
Abstract
Objective:
Metabolic syndrome is a common disorder that predisposes to both cardiovascular disease and diabetes. There is paucity of data on cellular signal transduction pathways in metabolic syndrome. This study determined monocyte mitogen-activated protein kinase activity in patients with metabolic syndrome.
Research design and methods:
The p38, extracellular signal-regulated kinase-1/2 and Jun N-terminal kinase–mitogen-activated protein kinase activities were assayed in isolated monocytes from patients with metabolic syndrome and controls (n = 36 per group) and correlated with features of metabolic syndrome, inflammation and oxidative stress biomarkers.
Results:
A significant increase in p38 mitogen-activated protein kinase activity was observed in metabolic syndrome even following adjustment for adiposity. There were no significant differences in extracellular signal-regulated kinase-1/2 and Jun N-terminal kinase activities. P38 mitogen-activated protein kinase activity correlated significantly with homeostasis model assessment–estimated insulin resistance and biomarkers of inflammation and oxidative stress.
Conclusions:
We are first to observe a selective increase in monocyte p38 mitogen-activated protein kinase activity in metabolic syndrome and suggest it as a pivotal molecular target for ameliorating insulin resistance and inflammation.
Metabolic syndrome (MetS) affects 35% of US adults and confers an increased risk for diabetes and cardiovascular disease (CVD). 1 Circulating monocyte/macrophages and macrophages in adipose tissue contribute to both the pro-inflammatory state and insulin resistance of MetS. 2 However, studies investigating signal transduction pathways in MetS are sparse. Hence, we studied the critical mitogen-activated protein kinase (MAPK) pathways, which transduce external cues into important cellular responses including inflammation, in monocytes from patients with MetS. Since we have previously shown increased circulating and cellular biomarkers/biomediators of inflammation and oxidative stress in patients with MetS and shown correlations between these biomediators with insulin resistance, a role could be posited for these biomediators in the pathobiology of MetS since this is also supported in animal models.2–4
Research design and methods
Patients and methods
Subjects (aged 21–70 years) with MetS (n = 36) and healthy controls (n = 36) were recruited from Sacramento County using procedures as described previously.3,4 Hence, this report is on the same cohort. MetS was defined using the criteria of the National Cholesterol Education Program Adult Treatment Panel III (NCEP ATP III). 1 Briefly, nascent MetS is defined as patients with MetS without diabetes (fasting plasma glucose <125 mg/dL) and HbA1C < 6.5% (<47.5 mmol/mol) or clinical CVD (coronary artery disease, peripheral arterial disease, cerebrovascular disease) as reported previously.3,4 Control subjects had to have ⩽2 features of MetS and not be on blood pressure medications. Other exclusion criteria for control subjects were fasting plasma glucose (>100 mg/dL) and triglycerides (TGs; >200 mg/dL).
None of the subjects in both groups were on statins, angiotensin receptor blockers, fibrates, aspirin or peroxisome proliferator-activated receptor (PPAR) gamma agonists. MetS patients were on diuretics, beta blockers or angiotensin-converting enzyme (ACE) inhibitors (n = 2) for their hypertension. All subjects had a high-sensitive C-reactive protein (hsCRP) <10 mg/L and normal white cell and neutrophil counts and had no acute or chronic inflammatory diseases or recent infection. All other exclusion criteria including smoking, albuminuria >30 mg/G creatinine and clinical CVDs are detailed in previous publications.3,4
Informed consent was obtained from participants in the study, which was approved by the institutional review board at the University of California Davis. All human investigation was conducted according to the principles expressed in the Declaration of Helsinki. After history and physical examination, fasting blood was obtained for basic chemistries, hsCRP, homeostasis model assessment (HOMA) calculation and monocyte isolation as described previously.3,4
Monocyte isolation and cell signalling studies
Monocytes were isolated from fasting heparinized blood by Ficoll-Hypaque density gradient centrifugation followed by negative magnetic separation as described previously. 3 Using CD14 staining, >92% of the cells were identified to be monocytes. Monocyte lysates and nuclear extracts were prepared as described previously. 3 Phosphorylated and total p38 MAPK, extracellular signal-regulated kinase-1/2 (ERK1/2), and c-Jun N-terminal kinase (JNK), activity in the cell lysates and nuclear factor (NF)-κB p65 activity in the nuclear extracts were assessed using reagents from Bio-Rad using the Bioplex multiplex phosphoprotein detection assays following the manufacturer’s instructions. 5 We have previously validated these multiplex assays with western blots. 5 The principle of these bead-based assays is similar to a capture sandwich immunoassay. The assays use microscopic beads each with a different colour code (spectral address) to permit discrimination of individual phospho and total proteins. Briefly, 50 µL of internally dyed microbeads (1×) each coupled to antibodies directed against a different target phosphoprotein or total protein was added per well of 96-well plate. Thereafter, 25 µL of the cell lysate was added to each well and incubated overnight at 25°C on a plate shaker. Following repeated washing, 25 µL of biotin-labelled detection antibodies for each target was added per well of the plate for 30 min. Streptavidin–phycoerythrin conjugated fluorescent reporter was added to each well of the plate in dark following further washes. Data were acquired on multiplex plate reader. The dyed beads were identified by their internal fluorescent signature, and the level of target bound to beads was indicated by the intensity of reporter signal for the phospho and total MAPKs. The intra-assay coefficients of variations of the assays were <14%.
Statistical analysis
Results are expressed as mean and standard deviation (SD) or as median and interquartile range for skewed variables. Log transformations were applied to variables with skewed distributions prior to parametric analyses. Comparisons between the control and MetS groups were made with two-sample t-tests and analysis of covariance to control for age, body mass index (BMI) and waist circumference (WC). The association of phospho-p38 MAPK with the number of features of MetS and p trend was derived using the Jonckheere–Terpstra test. Combining the control and MetS groups, Spearman’s rank correlation coefficients were computed to assess the association of metabolic variables and the phospho-p38 MAPK variables. Data were analysed using SAS version 9.4 (SAS Institute, Cary, NC).
Results
As shown in Table 1, patients with MetS were balanced for age and gender but had significant differences in all features of MetS and increased hsCRP and homeostasis model assessment–estimated insulin resistance (HOMA-IR).
Salient baseline characteristics.
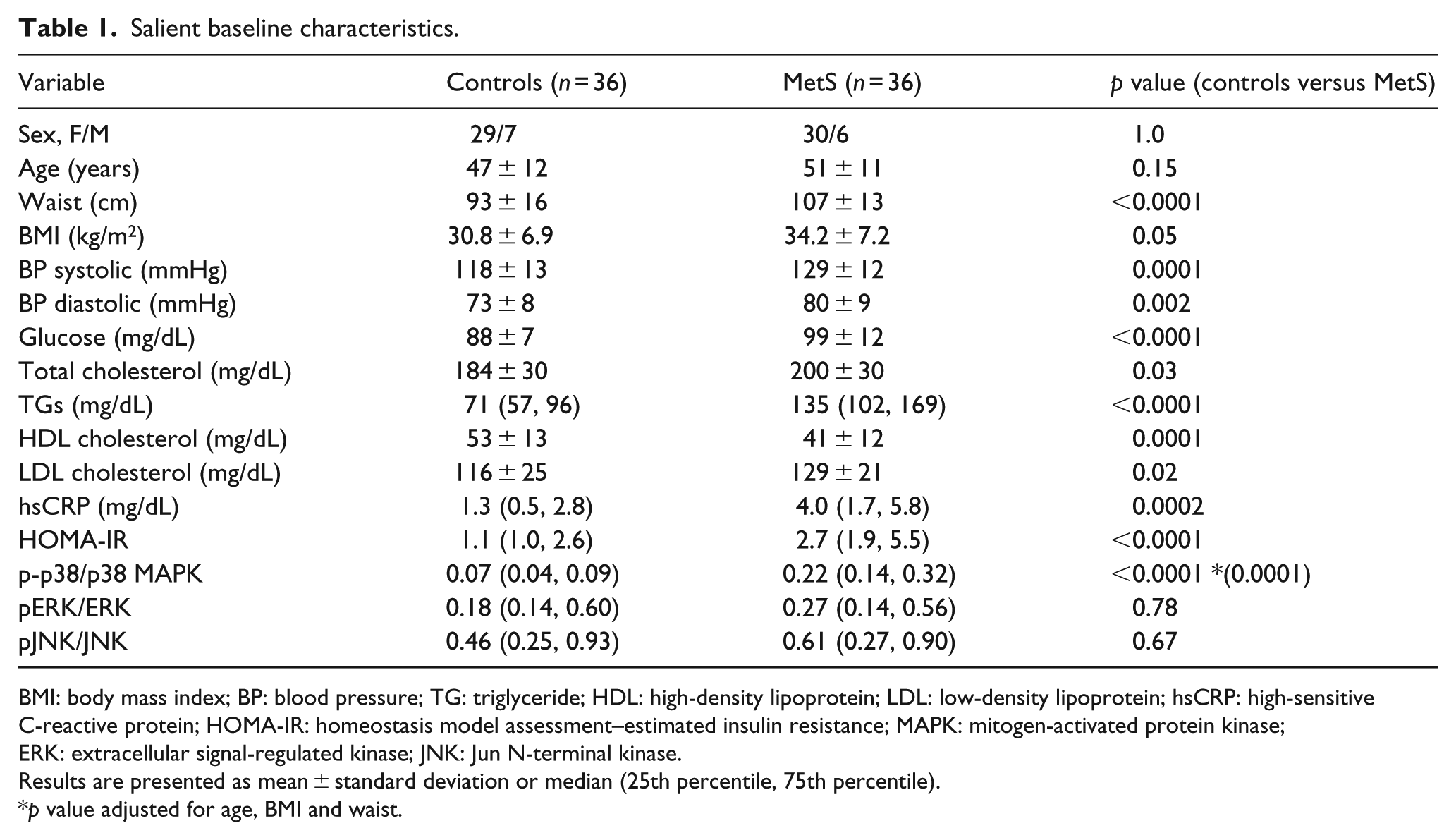
BMI: body mass index; BP: blood pressure; TG: triglyceride; HDL: high-density lipoprotein; LDL: low-density lipoprotein; hsCRP: high-sensitive C-reactive protein; HOMA-IR: homeostasis model assessment–estimated insulin resistance; MAPK: mitogen-activated protein kinase; ERK: extracellular signal-regulated kinase; JNK: Jun N-terminal kinase.
Results are presented as mean ± standard deviation or median (25th percentile, 75th percentile).
p value adjusted for age, BMI and waist.
With respect to monocyte MAPKs, there was a significant increase in phospho-p38/p38 MAPK ratio in MetS which persisted following adjustment for BMI and WC with no significant differences for pERK/ERK1/2 and pJNK/JNK (Table 1). Also phospho-p38 MAPK increased with increasing features of MetS (p < 0.001 for trend) and correlated significantly with all features: WC (r = 0.35, p = 0.002), glucose (r = 0.46, p < 0.001), TGs, (r = 0.53, p < 0.001), blood pressure systolic (r = 0.38, p = 0.001) and high-density lipoprotein (HDL) cholesterol (r = −0.27, p = 0.02).
Since we have reported increased inflammation and oxidative stress previously,3,4 correlations were undertaken between these variables and p38 MAPK activity. Phospho-p38 MAPK correlated significantly with HOMA-IR (r = 0.40, p = 0.005), hsCRP (r = 0.33, p = 0.005), interleukin-6 (IL-6; r = 0.38, p = 0.0009), interleukin-8 (IL-8; r = 0.32, p = 0.006), soluble tumour necrosis factor receptor-1 (sTNFR-1; r = 0.49, p = 0.001), toll-like receptor 2 (TLR2; r = 0.30, p = 0.01), toll-like receptor 4 (TLR4; r = 0.39, p = 0.001), phospho-NF-κB p65 (r = 0.56, p < 0.001), basal superoxide secretion (r = 0.33, p = 0.004) and oxidized low-density lipoprotein (LDL) levels (r = 0.60, p > 0.001). There was a trend to significance with interleukin-1 (IL-1; r = 0.23, p = 0.05).
Since two of our patients in this report were on ACE inhibitors which are known to inhibit MAPKs,6,7 we repeated all analyses following exclusion of these two patients. All the above repeated findings remained the same.
Conclusion
There are scanty data with respect to cell signalling in MetS. Previously, it has been shown that there is altered mRNA expression or protein abundance of receptors on monocytes of patients with MetS. 2 To date, an increase in CD40, CD32, CD64, CCR5 and TLR2 and TLR4 with a decrease in PPAR alpha and delta and the Vitamin D receptor has been reported. 2 However, signal transduction was only pursued with respect to monocyte TLR2 and TLR4 showing increased NF-κB activity and downstream biomediators. 3 Since the TLRs also signal through MAPKs, 8 we investigated MAPK activity in this report. The three well-defined sub-groups of MAPKs in mammalian cells are the ERK1/2, the JNK and the p38 MAPKs. 9
We show a significant increase in p38 MAPK activity even following adjustment for both BMI and WC. However, we failed to show an increase in ERK1/2 and JNK. The major focus of studies on the MAPK family is based on their role in impairing insulin signalling. 10 In a similar model of polycystic ovary syndrome in women, they showed an increase in basal ERK1/2 activity in skeletal muscle but no change in p38 activity. 11 However, in diabetic patients, in both skeletal muscle and adipose tissue, increase in p38 MAPK activity was reported.12,13 Koistinen et al. 12 showed an increase in basal p38 activity in skeletal muscle. Carlson et al. 13 showed a significant increase in basal p38, JNK and ERK1/2 with the greatest increase in p38 (5-fold) which contributed to loss of glucose transporter type 4 (GLUT4) expression. We also show in MetS a significant correlation with insulin resistance as determined by HOMA-IR.
P38 MAPK activity has been suggested to contribute to inflammation.14–16 We show that p38 MAPK activity correlated with biomediators of increased inflammation: TLR2 and TLR4, CRP, NF-κB activity, IL-6, IL-8 and sTNFR-1 in patients with MetS. Hence, in patients with MetS, this could be an additional molecular target to attenuate both insulin resistance and inflammation, the major pathophysiology underlying this disorder.1,2
To date, it is unclear what triggers the increase in p38 MAPK activity. It has been suggested that oxidative stress is one such stressor. 9 In our study, we show a significant correlation with both oxidized LDL and basal nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) oxidase activity (superoxide anion). While correlations cannot infer causality, this data support the link proposed in the published literature.
In conclusion, in this first study of MAPK activity in MetS, we make the original observation of a selective increase in p38 MAPK activity in MetS and thus present a novel molecular target to ameliorate both insulin resistance and inflammation.
Footnotes
Acknowledgements
I.J. participated in the study design, wrote the manuscript and read and approved the final manuscript; B.A.H. performed all the statistical analyses and read and approved the final manuscript and R.P. drafted the manuscript and read and approved the final manuscript. I.J. is the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis. Authors acknowledge assistance of Sridevi Devaraj, PhD, for performing the MAP kinase assays.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
This study was supported by an ADA grant (I.J.).