 3 The long-term cardiovascular safety of DAPA is currently being evaluated in an international multicenter trial, DECLARE-TIMI 58. 6
Identification of non-fluid-mediated weight loss, reductions in systolic blood pressure together with reductions in high-sensitivity C-reactive protein (hsCRP) and modest improvements in serum lipid ratio profiles in DAPA-treated patients 7 suggest potential cardiovascular benefits of this agent via either direct SGLT-2 inhibition or novelmolecular mechanisms.
Pre-clinical investigation into the effects and associated molecular mechanisms of action of DAPA in models of cardiovascular disease represents a novel and potentially fertile field of research with regard to expanding the clinical benefits ascribed to this class of agents beyond glycaemic control and potentially impacting on the accelerated cardiovascular disease burden identified in the type II diabetic patient population.
We aimed to (a) characterise the in vitro effects of DAPA on tumour necrosis factor α (TNFα) and hyperglycaemia-mediated induction of vascular peptide expression in human vascular endothelial cells and (b) evaluate the ex vivo and in vivo effects of acute and chronic DAPA administration on vascular reactivity and early markers of atherogenesis.
Materials and methods
In vitro studies
In vitro studies utilised TNFα-stimulated (10 ng/mL, 24 h) and hyperglycaemia-stimulated (10–30 mM, 24 h) human umbilical vein endothelial cells (HUVECs) [Lonza (CC-2519, pooled donor), Basel, Switzerland] maintained in Media-199 (Sigma, USA) supplemented with penicillin/streptomycin, 20% foetal calf serum (FCS), 20 µg/mL endothelial cell growth factor (Sigma) and 20 µg/mL heparin. Soluble intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and plasminogen activator inhibitor type 1 (PAI-1) protein levels were determined from conditioned medium of TNFα- or hyperglycaemia-stimulated, DAPA-treated (24 h, 1.0 nM–10.0 µM) HUVEC using enzyme-linked immunosorbent assay (ELISA; elisakit.com, Melbourne, Australia). DAPA was provided by AstraZeneca (AB, Sweden) in the non-hydrated form with a molecular weight (MW) of 408.9 Da. A 100-mM stock solution of DAPA was prepared using DAPA dissolved in water as advised. In vitro DAPA doses were determined based on the previous characterisation of EC50 of 1.0 nM for DAPA. 8
Real-time quantitative polymerase chain reaction
Total RNA was extracted from 24 h TNFα-stimulated, DAPA-treated (24 h, 1.0 nM–10.0 µM) HUVEC using Trizol reagent (Invitrogen, Walthan, MA, USA). An aliquot (2 µL) of total RNA was reverse transcribed in 20 µL reaction mixture at 50°C for 1 h. Complementary DNA (cDNA) was amplified by real-time polymerase chain reaction (PCR) using specific primers: PAI-1: forward primer 5′-CAG-ACC-AAG-AGC-CTC-TCC-AC-3′, reverse primer 5′-ATC-ACT-TGG-CCC-ATG-AAA-AG-3′; NFκB: forward primer 5′-CAG-GAA-GAT-GTG-GTG-GAG-GAT-3′, reverse primer 5′-TGT-CGT-GCT-CCA-CAC-AGC-CAG-GT-3′ and human β actin: forward primer 5′-CTG-GAC-TTC-GAG-CAA-GAG-AT-3′ and reverse primer 5′-GAT-GTC-CAC-GTC-ACA-CTT-CA-3′.
Each PCR run also included wells of no template control (NTC). A melting point dissociation curve generated by the instrument was used to confirm that only a single product was present. The fluorescence data were quantitated using the threshold cycle (CT) value. Data were normalised to β-actin and presented as the mean fold change compared with untreated samples
In vivo studies
Male C57BL/6J mice and B6 Apolipoprotein E–deficient (ApoE−/−) mice with a >99% C57BL/6J background weighing approximately 30 g were obtained from Monash Animal Research Laboratories (ARL). Mice were housed in the Pharmacology Animal House, Monash University, in standard mouse cages (with approximately four mice per cage) at 21°C ± 1°C–5°C, with a 12-h light–dark cycle. All mice had access to food and water ad libitum. All treatments and experimental procedures were approved by the School of Biomedical Sciences (SOBS) Animal Ethics Committee, Monash University (ethics no. SOBSB/PHAR/2013/118).
Vascular reactivity model
Aortic rings taken from 5-month-old C57Bl/6J mice maintained on a normal chow diet were utilised to evaluate direct vasorelaxation responses evoked by acute DAPA administration and acute vaso-protective effects of DAPA on hyperglycaemia-induced endothelial dysfunction in organ bath studies.
Atherogenesis models
Male ApoE−/− mice (7 months old) were maintained on either a normal fat or a high-fat diet (HFD; 22% fat and 0.15% cholesterol (Specialty Feeds, Glen Forrest, Perth, Western Australia) for 16 weeks with mice from each diet group randomly allocated to receive either DAPA (1.0 mg/kg/day, via drinking water) or vehicle (saline, via drinking water) for the final 4 weeks. These animals represent an aged cohort (11–12 months). Doses of DAPA used in the study were based on those utilised in previous pre-clinical models4,5
Male ApoE−/− mice (6 weeks old) were placed on a HFD (22% fat and 0.15% cholesterol; Specialty Feeds) for 16 weeks with mice being randomly allocated to receive either DAPA (1.0 mg/kg/day, via gavage) or vehicle (saline, via gavage) for the final 4 weeks. These animals represent an adult cohort (5–6-months old).
Biometric assessment
Baseline and end-of-treatment (4 weeks) body weight was measured in grams. Glucose tolerance test was performed. Blood glucose levels of DAPA or vehicle-treated ApoE−/− mice were measured after the animals were fasted for 8 h. Blood glucose was measured using a blood glucose monitoring system (Accu-Chek) prior to a glucose challenge load (2 g/kg i.p.) being administered. Blood glucose levels were then measured at 15, 30, 60 and 120 min after a glucose loading dose.
Aortic dissection
The whole heart and aorta were removed and placed in ice-cold Krebs-bicarbonate buffer (pH 7.4) comprising (in mM) NaCl 118, KCl 4.7, KH2·PO4 1.2, MgSO4·7H2O 1.2, CaCl2 2.5, NaHCO3 25 and glucose 11.7. Fat and connective tissue deposits were removed from both the heart and aorta. The aorta was dissected into abdominal and thoracic sections. Three aortic rings were taken from the abdominal aorta for vascular reactivity assessment using isolated organ baths, while the remaining abdominal aorta was embedded in optimal cutting temperature (OCT compound) and the thoracic aorta was snap frozen in liquid N2 and stored at −80°C.
Assessment of direct vascular reactivity of DAPA
Initial studies examined the direct effect of DAPA on pre-constricted aortic rings from C57Bl/6J mice. Three aortic rings (~3 mm/ring) from each mouse were suspended between two 100-µm stainless steel wires connected to an isometric force transducer (FT-03; Grass Instruments, Astro-Med Inc, R.I, USA). Each mouse had one aortic ring randomly allocated as a time control, one ring with intact endothelium and one ring with endothelium mechanically removed by gently rolling the lumen of the vessel on a thin wire. Aortic rings were immersed in organ baths containing Krebs solution as previously described 9 and were maintained at 37°C with bubbled carbogen (95% O2, 5% CO2). Tissues were gradually stretched to their optimum tension of 0.5 g over a 60-min period. After 15 min equilibration at 0.5 g a maximum contraction was provoked using U46619 (1.0 µM). When the response plateaued, tissues were washed with Krebs solution a number of times until the tissue returned to baseline tension. Any tissues that did not constrict to at least 0.5 g were excluded. All viable tissues were then pre-contracted to 70%–80% maximal contraction of U44619. After the response plateaued, 10 µM ACh was added to determine extent of endothelial cell integrity, with relaxation <30% indicating endothelial denudation and relaxation >70% indicating intact endothelium. Following this, tissues were washed, allowed to recover before being pre-constricted again to 70%–80% of the U46619 maximal contraction. A DAPA concentration response curve (CRC) was performed (1 × 10−9 M to 1 × 10−4 M) to assess direct effects of DAPA to relax the aorta and the dependence on an intact endothelium. Finally, 1 × 10−5 M of sodium nitroprusside (SNP) was added to each organ bath to determine the integrity of the vascular smooth muscle of each preparation. Investigators were blinded to the aortic ring treatment groups for vascular reactivity studies.
Assessment of acute vaso-protective effect of DAPA
Three aortic rings (~3 mm/ring) from each C57Bl/6J mouse were suspended between two 100-µm stainless steel wires connected to an isometric force transducer (FT-03; Grass Instruments). Aortic rings were immersed in organ baths containing Krebs solution as outlined above and were gradually stretched to their optimum tension of 0.5 g over a 60-min period. After 15 min of equilibration at 0.5 g, a maximum contraction was provoked using U46619 (1.0 µM). When the response plateaued, tissues were washed with Krebs solution a number of times until the tissue returned to baseline tension. To test endothelial integrity, all tissues were pre-contracted to 70%–80% maximal contraction of U44619 and then relaxed with 10 µM ACh. Tissues were considered viable if they constricted by at least 0.5 g to the maximal concentration of U46619 and if they relaxed by ⩾60%. Viable tissues were then randomly allocated to be bathed in normal glucose (11 mM) Krebs solution, high-glucose (HG-44 mM) Krebs solution or HG Krebs solution plus DAPA (1.0 µM) for 4 h to induce hyperglycaemia-mediated endothelial dysfunction. At the end of 4-h incubation, tissues were again submaximally pre-constricted with U46619, and an ACh CRC was performed (1 × 10−9 M to 1 × 10−4 M) to assess acute effects of DAPA to protect the aorta from hyperglycaemia-induced endothelial dysfunction. At the conclusion of ACh CRC, 10.0 µM SNP was added to each organ bath to determine the integrity of the vascular smooth muscle of each preparation. Investigators were blinded to the aortic ring treatment groups for vascular reactivity studies.
Assessment of chronic vaso-protective effect of DAPA
Using aged and adult atherogenesis models, the ability of chronic DAPA treatment to improve endothelial function was examined. Using a previously described protocol, 10 we assessed ACh-induced endothelium-dependent vasorelaxation in aortic rings from ApoE−/− mice chronically treated with either DAPA or vehicle.
Assessment of atherosclerotic lesion formation
Two methods for analysis of lesion development were performed. Using en face staining, the aortic arch and thoracic aortae were dissected free of connective tissue, and lipid deposition was visualised with Sudan IV staining. Images were captured using a Moticam-480P Digital Camera and quantified using the appropriate software (Motic Images Plus 3.0, Kowloon, HK). Lesion area was expressed as a percentage of the total luminal area of the vessel. In addition, cross-sectional analysis of luminal encroachment in haematoxylin-and-eosin-stained sections, expressed as an intimal:medial ratio (IMR), was performed at the aortic root as previously described. 11
Immunohistochemical localisation of inflammatory markers
Immunostaining was performed on frozen 10-µM-thick aortic cross sections taken from the area immediately adjacent to aortic segments used for vascular reactivity studies, the area known as the thoracoabdominal aorta, as previously described. 9 Sections were air dried and fixed in ice-cold acetone for approximately 15 min before washing with 0.01 M phosphate-buffered saline (PBS) buffer (3 × 10 min). Sections were then incubated with 10% goat serum in 0.01 M PBS for 30 min to reduce non-specific binding. Next, blocking buffers were removed, and the primary antibody to respective markers was applied overnight at room temperature based on the following dilution and origin of the antibodies: ICAM-1 rabbit polyclonal antibody (1:500; Abcam, Cambridge, UK), F4/80 macrophage rat monoclonal antibody (1:100; Serotec, CA, USA) and P-IκBα rabbit monoclonal antibody (1:200; Cell Signalling, MA, USA); appropriate negative controls were undertaken using antibody only diluent (Dako, CA, USA).Following this, 3 × 10-min washes in ice-cold PBS were performed on the second day. The sections were then incubated with the appropriate AlexaFluor 488 secondary antibody (Invitrogen). All immunofluorescent sections were viewed under ×20 magnification on an Olympus BX51 microscope (Tokyo, Japan), and images were analysed using ImageJ (JAVA, NIH).
Statistical analysis
All results are expressed as mean ± standard error of mean (s.e.m.). All statistical plots and analysis were performed using the Prism program (GraphPad Prism 6 Software, Inc., San Diego, CA, USA). Statistical comparisons of dose–response curves were performed using two-way repeated measures analysis of variance (ANOVA) with Bonferroni post hoc test to correct for multiple comparisons. All other data were compared using either one-way ANOVA with Tukey post hoc test for multiple comparisons or unpaired t-tests. p < 0.05 was deemed statistically significant.
Results
DAPA attenuates TNFα- and hyperglycaemia-induced human vascular endothelial cell activation in vitro
Significant induction of ICAM-1 and VCAM-1 protein levels was identified after 24 h of TNFα stimulation compared with unstimulated HUVECs (Figure 1). Addition of DAPA (1.0–5.0 nM) to TNFα-stimulated HUVECs for 24 h resulted in a significant attenuation of TNFα-mediated induction of ICAM-1 and VCAM-1 protein expression (Figure 1). Significant induction of ICAM-1 protein levels was identified after 24 h of hyperglycaemia (20–30 mM) stimulation compared with unstimulated HUVECs (Figure 2). Addition of DAPA (1.0–5.0 nM) to hyperglycaemia-stimulated HUVECs for 24 h resulted in a significant attenuation of hyperglycaemia-mediated induction of ICAM-1 expression (Figure 2(a)). No significant effect of high-dose DAPA (100 nM and 1.0 µM) was identified on hyperglycaemia-induced ICAM-1 expression in HUVECs (Figure 2(b)). No effect of DAPA alone on ICAM-1 or VCAM-1 protein expression was identified in HUVECs.
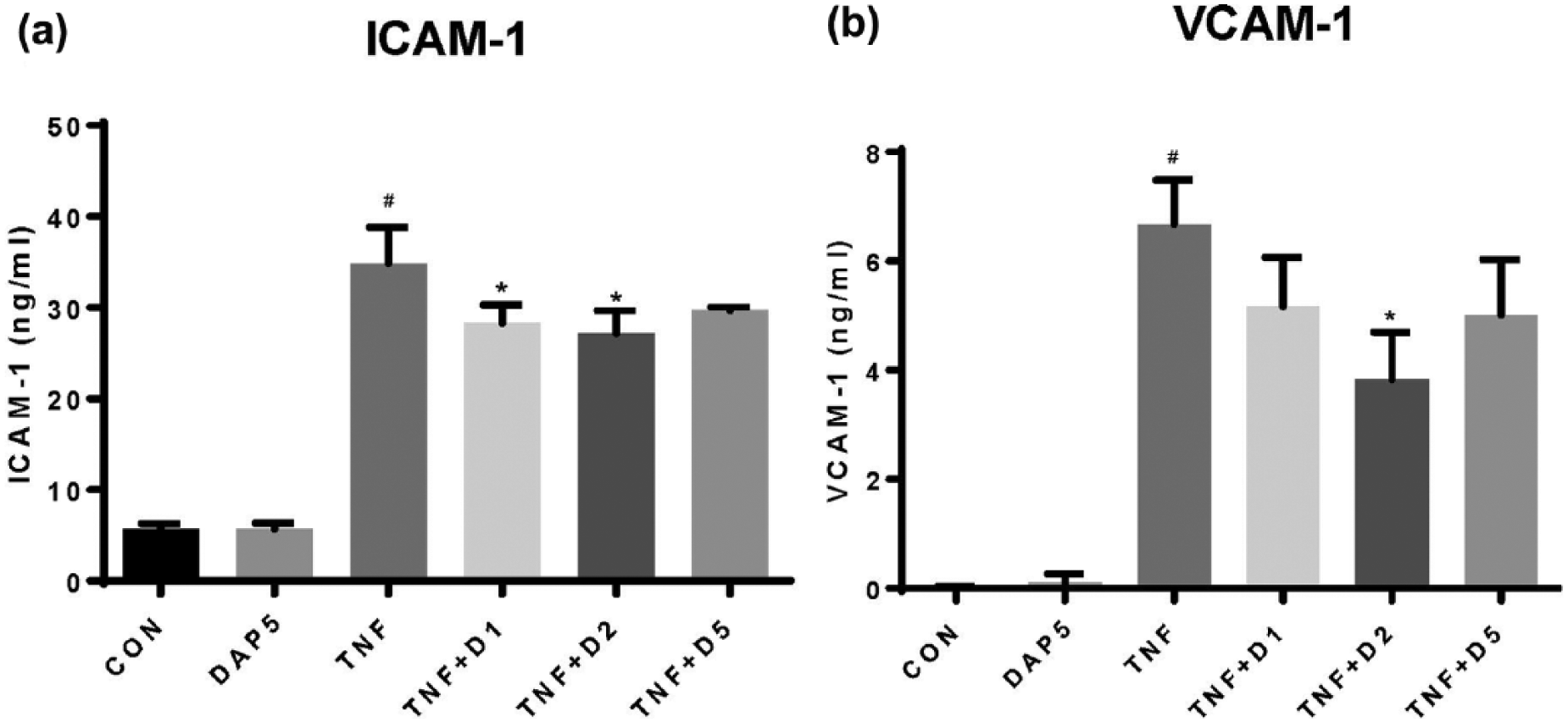
DAPA treatment attenuates 24 h TNFα induction of (a) ICAM-1 and (b) VCAM-1 protein expression in HUVECs.
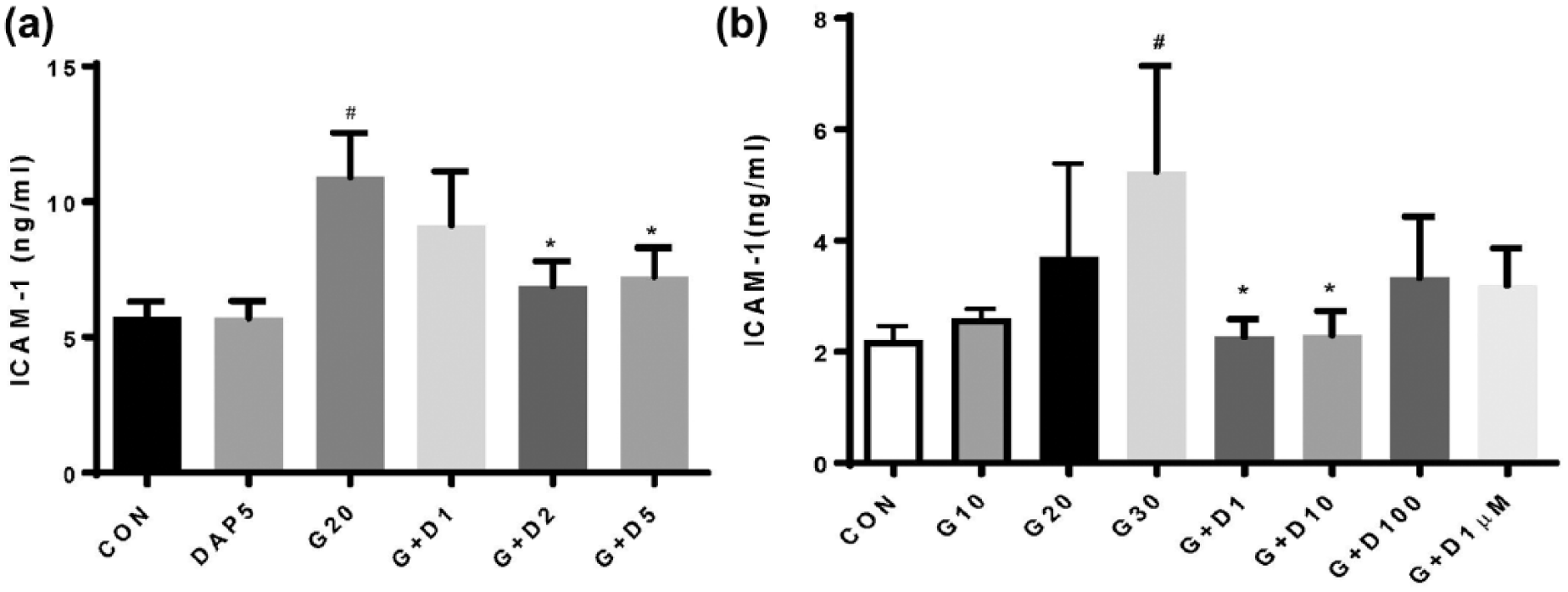
(a) Low-dose and (b) not high-dose DAPA treatment attenuates hyperglycaemia-induced ICAM-1 protein expression in HUVEC.
Significant induction of PAI-1 protein was identified after 24 h of TNFα stimulation compared with unstimulated HUVEC (Supplementary Figure 1). Addition of 100 nM DAPA to TNFα-stimulated HUVEC for 24 h resulted in significant attenuation of TNFα-mediated induction of PAI-1 messenger RNA (mRNA) and protein expression (Supplementary Figure 1).
DAPA attenuates TNFα-mediated induction of NFκB expression in human vascular endothelial cells
Given the restricted renal expression of SGLT-2 the target molecule for DAPA, with previous studies failing to identify SGLT-2 expression in either vascular endothelial or vascular smooth muscle cells, 12 we investigated a potential alternative molecular mechanism for the observed effects of DAPA on vascular peptide expression.
Further evaluation of a potential molecular mechanism of DAPA-mediated modulation of induced adhesion molecule expression was undertaken in HUVECs. Significant induction of NFκB mRNA was identified after 24 h of TNFα stimulation compared with unstimulated HUVEC (Figure 3). Addition of DAPA (1.0–2.0 nM) to TNFα-stimulated HUVEC for 24 h resulted in a significant attenuation of TNFα-mediated induction of NFκB expression (Figure 3) suggesting a potential transcriptional mechanism for DAPA-mediated regulation of vascular adhesion molecule expression.
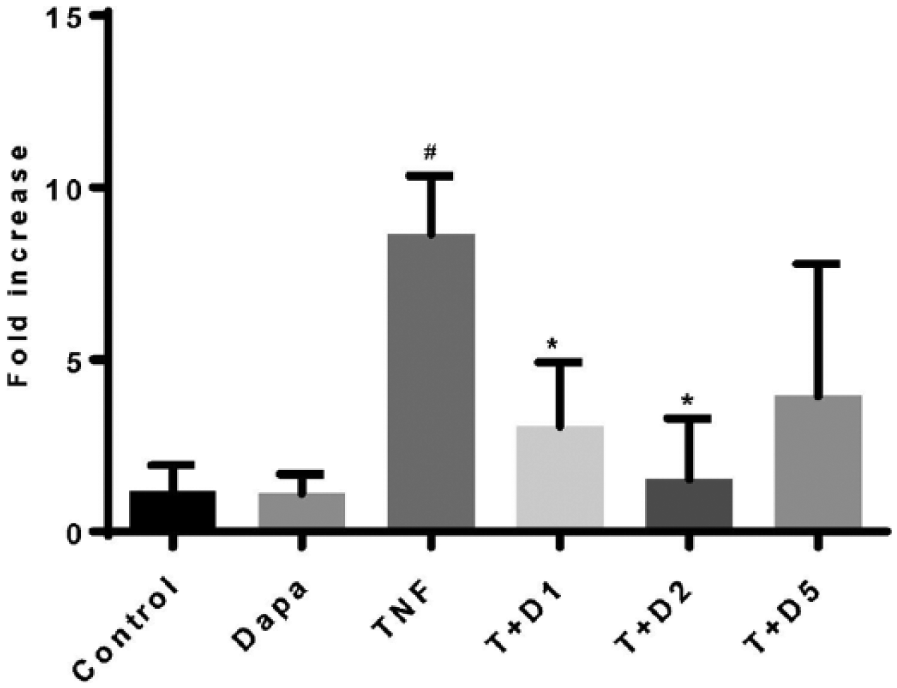
DAPA treatment attenuates 24 h TNFα-induced NFκB mRNA expression in HUVEC.
Acute DAPA treatment induces significant endothelial-independent vasorelaxation
Abdominal aortic rings from C57Bl/6J mice maintained on a normal chow diet were utilised to evaluate direct vasorelaxation responses evoked by acute DAPA administration (1.0 nM–10 µM). ACh-induced relaxation (10 µM) was reduced in aortic rings with denuded endothelium compared to aortic rings with intact endothelium (Figure 4(a)). Acute DAPA treatment induced direct, endothelial cell-independent, vasorelaxation in aortic rings (Figure 4b) with no difference in –LogEC50 or Emax between curves (Table 1). No effect of endothelial denudation on SNP-induced vasorelaxation of the same aortic rings was observed (Figure 4C). .
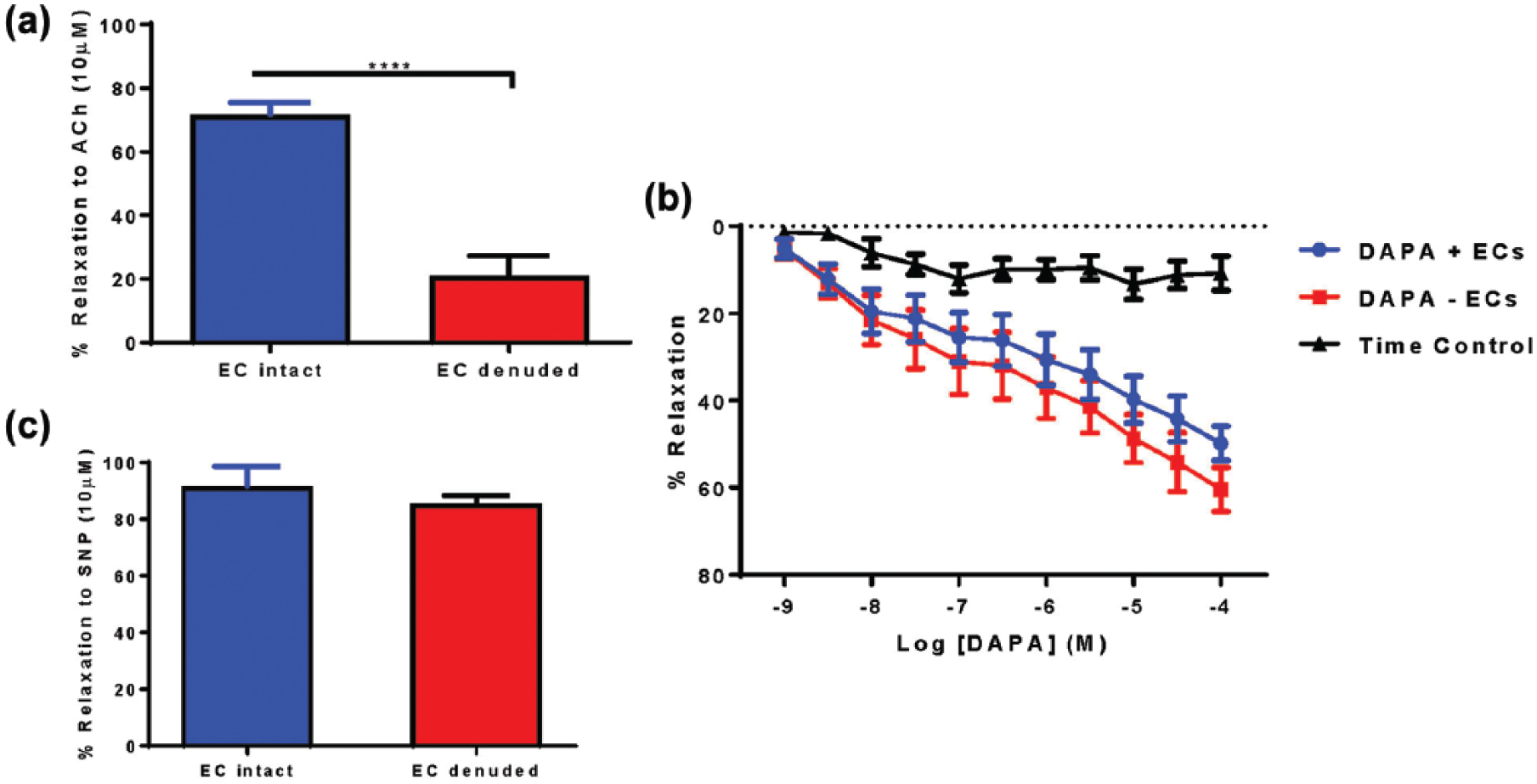
Acute DAPA treatment induced direct vasorelaxation in aortic rings from C57Bl/6J mice. (a) ACh-induced relaxation (10 µM) was reduced in aortic rings with denuded endothelium compared to aortic rings with intact endothelium. ****p < 0.0001, unpaired t-test, n = 4–7. (b) DAPA + EC = DAPA treatment with intact endothelium, DAPA − EC = DAPA treatment with denuded endothelium, n = 5–9. (c) EC denudation did not affect SNP-induced vaso-relaxation in aortic rings.
Effect of DAPA-induced changes in pD2, Emax and AUC in mouse aortic rings from the various vascular reactivity models.
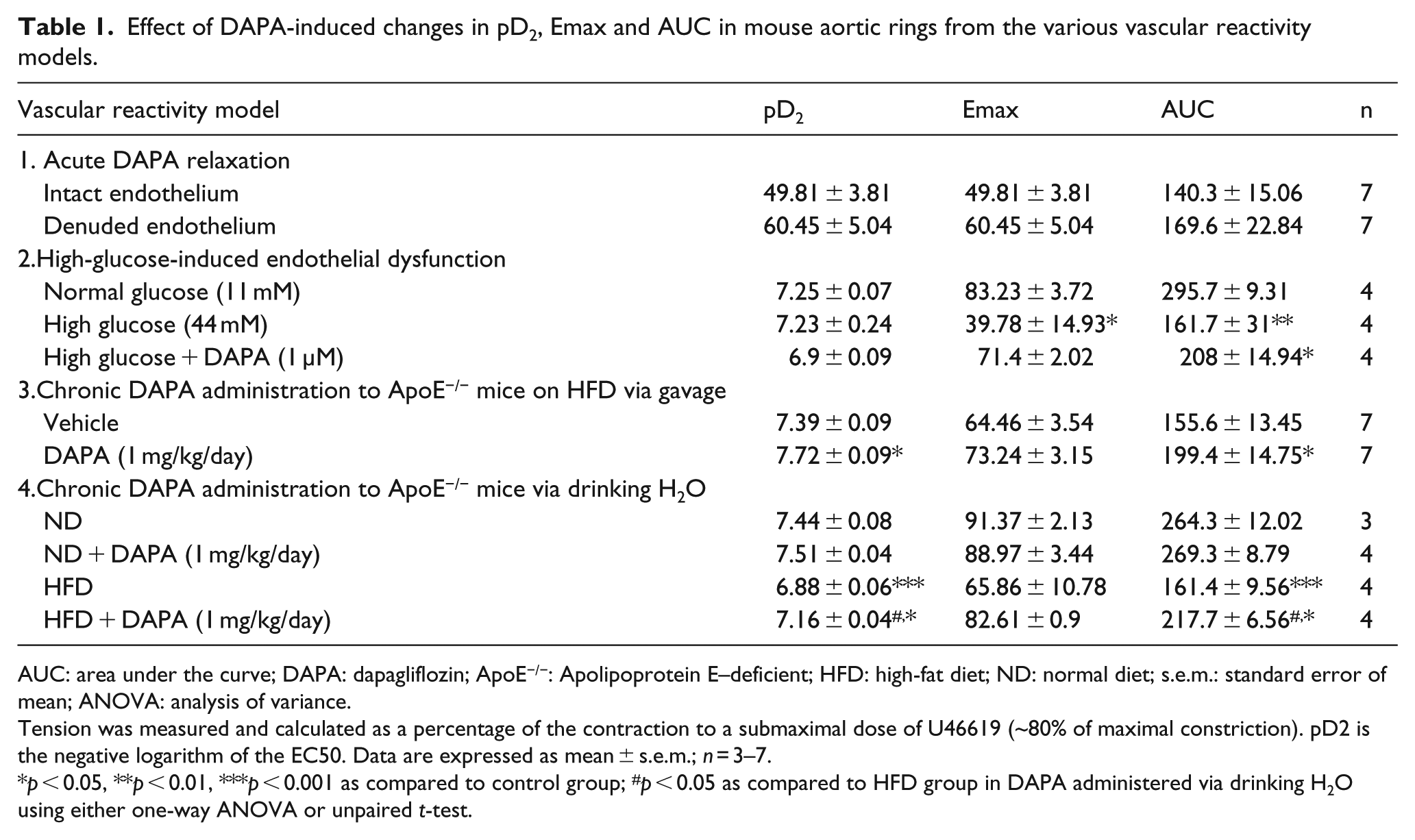
AUC: area under the curve; DAPA: dapagliflozin; ApoE−/−: Apolipoprotein E–deficient; HFD: high-fat diet; ND: normal diet; s.e.m.: standard error of mean; ANOVA: analysis of variance.
Tension was measured and calculated as a percentage of the contraction to a submaximal dose of U46619 (~80% of maximal constriction). pD2 is the negative logarithm of the EC50. Data are expressed as mean ± s.e.m.; n = 3–7.
p < 0.05, **p < 0.01, ***p < 0.001 as compared to control group; #p < 0.05 as compared to HFD group in DAPA administered via drinking H2O using either one-way ANOVA or unpaired t-test.
Evaluation of endothelial-dependent vasorelaxation in response to ACh administration (1 h) was performed on abdominal aortic rings subjected to 4-h hyperglycaemia- (44.0 mmol/L) induced endothelial dysfunction (Figure 5, Table 1). Acute DAPA administration (1-h preincubation) demonstrated a trend towards improved endothelial function (Figure 5(a), Table 1). Acute hyperglycaemia had no direct effect on vascular smooth muscle cell relaxation (Figure 5(b)).
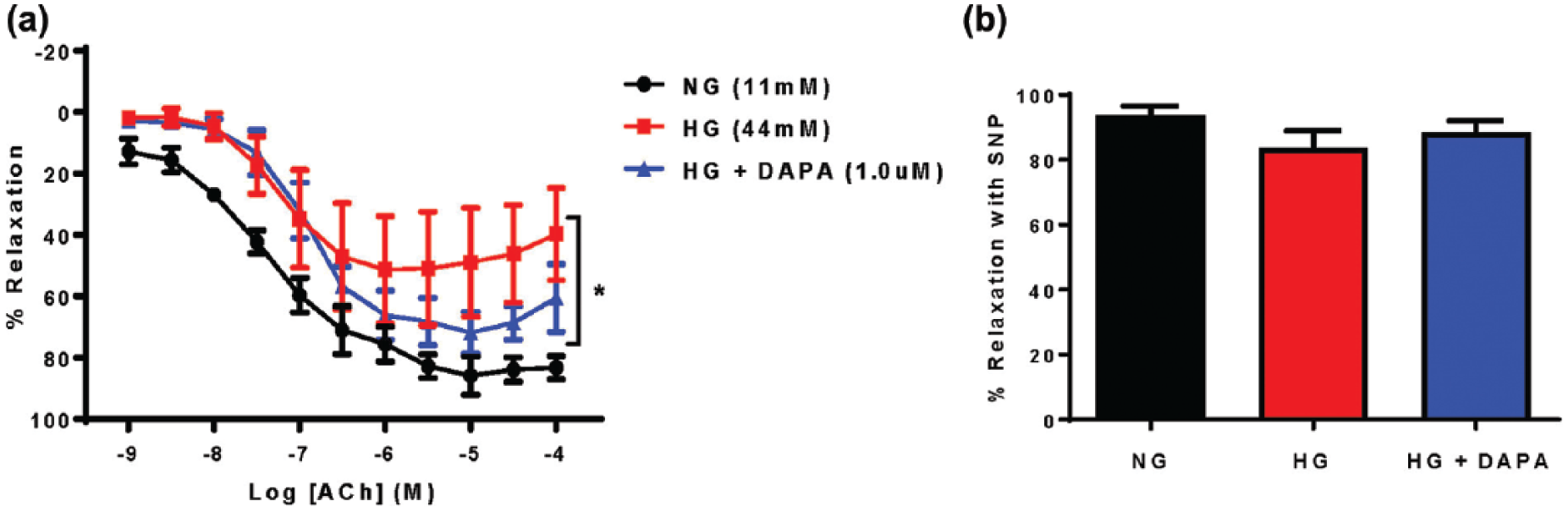
Acute DAPA treatment attenuated hyperglycaemia-induced endothelial dysfunction in isolated aortic rings from C57Bl/6J mice. (a) Acute DAPA treatment (1.0 µM) reduced hyperglycaemia-induced endothelial cell dysfunction. NG = normal glucose Krebs (11 mM), HG = high-glucose Krebs (44 mM), HG + DAPA = HG + DAPA 1.0 µM. n = 4; *p < 0.05 versus NG, two-way RM-ANOVA. (b) HG or HG + DAPA treatment had no effect on SNP-induced endothelial-independent relaxation (n = 4).
Chronic DAPA treatment attenuates endothelial dysfunction and expression of vascular adhesion molecules in adult and aged ApoE−/− mice
Chronic (4 weeks, 1.0 mg/kg/day) DAPA treatment via gavage in HFD-fed ApoE−/− adult mice had no significant effect on body weight (Figure 6(a)) or glucose clearance following a glucose tolerance test (Figure 6(b)). There was a significant improvement in endothelial-dependent vasorelaxation in aortae from DAPA-treated ApoE−/− mice compared to vehicle controls (Figure 6(c), Table 1) while endothelium-independent vasorelaxation mediated by SNP was not affected by either HFD or chronic DAPA treatment (Figure 6(d)). The vaso-protective effect of chronic DAPA treatment was independent of a significant change in atherosclerotic plaque area (Figure 6(e)).
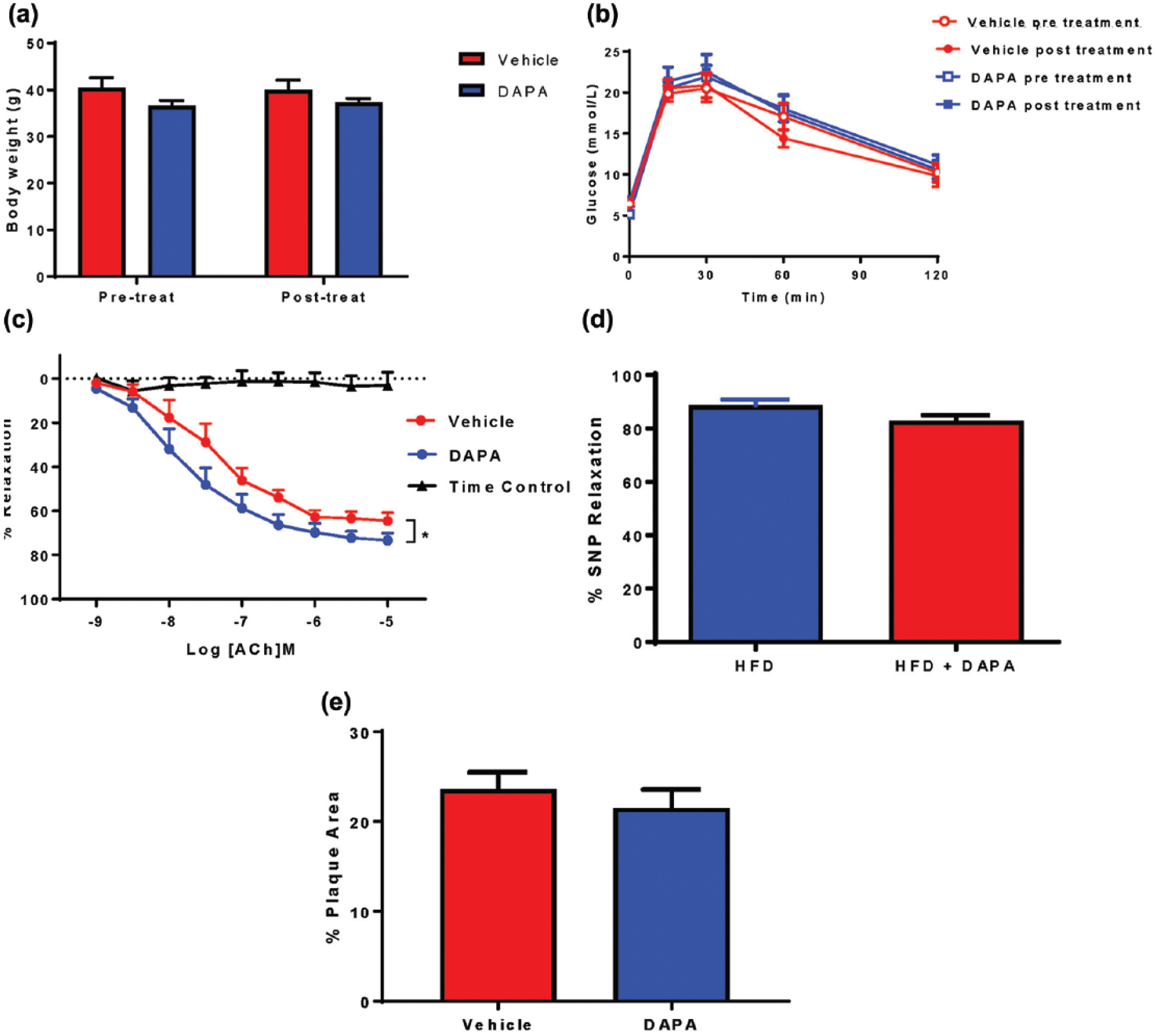
Chronic DAPA treatment (via gavage) attenuated HFD-induced endothelial cell dysfunction in Adult ApoE−/− mice. Chronic DAPA treatment had no effect on (a) body weight or (b) glucose clearance; (c) there was a significant improvement in ACH-induced endothelium-dependent vasorelaxation. n = 7. *p < 0.05 versus vehicle, two-way RM-ANOVA. (d) Endothelium-independent vasorelaxation mediated by SNP was not affected by either HFD or chronic DAPA treatment (HFD + DAPA). (e) Chronic DAPA treatment did not change aortic plaque area assessed by Oil Red O en face staining. n = 7.
Chronic DAPA treatment (4 weeks, 1.0 mg/kg/day) given in drinking water significantly attenuated HFD-induced endothelial cell dysfunction in aged ApoE−/− mice similar to that seen when DAPA was administered via gavage in younger adults (Figure 7(a)). The vaso-protective effects were again independent of any changes in progression of atherosclerotic lesion development (Figure 7(b)). Evaluation of inflammatory markers using immunofluorescence (Figure 7(c)) demonstrated reduced NFκB activation assessed as decreased P-IκBα expression in aortic sections from DAPA-treated mice. This was correlated with decreased ICAM-1 and F4/80 expression indicating reduced macrophage infiltration into aortic sections of ApoE−/− mice fed a HFD and chronically treated with DAPA (4 weeks, 1.0 mg/kg/day).
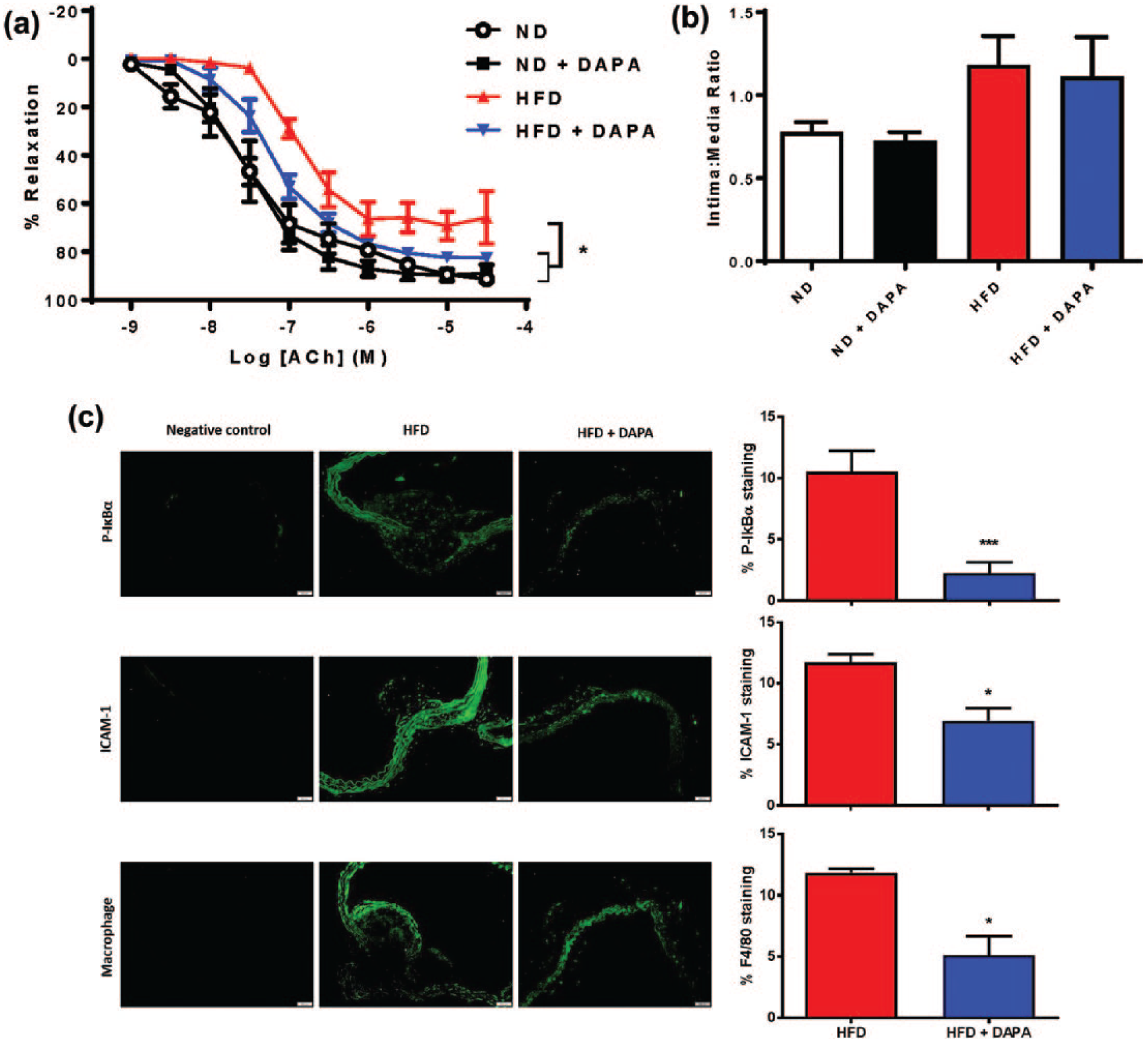
Chronic DAPA treatment (via drinking water) attenuated HFD-induced endothelial cell dysfunction and inflammatory mediators in Aged ApoE−/− mice. (a) ND = ApoE normal diet, ND + DAPA = ND + DAPA-treated, HFD = ApoE high-fat diet, HFD + DAPA-treated. n = 3–4. *p < 0.05 versus HFD; (b) DAPA treatment had no effect in either normal diet-fed (ND) or high-fat diet-fed (HFD) mice on atherosclerotic neointima:media ratio; (c) chronic DAPA treatment attenuated HFD-induced P-IκBα, ICAM-1 and F4/80 expression, ×20 magnification. n = 3–4.
Discussion
Recent cardiovascular safety studies suggest a potential cardiovascular benefit from treatment with the SGLT-2 inhibitor empagliflozin in the setting of type 2 diabetes; 2 however, delineation of the mechanisms responsible for these effects and confirmation of a class effect are still under investigation. Our in vitro, ex vivo and in vivo studies investigated the effects of the SGLT-2 inhibitor DAPA in models of vascular reactivity and early atherogenesis. Results from our studies suggest that DAPA is able to (a) attenuate endothelial cell activation in vitro, (b) induce direct ex vivo vasorelaxation and (c) reduce endothelial cell dysfunction and molecular changes associated with early atherogenesis under conditions of chronic in vivo administration. In addition, we postulate a molecular and cellular mechanism involving DAPA-mediated regulation of NFκB expression and macrophage infiltration which may, in part, be responsible for our observations.
Pre-clinical evidence implicating members of the SGLT-2 inhibitor class of agents in modulation of vascular endothelial cell activation and endothelial cell dysfunction, early steps in the atherogenic process, points to a potentially complex role for these agents in regulation of vascular function. Strikingly, we found that DAPA exerted a marked vasorelaxant effect in aortic tissue, which is consistent with a recent report in C57BL/6 mice suggesting that SGLT-2 inhibitors directly induce vasorelaxation although this observation was vascular bed-dependent since vasorelaxation occurred in pulmonary but not coronary arteries acutely, while chronic in vivo SGLT-2 inhibitor administration enhanced coronary artery, but not pulmonary, vasorelaxation evoked by SNP. 12 Our observations also identify complex effects of DAPA when administered acutely or chronically. For example, DAPA exerted direct endothelium-independent effects, yet also tended to improve ACh-mediated vasorelaxation during acute, 44 mmol/L hyperglycaemia, an informative, although extreme, representation of diabetes pathophysiology. In contrast, chronic DAPA administration improved endothelial function in both adult and aged ApoE−/− mice. In the context of chronic DAPA administration, a sustained reduction in plasma glucose concentration may explain improvements in endothelial cell function. However, given ApoE−/− mice used in our studies were non-diabetic, with no change in glucose handling evoked by DAPA, a potentially novel molecular mechanism for DAPA-mediated regulation of endothelial cell dysfunction required exploration. Previous studies have failed to identify SGLT-2mRNA expression in vascular endothelial and vascular smooth muscle cells 12 and suggest an alternate molecular mechanism may be implicated in DAPA-mediated modulation of endothelial cell dysfunction and vascular peptide expression. Given oxidative stress is a hallmark of endothelial cell dysfunction 13 and reactive oxygen species (ROS) mediate effects on vascular endothelial cell dysfunction via activation and modulation of expression of components of the NFκB pathway, 14 we investigated the in vitro effects of DAPA on TNFα-induced NFκB mRNA expression. DAPA-mediated attenuation of TNFα-induced NFκB mRNA expression implicates DAPA in transcriptional regulation, a novel observation, and in keeping with recently identified ‘off target’ effects involving activation of cell signalling pathways independent of SGLT-2 by other members of the SGLT-2 class of agents.5,15
Endothelial cell dysfunction is identified as a significant contributor to early atherogenesis particularly in the vulnerable type 2 diabetic patient population.16,17 Upregulation of adhesion molecule expression is a marker of endothelial cell dysfunction and a precursor to the development of atherosclerosis. 18 Our in vitro observations of DAPA-mediated inhibition of TNFα and hyperglycaemia-induced adhesion molecule expression together with in vivo attenuation of endothelial dysfunction and HFD-induced upregulation of ICAM-1 expression, P-IκB and macrophage infiltration in ApoE−/− mice would suggest a potential benefit of DAPA in modulating the onset and progression of atherogenesis and a potential immunomodulatory role for DAPA although these findings require further investigation regarding underlying molecular mechanisms, particularly poignant given the previously identified absence of SGLT-2 expression in our in vitro endothelial cell systems. 12
The absence of an effect of DAPA on atherosclerotic lesion burden in our preliminary in vivo studies suggests that the hypoglycaemic effects of DAPA may be required to elicit, at least in the shorter term, DAPA-mediated attenuation of atherosclerotic burden in the ApoE−/− mouse model although larger studies with an increased sample size are required to confirm this contention and represent a limitation of this study. Indeed, recent pre-clinical studies support our preliminary observations with identification of reduced atherosclerotic burden in diabetic ApoE−/− mice treated with DAPA with no significant reductions in non-diabetic ApoE−/− mice.4,5 However, in vitro and in vivo attenuation in vascular peptide expression, improvements in endothelial function, non-significant reductions in weight together with previously documented attenuation of expression of inflammatory mediators and molecules including NFκB and interleukin-1 (IL-1)β in the non-diabetic ApoE−/− mouse model 5 may all combine to produce reductions in atherosclerotic disease burden over extended time periods independently of improvements in hyperglycaemia.
Given the results of large-scale cardiovascular safety studies with the SGLT-2 inhibitor empagliflozin suggest potential cardiovascular benefits 2 together with recent observations of improvements in vascular endothelial function in early-stage type 2 diabetic patients treated with DAPA, 19 our pre-clinical observations identifying DAPA-mediated effects on vascular reactivity and endothelial cell activation may portend potential cardiovascular benefits from DAPA treatment. The results of cardiovascular safety studies in the setting of type 2 diabetic patients treated with DAPA 6 will further elucidate the cardiovascular effects of this agent in the clinical arena.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study was sponsored, in part, by a scientific research grant from AstraZeneca AB, Sweden. AstraZeneca AB, Sweden, had no role in the study design or article production.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.