
Abstract
Purpose:
The purpose of this study was to determine the relative expression of clinically-relevant components of the renin-angiotensin system (RAS) in the adult human eye.
Methods:
We obtained 14 post-mortem enucleated human eyes from patients whom had no history of inflammatory ocular disease nor pre-mortem ocular infection. We determined the gene expression for prorenin, renin, prorenin receptor, angiotensin-converting enzyme, angiotensinogen and angiotensin II Type 1 receptor, on tissue sections and in cultured human primary retinal pigment epithelial and iris pigment epithelial (RPE/IPE) cell lines, using both qualitative and quantitative reverse transcription polymerase chain reaction (RT-PCR). Protein expression was studied using indirect immunofluorescence (IF).
Results:
Almost all components of the classical RAS were found at high levels, at both the transcript and protein level, in the eyes’ uvea and retina; and at lower levels in the cornea, conjunctiva and sclera. There was a much lower level of expression in the reference cultured RPE/IPE cells lines.
Conclusion:
This study describes the distribution of RAS in the normal adult human eye and demonstrates the existence of an independent ocular RAS, with uveal and retinal tissues showing the highest expression of RAS components. These preliminary findings provide scope for examination of additional components of this system in the human eye, as well as possible differential expression under pathological conditions.
Keywords
Introduction
The classical renin angiotensin system (RAS) is demonstrated to have an increasingly important role in ocular pathology and homeostasis. It has been implicated in a variety of eye diseases, including uveitis, macular degeneration, diabetic retinopathy and glaucoma. 1 Classical RAS originates with renin (REN), an aspartyl protease derived from its inactive precursor pro-renin, within the juxtaglomerular cells of the kidney.
REN cleaves its substrate angiotensinogen (AGT), an α2 globulin produced by the liver, in order to form the decapeptide angiotensin I (AngI). Subsequent cleavage of Ang I by the angiotensin-converting enzyme (ACE), results in formation of angiotensin II (AngII), an important vasoconstrictor, trophic, angiogenic and pro-inflammatory octapeptide (Figure 1). ACE also metabolises bradykinin, a powerful vasodilator involved in mediation of the inflammatory response. The RAS was initially considered to be a classical endocrine system with activity restricted to systemic effects; however, recent studies demonstrate localised RAS within numerous tissues, such as the endothelium of the vascular system, brain, adrenal glands, thymus and reproductive tract. In addition, there is mounting molecular, biological and immunohistochemical evidence for a localised RAS in the human eye.2–7 Subsequent work in this area established not only the localised presence of all the constituents of the RAS, but also their expression and active transcription within human ocular tissue.3, 5, 8–10 Published data to date points to the existence of a local RAS within the eye, independent of systemic precursor substrates and peptides. 11

The classical RAS in the systemic circulation.
It has been proposed that the intra-retinal RAS may have a physiological role in the neurotransmission and modulation of retinal ganglion cells. 12 AngII appears to have a multifaceted role in the physiology of the eye; and studies demonstrate its influence on the vasculature of the eye,13,14 aqueous humour outflow regulation and intraocular pressure (IOP) control.2,15 In addition, its presence in certain types of neural cells in the retina, such as Müller glial cells, has implicated AngII in ocular neuromodulation of the pathogenesis of vasoproliferative disorders, such as proliferative retinopathy,3,5,8,16,17 stimulating VEGF-induced angiogenesis within the retina18–22 in disease states such as retinopathy of prematurity (ROP)23–25 and diabetic retinopathy. 24 Plasma AngII is also believed to play an important role in mediating the influence of systemic RAS on intraocular RAS. 26
RAS is also implicated in other disease processes in the eye, including inflammatory disorders.27–30 AngII is shown to enhance vascular permeability in an inflammatory setting, predominantly through the induction of chemokines and adhesion molecules, as well as being responsible for recruitment of inflammatory cells. 31 Inhibition of the RAS was also demonstrated to have a neuroprotective effect in glaucoma models, such as for ocular hypertension, 32 reperfusion 33 and deaxotomised retina explants. 34
The aim of our study was to provide a comprehensive picture of the RAS gene and protein expression patterns in normal adult human ocular tissue, which will aid in targeting treatments for a variety of RAS-mediated eye diseases.
Materials and methods
Tissue specimens
We obtained 10 fresh and 5 fixed whole human eyes, post-mortem, from the Lions NSW Eye Bank in Sydney, Australia. Informed consent was obtained. The experimental protocols followed the tenets of the Declaration of Helsinki and the study was approved by the Human Research Ethics Committee of the University of NSW.
The cornea and conjunctiva of the 10 fresh eye specimens had been removed for use in corneal graft surgery; therefore, these tissues were unavailable for RT-PCR testing. Exclusion criteria for all specimens were having inflammatory ocular disease and pre-mortem ocular infection. Other ocular comorbidities are noted in Tables 1 and 2. We did not have detailed information on systemic comorbidities of the donors, beyond what was relevant for transplant safety. Tables 1 and 2 highlight the clinical features of the 15 cases.
Clinical features of normal eyes used for RT-PCR experiments.

F: female; L: left; hrs: hours; M: male; N/A: not applicable; PMI: post-mortem interval; R: right; RT-PCR: reverse transcriptase polymerase chain reaction; SD: standard deviation
Clinical features of eyes used for immunohistochemistry experiments.
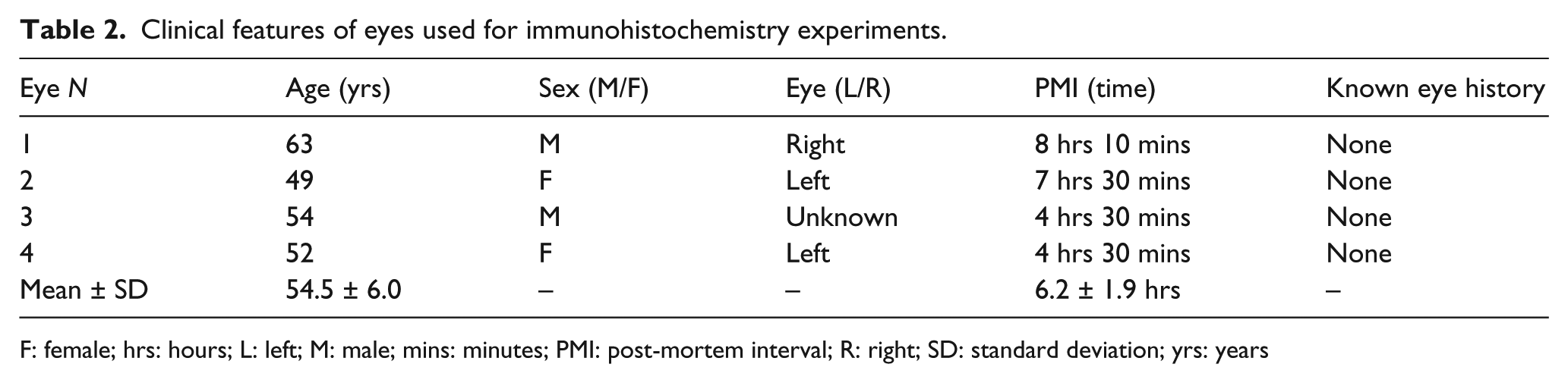
F: female; hrs: hours; L: left; M: male; mins: minutes; PMI: post-mortem interval; R: right; SD: standard deviation; yrs: years
Tissue dissection
Enucleation of the globe was performed post-mortem by an ophthalmic surgeon. The surgeon cut a further circular incision around the eyeball, slightly anterior to the insertion of the lateral rectus muscle, to remove the cornea and bulbar conjunctiva for corneal transplant/graft purposes. Upon enucleation and removal of the cornea and conjunctiva for use by the Eye Bank, further tissue dissection was carried out under an optical microscope, to minimise any tissue-to-tissue contamination. The iris, ciliary body, neural retina, RPE and attached choroid were dissected from the globe, sliced into fine pieces and placed into individual eppendorf tubes for ribonucleic acid (RNA) extraction. The entire ciliary body and iris (approximately 50 – 100 mg each) were small enough to only need one tube each; however, the whole retina and choroid (approximately 200 mg each) were split across three tubes each, due to the larger amount of these tissues in the eye. A sample of approximately 100 mg in weight of sclera was also used.
Cell cultures
We purchased human primary IPE and RPE cells with epithelial cell medium from ScienCell Research Laboratories (San Diego, CA) and cultured them as previously described. 35 Two control cell lines, THP1 (a human acute monocytic leukaemia cell line) and SKBR3 (a breast cancer cell line) were donated in the form of freshly extracted RNA by the ocular immunology group at the University of NSW, Kensington.
RT-PCR methods
Total RNA was extracted from both the post-mortem tissues and cell lines, and DNase treated. We isolated RNA, performing reverse transcription and PCR with SYBR green dye. A total of 50 positive (RT +) and 50 negative (RT –) samples were available for RT-PCR, from five ocular tissue samples from each of the 10 donor eyes. We constructed positive control standard curves for relative quantification. We used the standard curve method for relative quantification, to calculate unknown sample expression as a fold expression over the endogenous control gene (GAPDH).
We performed PCR for the AGTR1 Transcript 5 (AT1 receptor) gene on synthesised cDNA, in accordance with the Quantace SensiMix protocol (Bioline, Sydney, Australia). As the primer set used produced an amplicon too large for RT-PCR, we used qualitative PCR for the AGTR1 transcript variant 5 gene, instead of quantitative RT-PCR. We ran the amplified products from the AGTR1 PCR in a 10% polyacrylamide gel electrophoresis (PAGE).
Immunohistochemical analysis
Whole eyes (n = 4) obtained from the Lions Eye Bank were fixed in 10% neutral-buffered formalin for 3 – 5 days and then processed overnight (VIP Tissue Processor, Olympus, Notting Hill, VIC, Australia). The whole eyes were then paraffin embedded and sectioned.
IPE and RPE cells at passage 4’ were cultured in epithelial cell medium to confluence in chamber slides. The cells were washed in phosphate buffered saline (PBS), then fixed in 25% v/v methanol, 25% v/v acetone and 50% v/v reverse osmosis water for 10 minutes, followed by Tris buffered saline (TBS) washes prior to the immunostaining procedure detailed below.
Immunofluorescence staining was performed on 5µm thick paraffin sections, mounted on coated slides (Vision BioSystems, Melbourne, Australia; Leica MicroSystems, Sydney, Australia), and allowed them to dry overnight in an oven set at 37°C. Paraffin slides from each eye were deparaffinised and rehydrated. Antigen retrieval was performed in a microwave, using Epitope Retrieval Solution pH 6.0 (Novacastra, Newcastle upon Tyne, UK) for 10 minutes (3 mins at 100% power and 4 mins at 50% power). All the sections were then incubated in 2% skim milk powder in TRIS buffer, to minimise non-specific binding. All tissue sections were then incubated with the primary antibodies (list in Table 3) overnight, at 4°C. Sections were washed in buffer (Novacastra, Newcastle upon Tyne, UK) and followed by incubation with secondary antibodies that included: goat anti-mouse Alexa Fluor 546 (Invitrogen, CA, USA), goat anti-rabbit Alexa Fluor 594 (Invitrogen, CA, USA), goat anti-sheep Alexa Fluor 594 (Invitrogen, CA, USA) and donkey anti-goat Alexa Fluor 594 (Invitrogen, CA, USA). Sections were treated with 0.03% Sudan black for 2 mins and washing in running water, before mounting with Prolong Gold antifade media.
Antibodies used in immunostaining experiments.

Images were taken using the Olympus Fluoview program and original slides were imaged using a confocal microscope (Olympus FV1000 Inverted Laser Scanning Microscope).
Results
Gene expression
Qualitative and quantitative RT-PCR
Qualitative PCR showed the AGTR1-transcript for the AT1 receptor to be present in RPE choroid samples from donor eyes only (Figure 2). Quantitative PCR was used to determine the relative expression of ACE, AGT, REN/prorenin and prorenin receptor (ATP6AP2) genes. Only ocular samples which expressed levels of GAPDH (the control gene) above the detectable limit of the assay in three or more samples were quantified. Relative gene expression as a fold difference in reference to the calibrator samples was calculated. The relative expression of these genes is shown in Figure 3.

Qualitative expression of AT1R by RT PCR. The lanes are numbered as follows: (a) 0

Expression of mRNA by RT-PCR of components of the RAS in iris, ciliary body, retina, choroid and sclera. Expression was calculated as a relative increase in SYBR green intensity, relative to a control gene (GADPH). Calculations were based on a minimum of three replications.
Patterns and locations of protein immunoreactivity
Protein expression and localisation of RAS components (AGT, prorenin, prorenin receptor, ACE and AT1 receptor) within ocular tissue was determined using indirect immunofluorescence. Figure 4 is representative of the staining obtained for each of the RAS components tested in the eye. In this case, we show staining for the AT1 receptor across the eye.
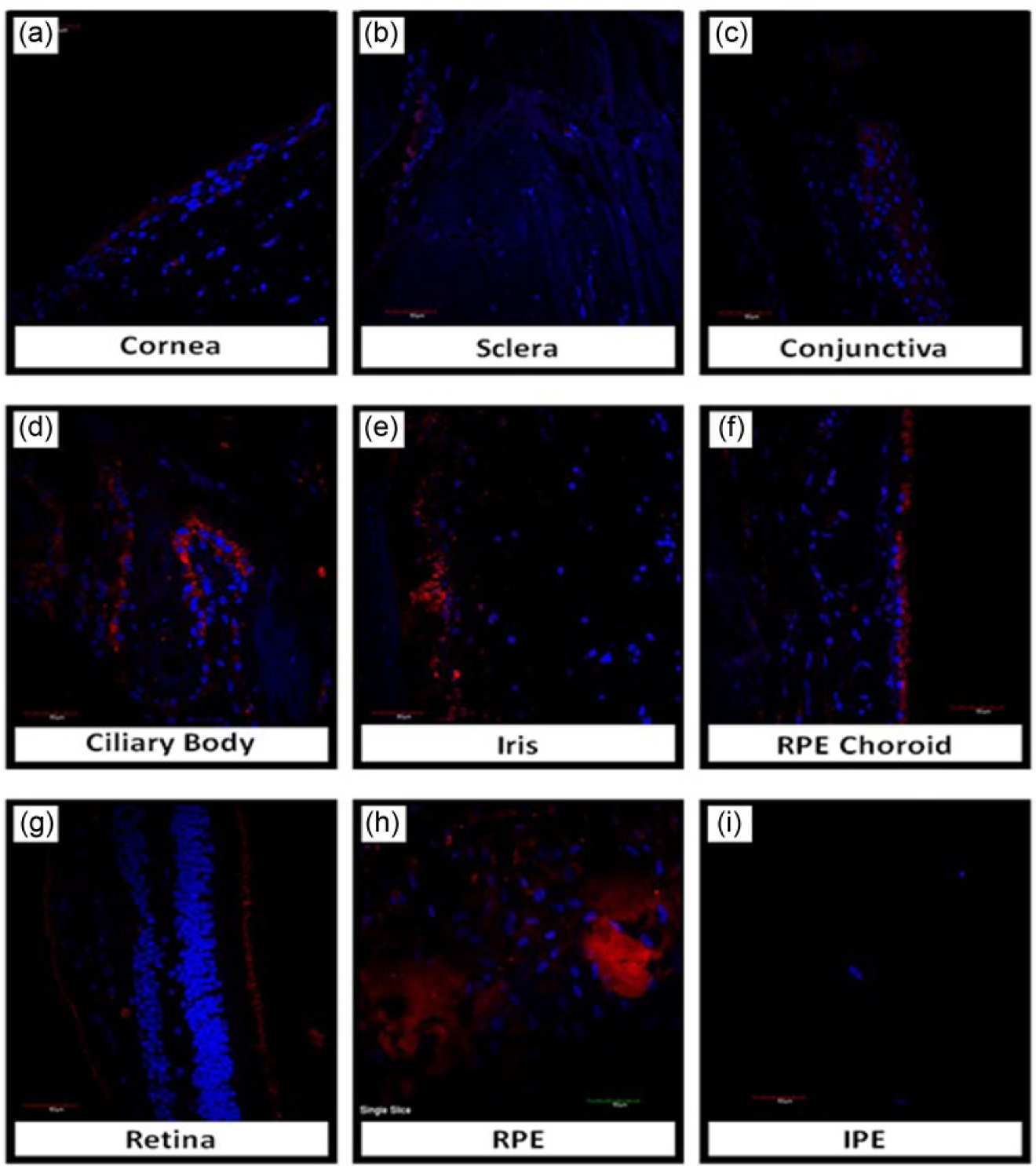
Immunostaining with the angiotensin II Type I receptor antibody (red) across various samples of eye tissue, as labelled. Counterstain of cell nuclei is DAPI (blue). Immunoreactivity patterns are as follows: (a) Corneal epithelium; (b) None in sclera; (c) conjunctival epithelium; (d) ciliary body pigmented epithelium; (e) iris pigment epithelium; (f) retinal pigment epithelium and choroidal vasculature; (g) layer of rods and cones and inner limiting membrane in the retina; (h) RPE cultured cells; (i) IPE cultured cells.
The staining was assessed semi-quantitatively by three independent observers (other staff in the lab unconnected to the project) looking at the images with their naked eyes, on the computer screen. A score of ‘0’ was given for no staining visible, ‘1’ for some staining and ‘2’ for intense staining. The benchmark to decide what constituted no staining or maximum staining was based on control (non-eye) tissues, which had no discernible staining or diffuse, intense staining, respectively. We chose the control tissues based on tissues where that protein was known to exist, such as REN in the kidney. The scores for each individual slide were then averaged to get a total mean score between 0 – 2 for each slide. Figure 5 shows how intensity scoring was derived in the control samples.
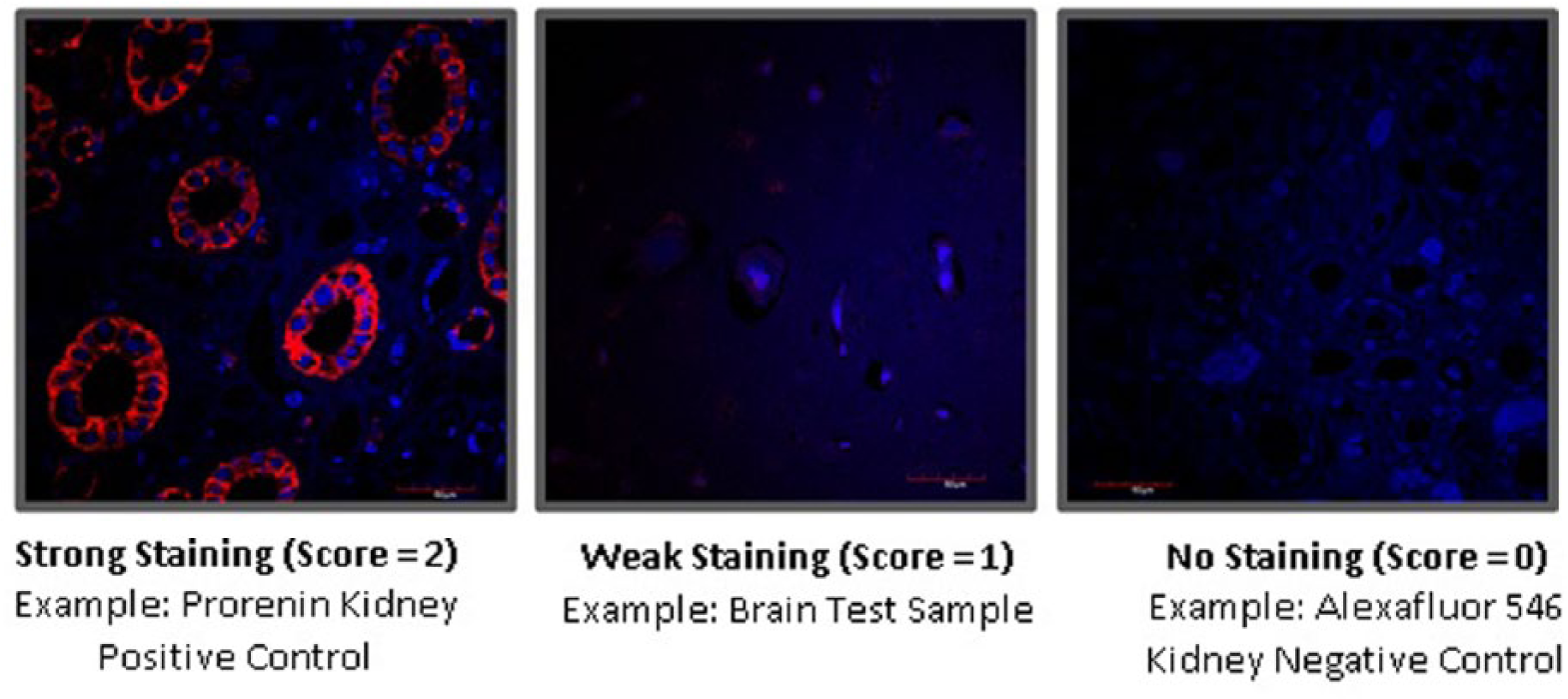
An example of staining of control tissue, illustrating the scoring system used to quantify immunofluorescence staining. Immunostaining is red, counterstained with DAPI (blue).
Table 4 summarises the average staining intensity of the various RAS components in different structures of the eye.
Summary of RAS protein expression a using indirect immunofluorescence staining pattern.

Key: +++ (strong) = 1.5 to 2; ++ (moderate) = 0.5 to < 1.5; + (weak) = 0 to < 0.5
ACE: angiotensin-converting enzyme; AGT: angiotensinogen; AT1: angiotensin 1; CB: ciliary body; IPE: iris pigment epithelial; RAS: renin-angiotensin system; RPE: retinal pigment epithelial
Discussion
This is the first study to catalogue the expression patterns of prorenin, REN, the prorenin receptor, angiotensinogen, ACE and the AT1 receptor in the human eye, at both the transcript and protein levels. Previous studies on RAS in the human eye looked at mRNA expression only, and not across all structures of the eye. 3 The presence and active transcription of genes encoding RAS proteins, within different types of eye tissue, are further evidence of a localised RAS in the eye.
Importantly, this study demonstrated a strong expression of RAS mRNA and protein in uveal and retinal tissues, relative to the corneoscleral layer. This finding has important implications in uveo-retinal disease, as these tissues are more likely to produce angiotensin in large amounts. The proinflammatory, profibrotic, mitogenic and vasoconstrictor properties of angiotensin II, which contribute to vascular and inflammatory disease by altering vascular tone, structural remodelling and endothelial dysfunction36–38 are likely to be the same mechanisms that promote many RAS-related diseases of the eye.
Almost all layers of the retina were positive for RAS proteins, which correlates with previous findings of a neuromodulatory or possible neurotransmitter role for angiotensin II in the retina.8,16 The modulation of reactive oxygen species (ROS) generation via an AT1R-mediated mechanism, demonstrated in in vitro glaucoma models to confer a neuroprotective effect, lends weight to this hypothesis.33,34 ROS ‘bursts’ are implicated as a trigger mechanism for apoptotic cascades in the eye and optic nerve.34,39
Human RPE and IPE cell cultures showed either low or no expression of the RAS constituents at either transcript or protein levels. Contributing factors could include differences between functionality in the in vitro and in vivo setting, the conditions under which the cells were cultured and the presence of other cell types within the epithelial layer being the source of RAS proteins.
There are important limitations to this study. There were significant variations in expression of RAS components between donors. The medication histories of our donors were unavailable; and therefore, the effects of medications that could have influenced the results of this study are unknown. The impact of anti-RAS medications, such as ACE inhibitors and angiotensin receptor antagonists, on ocular tissues may explain significant differences in protein expression. RNA degradation in the RPE and other ocular tissues, due to the time between donor death and tissue processing (mean time: 11.9 h) may have also been a factor. Furthermore, only a total of 15 human eyes were available for this exploratory study. A much larger sample size would overcome many of these difficulties, as well as allow a direct comparison between RAS expression levels in healthy and diseased eyes.
Footnotes
Acknowledgements
We are very grateful for the very helpful comments on the manuscript from Olaf Strauss, Charité Universitätsmedizin, Berlin.
Conflict of interest
None declared.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.