
Abstract
Background:
The regulation of miR-126 by rs4636297 single nucleotide polymorphism (SNP) has been implicated in the pathogenesis of neovascularisation by promoting vascular endothelial growth factor, suggesting it could be associated with sight threatening diabetic retinopathy (STDR), but has not been previously investigated or reported.
Materials and methods:
A case control study of 531 individuals with diabetes was genotyped for the rs4636297 SNP, using the Sequenom iPLEX Gold chemistry. STDR included people with severe non-proliferative diabetic retinopathy (NPDR) or proliferative diabetic retinopathy (PDR). Association was tested using logistic regression analysis, adjusting for confounding variables.
Results:
In an additive model, the A allele of rs4636297 SNP is significantly associated with STDR compared to people with none or mild diabetic retinopathy (DR) (odds ratio (OR) = 2.02, 95% confidence interval (CI) = 1.22–3.35, p = 0.006).
Conclusion:
The A allele of rs4636297, known to be the non-functional allele for post-translational regulation of miR-126, is associated with STDR. This finding suggests that this locus would be a potential therapeutic target for inhibiting the development of DR.
Introduction
Diabetic retinopathy (DR) is a common sight threatening microvascular complication of diabetes mellitus (DM). Early stage disease, characterised by microaneurysm formation and small intraretinal haemorrhages, is often associated with preservation of vision. With severe retinal ischaemia abnormal retinal angiogenesis can develop and in turn lead to vitreous haemorrhage, fibrosis or tractional retinal detachment. This neovascularisation of the retinal microvasculature is clinically referred to as proliferative diabetic retinopathy (PDR).
The major known biochemical contributor to the pathogenesis of PDR is vascular endothelial growth factor (VEGF). The identification of increased intraocular VEGF in PDR has led to a breakthrough in the treatment of neovascularisation in patients with PDR via anti-VEGF therapies. 1 Recently, it has been identified that mature circulating microRNA-126 (miR-126) regulates the response of endothelial cells to VEGF in-vitro. 2 miR-126 has been implicated in the pathogenesis of neovascularisation involving endothelial cells, by promoting VEGF via the inhibition of Sprouty-related EVH1 domain-containing protein-1 (SPRED1) and phosphatidylinositol 3-kinase regulatory subunit beta (PIK3R2).2–4 Additionally, miR-126 has been shown to reduce the inflammatory response, via inhibiting vascular adhesion molecule-1 (VCAM-1) expression in the endothelium, thereby decreasing leukocyte interactions. 5 miR-126 involvement in neovascularisation and immune regulation within endothelial cells suggests that it is a key regulator of vascular function.5,6
The single nucleotide polymorphism (SNP) rs4636297 is located in an intronic region of the miR-126 terminal within the epidermal growth factor-like protein 7 (EGFL7) gene.6–10 This rs4636297 SNP post-transcriptional influences the expression of miR-126, and polymorphisms of rs4636297 SNP have differing inhibitory effects on circulating miR-126. 6 When the ancestral allele (G) of the rs4636297 SNP is present, it inhibits the processing of primary miR-126 to precursory miR-126, resulting in less circulating mature miR-126. 6 The less common A allele of rs4636297 has reduced function and is less effective at inhibiting the conversion of primary miR-126 to precursory miR-126. 6 This genetic variant alters the level of mature circulating miR-126, which promotes VEGF production. 6
An increase in VEGF is known to occur in ocular tissue during the development of PDR, 1 and the post-transcriptional regulation on VEGF from the rs4636297 SNP suggests that it is a prime candidate for driving microvascular dysfunction in DR. To understand the role of the rs4636297 SNP in DR, we hypothesised that the A allele variant of rs4636297 is associated with sight threatening diabetic retinopathy (STDR).
Materials and methods
Patients were recruited between March 2009 and December 2012 as part of the Diabetes Management Project (DMP) from clinics at the Royal Eye and Ear Hospital (RVEEH), Melbourne, VIC, Australia. 11 Ethical approval was provided by the RVEEH Human Research and Ethics Committee (08/815H) and informed consent was obtained from all participants. The DMP protocol adhered to the Declaration of Helsinki, and all privacy requirements were met.
Data collection
Each participant underwent an extensive examination, including comprehensive ocular and anthropometric measurements as well as completing socio-demographic and medical history questionnaires. Mid-stream urine samples were collected to assess albumin and creatinine levels. Peripheral blood samples were collected following a 12-h fast, and measurements of glycosylated haemoglobin (HbA1c), low-density lipoprotein (LDL), high-density lipoprotein (HDL), cholesterol, triglycerides and glucose were performed. All biochemical parameters were analysed at Melbourne pathology, Melbourne, VIC, Australia.
Phenotypic characterisation
Participants were defined as having type 1 diabetes if they were diagnosed as a juvenile and were insulin dependent from diagnosis, while those with a non-juvenile diagnosis or who did not require immediate insulin therapy were defined as type 2 diabetes. Severity of DR and diabetic macular oedema (DME) was obtained with the use of two-field, 45°, digital, non-stereo, colour fundus photographs from both eyes, collected using a non-mydriatic retinal camera (Canon CR6 – 45NM, Canon Inc., Tokyo, Japan). The presence of retinopathy was determined using the internationally recognised modified Airlie House classification as used in the Early Treatment of Diabetic Retinopathy Study protocol.12–14 The DR and DME assessment was independently validated by an ophthalmologist (PPC). Severity classification was based on the unilateral worst eye. DME was classified as those with signs of DME less than 2 disc diameter from the fovea. Classifications of retinopathy were selected based on two groups; the controls, those without DR or mild non-proliferative diabetic retinopathy (NPDR); and STDR, classified as those with severe NPDR/PDR (includes those previously treated with laser photocoagulation therapy for PDR).
SNP analysis
DNA was extracted using the QiaAmp Blood midi kit (Qiagen, Valencia, CA, USA). Participants were genotyped for the rs4636297 SNP using the Sequenom iPLEX Gold chemistry (Sequenom, San Diego, CA, USA) on an Autoflex mass spectrometer (Sequenom, Brisbane, QLD, Australia) at the Australia Genome Research Facility, Brisbane, QLD, Australia.
Structure of allelic variation
The nucleotide sequence variants for rs4636297 SNP were obtained from the Single Nucleotide Polymorphism Database (dbSNP) hosted by The National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/projects/SNP/). The sequence was utilised using default parameters in mfold (http://mfold.rna.albany.edu/?q=mfold/DNA-Folding-Form) to determine the predicted structural changes between allelic variants. To better understand the structural impact of rs4636297 on the genomic region, the surrounding nucleotides (AC_000141.1) were uploaded into KineFold (http://kinefold.curie.fr/) using the default parameters. Structures with the lowest free energy were selected for each allele from both programs. 15
Statistical analysis
All demographic parameters were compared using the Student t-test using Statistical Package for the Social Sciences (SPSS, version 19). Genetic data were analysed using PLINK v1.07. 16 The allelic and genotypic frequencies were initially compared using a χ2 test and was found to be in Hardy–Weinberg equilibrium. To understand the mode of inheritance, genotypic models of additive (A allele), dominant (AA and AG vs GG) and recessive (AA vs AG and GG) were applied. Multivariate logistic regression models were constructed and adjusted for: age, sex, duration of DM, HbA1c, body mass index, mean arterial pressure, cholesterol, triglycerides and microalbumin/creatinine ratio. A p value less than 0.05 was considered statistically significant in all analyses.
A total of 84% were of Northern-European descent. To test for any effect of ethnicity, allele frequencies between Caucasian and non-Caucasian individuals were compared. Given that no significant difference in allele frequency was identified (χ2 = 0.019; p = 0.892), all individuals were analysed together. Power calculations were conducted using case control for discrete traits analysis through the Genetic Power Calculator, 16 assuming a disease prevalence of 0.6% for STDR, a genotyped-to-causal variant linkage disequilibrium (LD) of D′ = 1, an identical marker-disease minor allele frequency (MAF) of 0.4, and an additive disease risk model, and using a significance threshold of α = 0.05. This study was adequately powered (>80%) to identify a heterozygotic effect (genotypic relative risk) of greater than 1.1 under an additive model for STDR and in the type 2 diabetes subgroup.
Results
The demographic characteristics of the study participants are presented in Table 1. There were 163 STDR cases and 368 control patients used for diabetes analyses. The sub-analyses for type 1 included 24 STDR cases and 23 controls, and for type 2 diabetes 138 STDR cases and 344 controls. Age, sex, duration of diabetes, and haematological levels of HbA1c, total HDL cholesterol ratio, fasting glucose, serum creatinine, the estimated glomerular filtration rate (EGFR), urinary microalbumin levels and the microalbumin/creatinine ratio differed significantly between patients with STDR and those patients with either no DR or only mild NPDR. To adjust for confounding effects, variables shown to impact DR were included simultaneously in a logistic regression model to assess the association of rs4636297 with DR. 17
Demographic features and clinical characteristics of the participants.
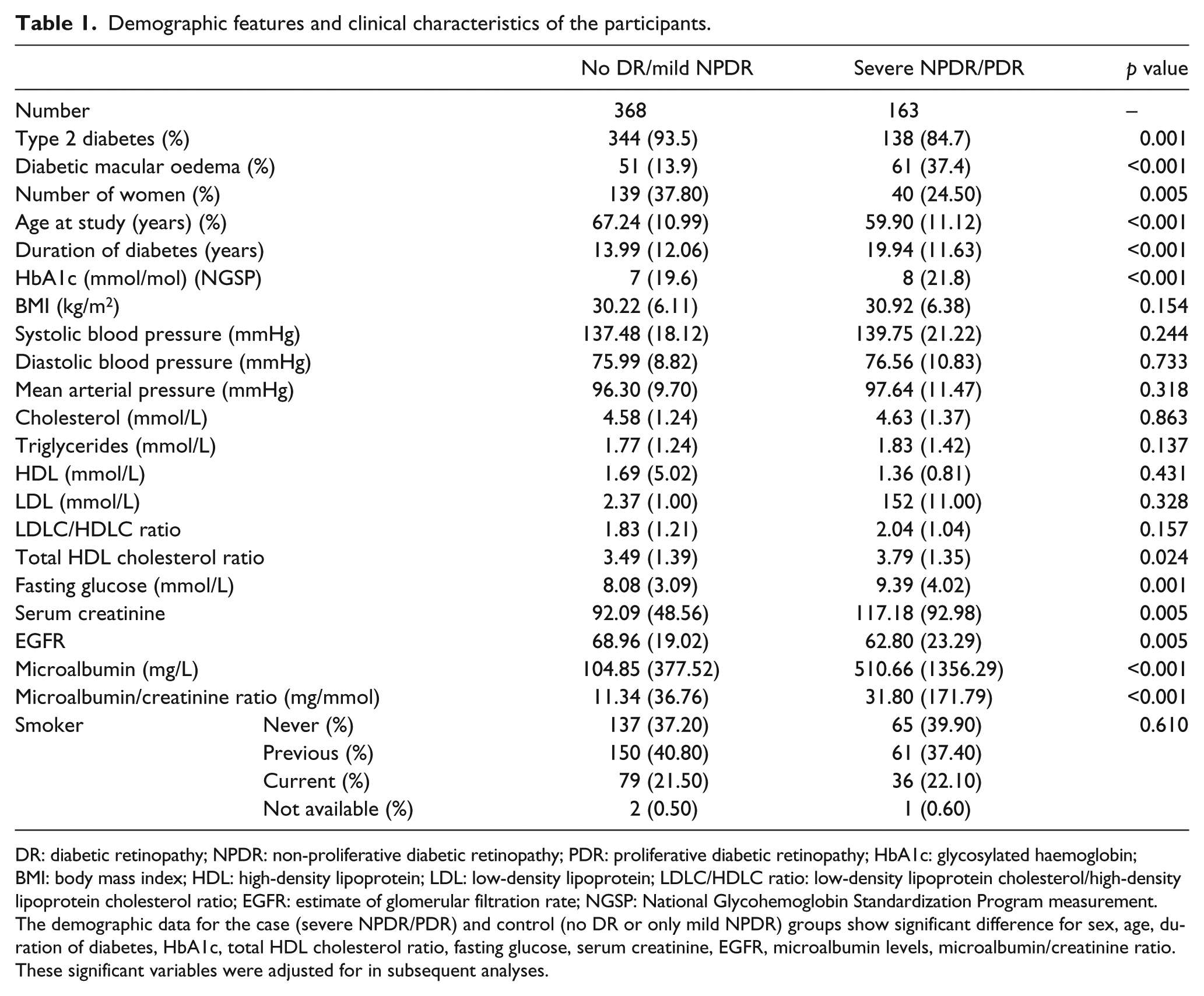
DR: diabetic retinopathy; NPDR: non-proliferative diabetic retinopathy; PDR: proliferative diabetic retinopathy; HbA1c: glycosylated haemoglobin; BMI: body mass index; HDL: high-density lipoprotein; LDL: low-density lipoprotein; LDLC/HDLC ratio: low-density lipoprotein cholesterol/high-density lipoprotein cholesterol ratio; EGFR: estimate of glomerular filtration rate; NGSP: National Glycohemoglobin Standardization Program measurement.
The demographic data for the case (severe NPDR/PDR) and control (no DR or only mild NPDR) groups show significant difference for sex, age, duration of diabetes, HbA1c, total HDL cholesterol ratio, fasting glucose, serum creatinine, EGFR, microalbumin levels, microalbumin/creatinine ratio. These significant variables were adjusted for in subsequent analyses.
Analysis at the allelic level revealed that the rs4636297-A allele was significantly associated with STDR (odds ratio (OR) = 2.02, 95% confidence interval (CI) = 1.22–3.35, p = 0.006) after multivariable adjustment (Table 2). A similar association was identified under an additive (OR = 2.00, 95% CI = 1.19–3.75, p = 0.009) and dominant genotypic model (OR = 2.44, 95% CI = 1.19–5.00, p = 0.015) (Table 3).
Allelic associations of the rs4636297 SNP with sight threatening diabetic retinopathy.
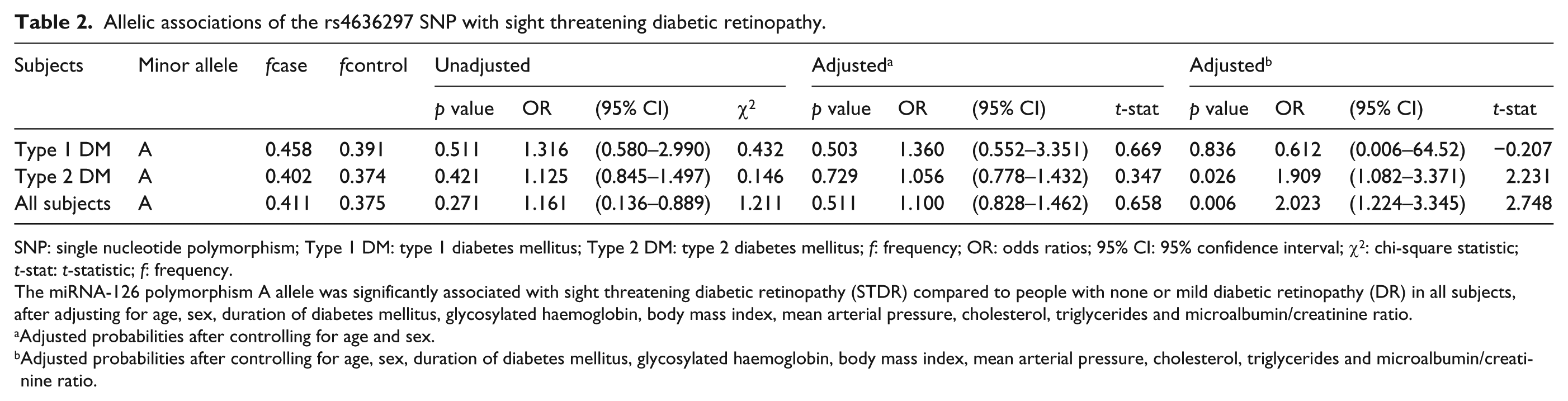
SNP: single nucleotide polymorphism; Type 1 DM: type 1 diabetes mellitus; Type 2 DM: type 2 diabetes mellitus; f: frequency; OR: odds ratios; 95% CI: 95% confidence interval; χ2: chi-square statistic; t-stat: t-statistic; f: frequency.
The miRNA-126 polymorphism A allele was significantly associated with sight threatening diabetic retinopathy (STDR) compared to people with none or mild diabetic retinopathy (DR) in all subjects, after adjusting for age, sex, duration of diabetes mellitus, glycosylated haemoglobin, body mass index, mean arterial pressure, cholesterol, triglycerides and microalbumin/creatinine ratio.
Adjusted probabilities after controlling for age and sex.
Adjusted probabilities after controlling for age, sex, duration of diabetes mellitus, glycosylated haemoglobin, body mass index, mean arterial pressure, cholesterol, triglycerides and microalbumin/creatinine ratio.
Genotypic associations of the rs4636297 SNP with sight threatening diabetic retinopathy.
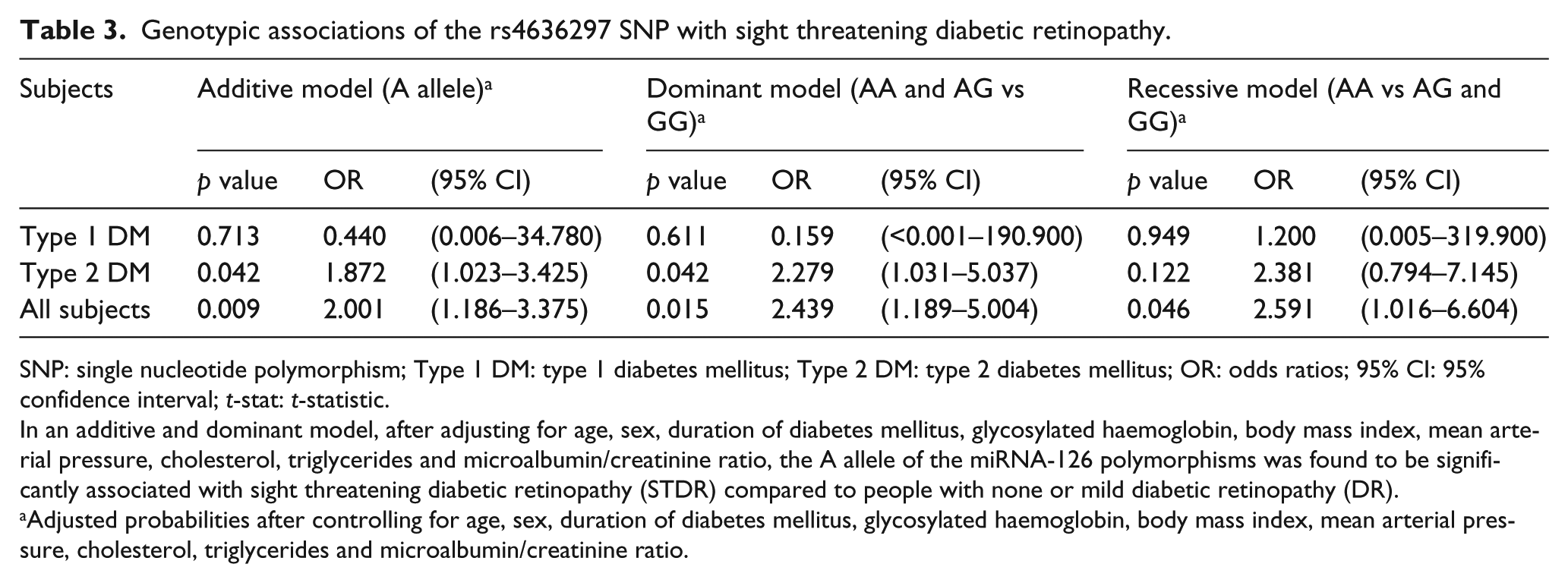
SNP: single nucleotide polymorphism; Type 1 DM: type 1 diabetes mellitus; Type 2 DM: type 2 diabetes mellitus; OR: odds ratios; 95% CI: 95% confidence interval; t-stat: t-statistic.
In an additive and dominant model, after adjusting for age, sex, duration of diabetes mellitus, glycosylated haemoglobin, body mass index, mean arterial pressure, cholesterol, triglycerides and microalbumin/creatinine ratio, the A allele of the miRNA-126 polymorphisms was found to be significantly associated with sight threatening diabetic retinopathy (STDR) compared to people with none or mild diabetic retinopathy (DR).
Adjusted probabilities after controlling for age, sex, duration of diabetes mellitus, glycosylated haemoglobin, body mass index, mean arterial pressure, cholesterol, triglycerides and microalbumin/creatinine ratio.
In the subgroup of people with type 2 diabetes, rs4636297-A allele is significantly associated with STDR (OR = 1.91, 95% CI = 1.08–3.37, p = 0.026). Genotypic analysis suggested that this allele was also significantly associated with STDR under an additive (OR = 1.87, 95% CI = 1.02–3.43, p = 0.042) or dominant model (OR = 2.28, 95% CI = 1.03–5.04, p = 0.042). The subgroup analyses in people with type 1 diabetes did not reveal any significant allelic or genotypic association between the rs4636297-A allele and STDR.
Discussion
Our findings have provided insight into a novel genetic regulator of microvascular dysfunction in STDR. We have confirmed our primary hypothesis that the rs4636297-A variant is associated with STDR. Further subgroup analyses revealed a significant association of this SNP with STDR in people with type 2 diabetes alone. The primary role of the non-protein coding regulatory elements at this locus is to inhibit mature miR-126 expression which indirectly regulates the amount of circulating VEGF. 6 It is important to note that the rs4636297-A variant does not inhibit mature miR-126. 6 This variant allows mature miR-126 to be expressed and there is a lack of regulation on circulating VEGF. As miR-126 functions to ‘inhibit the inhibitors’ of VEGF it can be considered as a VEGF promoter. Our bioinformatics analysis suggests that one copy of the rs4636297-A allele will impair the inhibition of mature miR-126 to increase VEGF promotion and increase the risk for developing STDR.
To further explore the nature of rs4636297-A, we examined the structural changes, including bulges and hairpins, observed between the variants of rs4636297 SNP (Figure 1). There was structural allelic variation present between the G and A variants of rs4636297 SNP (Figure 1). This was illustrated by the lack of two ‘bulges’ in the variant A as well as the addition of a hairpin in the nucleotide structure. Zooming into the EGFL7 locus containing the rs4636297 variant revealed an obvious difference in the predicted structure and in free energy between allelic variants. These changes could impact the regulation of DNA transcription in the region of the nucleotide variation. 18
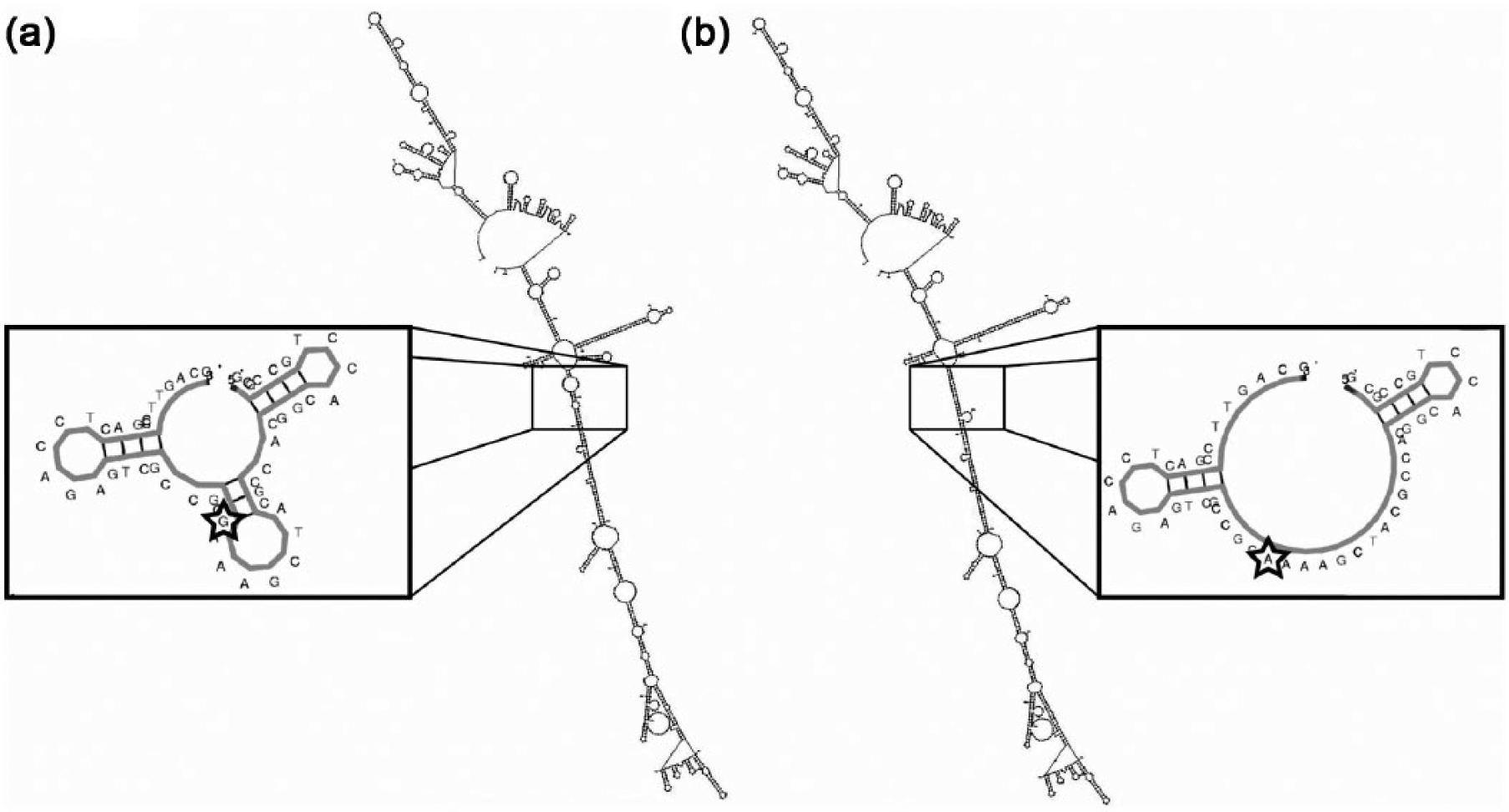
Secondary DNA structure analysis of the G and A allelic variants of rs4636297 in mfold. The image is zoomed into the region with the sequence variation in rs4636297, generated using KineFold. (a) The ancestral rs4636297 allele (G) has an additional hairpin in the nucleotide structure with a free-energy profile (ΔG) of −8.0 kcal/mol; (b) the variant allele (A) lacks two ‘bulges’ in the nucleotide structure, and has a free energy of −6.4 kcal/mol.
The rs4636297 variant is located in the precursory region of miR-126 within the EGFL7 region. 19 EGFL7 is exclusively expressed in endothelial cells and has recently been implicated as a regulator of the notch pathway in the development of neovascularisation, 20 and epigenetic changes in the EGFL7 promoter region are associated with miR-126 expression in tumour growth. 21 As such, the EGFL7 gene region is a particularly encouraging target for future microvascular dysfunction exploration in DR. Future studies could investigate the specific epigenetic regulatory role of the precursory region in miR-126 expression. Understanding epigenetic regulation of miR-126, EGFL7 and its role in VEGF could lead to new gene therapies and increase our understanding of DR pathogenesis.
A limitation of this study was that our sub-analysis in people with type 1 diabetes had limited power due to the smaller sample size. Based on initial analysis, we expect the association with rs4636297 and DR to be consistent across the types of diabetes, and future studies could investigate this further using a larger type 1 diabetes specific cohort. However, our findings have allowed an assessment of DR irrespective of DM, and type 2 diabetes, in a well-defined clinical cohort. Overall, we had sufficient power to detect a variant of modest effect size.
In summary, this association of rs4636297 provides a potentially novel genetic marker for PDR and leads us closer to understanding a pathogenic pathway involved in this blinding condition. Ongoing work investigating the clinical utility and pathogenic importance of this variant in DR is required.
Footnotes
Declaration of conflicting interests
No authors of this article have any competing interests related to this research.
Funding
This research was supported by the Brownless Perpetual Charitable Trust, the Joan and Peter Clemenger Trust, the Diabetes Australia Research Trust, and a National Health and Medical Research Council Centre for Clinical Research Excellence grant – Translational Clinical Research in Major Eye Diseases (#529923). A.K.M. is supported by Australian National Health and Medical Research Council Postgraduate Scholarship (#590231). J.J.W., E.L. and A.W.H. are supported by National Health and Medical Research Council Fellowships (#632909, #1045280 and #1037838, respectively). The Centre for Eye Research Australia receives Operational Infrastructure Support from the Victorian Government.