
Abstract
Background and objective:
Incidences of hypertension are increasing and this condition is more common in men than in women. We selected six well-characterized polymorphisms from three X-linked genes (ACE2, AGTR2, apelin) aiming to investigate their interactive association with hypertension among northeastern Han Chinese.
Methods and results:
This was a case-control study involving 1009 hypertensive patients and 756 normotensive controls. All polymorphisms except rs3761581 in the apelin gene satisfied the Hardy-Weinberg equilibrium in females. The genotype and allele distributions of rs1403543 in the AGTR2 gene and rs56204867 in the apelin gene differed significantly between patients and controls for both genders, even after the Bonferroni correction (P<0.05/6). The risk prediction was significant for rs1403543 and rs56204867 under both additive and dominant models for both genders. In haplotype analysis, significance was seen for haplotype G-T-G-G-A (alleles in order of rs1978124, rs2106809, rs1403543, rs5194 and rs56204867), which was overrepresented in patients (5.15% versus 1.10% in controls, PSim=0.004). Interaction analysis indicated that all derived multifactor dimensionality reduction (MDR) models were non-significant for both genders.
Conclusion:
Our findings demonstrate that genetic defects in AGTR2 and apelin genes by themselves may play an independent leading role in determining susceptibility to hypertension in both genders.
Introduction
Hypertension is a multifactorial polygenic disease. Twin and family studies estimate that the genetic contribution to blood pressure variation is approximately 30–60%.1,2 Incidences of hypertension are increasing and it is more common in men than in women. 3 Substantial evidence supports the notion that genetic defects on sex chromosomes may be associated with gender-specific susceptibility to hypertension. For example, our previous study on the angiotensin-converting enzyme 2 (ACE2) gene found an intronic polymorphism called rs1978124 was significantly associated with stage 2 hypertension in men only. 4 There are a cluster of genes on the X chromosome, and to determine how many genes and which genetic determinants are actually involved in the pathogenesis of hypertension remains an interpretative challenge.
Generally, a single gene or polymorphism explains only a small proportion of individual variation in disease susceptibility, and it is therefore important to assess the joint effect of multiple genes from a common pathologic pathway. 5 With this in mind, in our study we selected six well-characterized polymorphisms from three X-linked genes, two in the renin-angiotensin system (Xp22: ACE2; Xq22-q23: angiotensin II type 2 receptor (AGTR2)) and one in apelin/angiotensin receptor-like 1 (apelin/AGTRL1) system (Xq25: apelin) to investigate their interactive association with the risk of having hypertension in a large northeastern Han Chinese population. There is a close relationship between the renin-angiotensin system and the apelin/AGTRL1 system, as evidenced from both human and animal studies, which has implicated the involvement of both systems in regulating body fluid homeostasis and cardiovascular functions.6–10 It also is worth noting that the apelin/AGTRL1 system is believed to play an important role in counter-regulating the effect of the renin-angiotensin system.11–13 Therefore, it is reasonable to speculate that the three X-linked genes selected from the two systems might be logical candidates for studying the genetic underpinnings of hypertension.
Methods
Study population
This was a hospital-based case-control study as we previously reported.14–16 All study participants were recruited from Qiqihar city, Heilongjiang, China. The Institutional Review Board of Qiqihar Medical University approved this study and written informed consent was obtained from each participant at enrollment. This study was performed in accordance with the guidelines of the Declaration of Helsinki.
Blood pressure was measured by a calibrated mercury sphygmomanometer with an appropriate adult cuff size by certified examiners. Hypertension was defined as a mean systolic blood pressure of more than 140 mmHg or a diastolic blood pressure of more than 90 mmHg or the current use of antihypertensive drugs. As recommended by Tobin et al., 17 for participants under antihypertensive medication, blood pressure was determined by adding 15 and 10 mmHg to the systolic and diastolic blood pressure, respectively.
All study participants were classified into the hypertensive group and the normotensive group according to the clinical and laboratory examinations. The hypertensive group consisted of inpatients or outpatients of the Second Affiliated Hospital of Qiqihar Medical University. Hypertensive patients with clinical evidence of secondary hypertension and renal diseases were excluded. Normotensive controls who underwent medical examinations at the same hospital were clinically confirmed to be free of hypertension and had a negative family history of hypertension in their first-degree relatives. All study participants were genetically unrelated local residents of Han Chinese descent who were consecutively recruited between June 2008 and December 2012.
Sample size
The hypertensive group had 1009 sporadic patients aged 64.48 (standard deviation: 8.53) years and 54.31% of the patients were men. The rest of the participants (n=756), who had normal blood pressure, formed the age-, gender- and ethnicity-matched control group.
Demographic and clinical measurement
At enrollment, age, gender, body weight and height were recorded. Body mass index (BMI) was calculated as weight in kilograms divided by height in meters squared.
Fasting venous blood samples were obtained from all participants for DNA extraction and biochemical marker detection. Plasma triglyceride, total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), blood urea nitrogen (BUN), serum creatinine and uric acid levels were determined enzymatically using available kits and auto analyzers. High-sensitivity C-reactive protein (hsCRP) level was measured using a particle-enhanced immunoturbidimetric method.
Selection of polymorphisms
Six polymorphisms were evenly selected from three X-linked genes, that is, rs1978124 (A1075G, intron 1) and rs2106809 (intron 1) from the ACE2 gene, rs1403543 (G1675A, intron 1) and rs5194 (G2274A, 3’ UTR) from the AGTR2 gene and rs56204867 (T-1860C, promoter) and rs3761581 (A/C, promoter) from the apelin gene. These six polymorphisms have been well characterized and showed potential associations with hypertension on the basis of a comprehensive overview of published studies.4,18–25
Genotype determination
Genomic DNA was extracted from peripheral blood leukocytes according to a standard phenol-chloroform method and stored at -40°C until required for batch genotyping. The polymerase chain reaction − ligase detection reaction (PCR-LDR) method was used to determine the genotypes of six examined polymorphisms according to a previous report. 26 For each polymorphism, two specific probes were synthesized to discriminate specific bases, and in addition one common probe was synthesized and labeled at the 3’ end with 6-carboxy-fluorescein (FAM) and phosphorylated at the 5’ end. The fluorescent products of LDR were differentiated using a ABI 3730XL sequencer (Applied Biosystems, California, USA).
To test the accuracy of this PCR-LDR method, 96 DNA samples were randomly selected and run in duplicates for all six polymorphisms with 100% concordance.
Statistical analysis
Continuous and categorical variables were compared between hypertensive patients and normotensive controls by unpaired t-test, Mann-Whitney U test and χ2 test when appropriate. The Hardy-Weinberg equilibrium was tested by Pearson goodness-of-fit test. The genotype and allele distributions between the two groups were compared by χ2 test. The statistical analyses described above were completed with the STATA software (version 11.0) for Windows (StataCorp LP, College Station, TX, USA). Study power was estimated by the Power and Sample Size Calculations (PS) software (version 3.0.7).
Because six examined polymorphisms were located on the X chromosome, haplotypes were constructed and their frequencies were estimated by the haplo.em program. To avoid chance results, only a haplotype with frequency ≥1% in all participants was analyzed. P values were simulated based on 1000 replicates. The odds ratio (OR) and its 95% confidence interval (95% CI) were used as a measure of strength for the association between different derived haplotypes and hypertension risk before and after adjusting for age and BMI. Moreover, the haplo.score program is used to model an individual’s phenotype as a function of each inferred haplotype, weighted by their estimated probability, to account for haplotype ambiguity. The haplo.em and haplo.score programs are implemented in the Haplo.Stats software (version 1.4.0) operated in the R language (version 2.14, available at http://www.r-project.org).
To further detect and characterize the interaction of the three X-linked genes, an open-source data-mining multifactor dimensionality reduction (MDR) approach (version 3.0, available at http://www.epistasis.org) was adopted.27,28 The aim of this approach is to construct all possible combinations of examined polymorphisms to identify the overall best model. The accuracy of each MDR model is evaluated by a Bayes classifier in the context of 10-fold cross-validation. A single best model has the maximal testing accuracy and cross-validation consistency simultaneously. Statistical significance was evaluated using a 1000-fold permutation test to compare observed testing accuracies with those expected under the null hypothesis of null association. Permutation testing corrects for multiple testing by repeating the entire analyses on 1000 datasets that are consistent with the null hypothesis.
Results
Baseline characteristics of the study population
Table 1 shows the baseline characteristics of the study population in both males and females. For both genders, there was no significant difference in age between hypertensive patients and normotensive controls. The levels of BMI, adjusted systolic and diastolic blood pressure, fasting glucose and hsCRP were significantly higher, but the HDL-C level was significantly lower in patients than in controls. Relative to normotensive controls, hypertensive patients had higher TG levels for females (P=0.014) and higher creatinine level for males (P=0.002). A percentage of 16.05% of females and 12.59% of males received antihypertensive treatment.
The baseline characteristics of all study participants.

BMI: body mass index; SBP: systolic blood pressure; DBP: diastolic blood pressure; TC: total cholesterol; TG: triglyceride; HDL-C: high-density lipoprotein cholesterol; BUN: blood urea nitrogen; hsCRP: high sensitivity C-reactive protein.
P values were computed by unpaired t-test or Mann-Whitney U test for quantitative variables and by χ2 test for qualitative variables.
Single-locus analysis
The genotype distributions and allele frequencies of the six examined polymorphisms are presented in Table 2. Because males have only one X chromosome, genotype information is not available. In females, all polymorphisms but rs3761581 (P<0.01) satisfied the Hardy-Weinberg equilibrium at a significance level of 5%. The genotype and allele distributions of rs1403543 and rs56204867 differed significantly between patients and controls for both genders, even after the Bonferroni correction (Bonferroni significance threshold P=0.05 divided by the number of polymorphisms under study (n=6): P=0.0083). The power to reject the null hypothesis of no differences in the mutant allele frequencies of rs1403543 and rs56204867 between the two groups was 99.6% and 94.3% in females, and 77.7% and 82.9% in males, respectively. In addition, there was marginal significance in allele distributions of rs2106809 for both genders, while this significance disappeared after the Bonferroni correction. No observable difference was identified for the other polymorphisms.
Genotype distributions and allele frequencies of six examined polymorphisms in three X-linked genes between hypertensive patients and normotensive controls according to gender.

W/M, wild allele/mutant allele.
P values were computed by using χ2 test based on the 3×2 contingency tables for genotype comparisons and on the 2×2 contingency tables for allele comparisons.
For females, the genotypes of all examined polymorphisms were investigated under additive, dominant and recessive models of inheritance; however, since there is only one X chromosome in males, only the allelic model was calculated (Table 3). For females, after the Bonferroni correction (Bonferroni significance threshold P=0.05 divided by the product of number of polymorphisms under study and the number of genetic models (n=6×3): P=0.0028) and adjusting for confounding factors including age and BMI, there was a statistical significance for rs1403543 and rs56204867 under both additive and dominant models. For example, the adjusted odds of having hypertension was 1.8 (95% CI: 1.4–2.31; P<0.001) for rs1403543 and 1.48 (95% CI: 1.19–1.85; P=0.001) for rs56204867 under additive model. For males, with a Bonferroni corrected alpha of 0.0083 (0.05/6), significance was still seen for rs1403543 (OR=1.72; 95% CI: 1.15–2.58; P=0.008) and rs56204867 (OR=1.51; 95% CI: 1.14–1.99; P=0.004) under allelic model.
Risk prediction of six examined polymorphisms for hypertension under different genetic models of inheritance by gender.

Data are expressed as odds ratio (OR); 95% confidence interval (95% CI); P value. All genetic models were adjusted for age and body mass index.
Haplotype analysis
In view of the deviation of rs3761581 from the Hardy-Weinberg, haplotypes were constructed based on the other five polymorphisms (Table 4). In females, haplotype G-T-G-A-G (alleles in order of rs1978124, rs2106809, rs1403543, rs5194 and rs56204867) was the most common haplotype, and its frequency was comparable between patients and controls (PSim=0.393). After the Bonferroni correction (Bonferroni significance threshold P=0.05 divided by the number of derived haplotypes (n=12): PSim=0.0042), significance was only seen for haplotype G-T-G-G-A, which was overrepresented in patients (5.15% versus 1.10% in controls, PSim=0.004), and there was 99.7% power to detect this significance. In contrast to males, haplotype G-T-G-A-G was the most common haplotype, and its frequency was slightly higher in controls than in patients (PSim=0.070). There was no observable significance for the other derived haplotypes.
Haplotype frequencies (≥3%) of five polymorphisms between hypertensive patients and normotensive controls by gender.
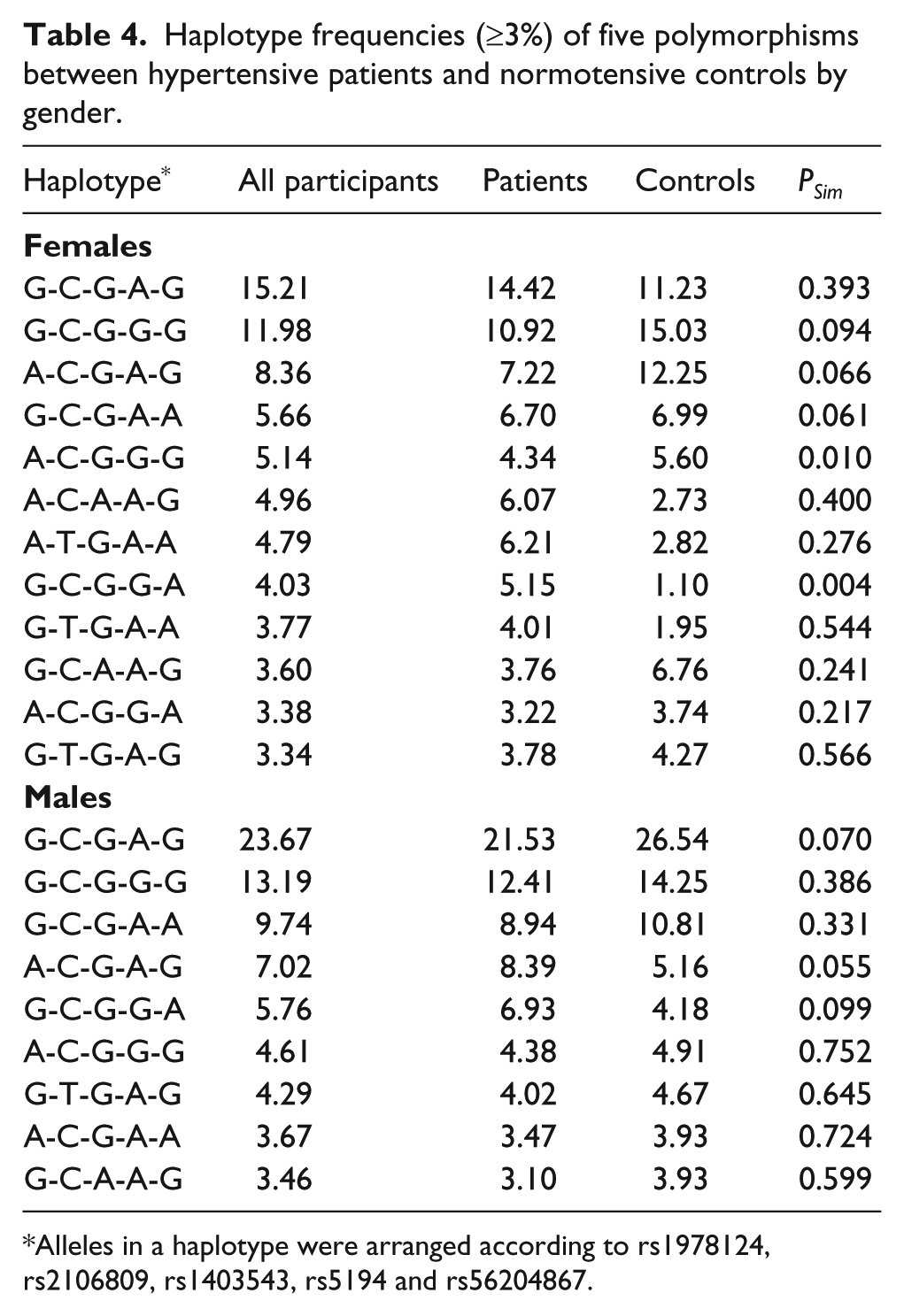
Alleles in a haplotype were arranged according to rs1978124, rs2106809, rs1403543, rs5194 and rs56204867.
Interaction analysis
To yield more information, a promising data-mining analytical approach MDR was adopted to detect and characterize the potential interactions of the five aforementioned polymorphisms in Hardy-Weinberg equilibrium. Each best model is evaluated by the testing accuracy, cross-validation consistency and significance level. As shown in Table 5, all derived MDR models were non-significant for both genders, suggesting the independence of examined polymorphisms in susceptibility to hypertension.
Summary of MDR analysis in both genders.
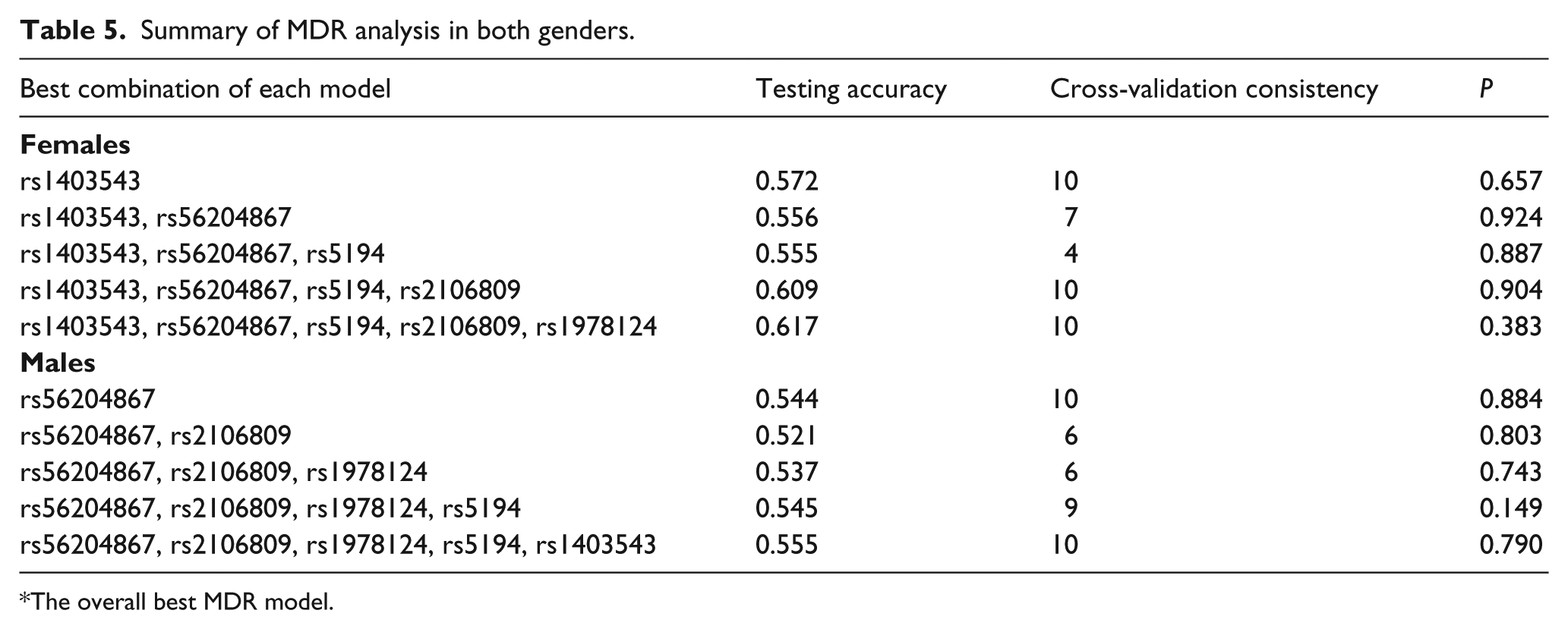
The overall best MDR model.
Discussion
In the present study, we aimed to investigate the interactive association of six well-characterized polymorphisms from three X-linked candidate genes in both renin-angiotensin and apelin/AGTRL1 systems with the risk of having hypertension in a large northeastern Han Chinese population. The key finding of this study was that genetic defects in AGTR2 and apelin genes by themselves might play an independent leading role in determining susceptibility to hypertension in both genders. Although there were no signs of observable interaction, this study, to the authors’ knowledge, is the first report dealing with the association of three X-linked genes, both individually and interactively, with hypertension in a Chinese population.
Several strengths distinguishing the present study merit adequate consideration. First, this study focused on three X-linked genes from the renin-angiotensin and apelin/AGTRL1 systems, two promising pathways leading to hypertension. Our previous findings supported the potential genetic susceptibility of the two systems to hypertension,4,18,19,29–34 highlighting the biological plausibility of selected genes. Second, this study population of northeast China is characterized by genetic homogeneity and geographic stability, and the study participants are most likely similar in their environmental exposures, including the habitual intake of high-salt and high-fat diets and lower rates of hypertension recognition and treatment. All these characteristics make this population more suitable for studying the genetics of hypertension.35,36 Third, all examined polymorphisms but one respected the Hardy-Weinberg equilibrium, lowering the likelihood of being biased by faulty genotyping or population stratification. Fourth, this study had more than 94% power to detect statistical significance in females, and the average power in males reached almost 80%. Fifth, data from 1765 study participants were both statistically corrected and adjusted for traditional confounders, reinforcing the soundness of our findings. Nonetheless, despite the possible existence of residual confounding from incompletely measured or unmeasured physiologic covariates, it seems unlikely that our results could be explained by chance.
Considering the ubiquitous nature of epistasis in genetic architecture of common human diseases, 37 it is a high priority to examine the interaction of multiple genes in a common pathologic pathway. There are many data-mining techniques to identify high-order gene-gene and gene-environment interactions, and among them the MDR approach is the most widely used in recent years. This approach is nonparametric and model-free in design and has been successfully applied to detect and characterize high-order gene-gene and gene-environment interactions in studies with relatively small samples.31,38 By using this approach, we constructed all possible overall best models, and unfortunately our present data failed to observe any signs of potential interaction between the three X-linked candidate genes. With respect to the null interaction observed in this study, we think epistasis is unlikely. Instead, findings from single-locus analyses supported the leading role of two polymorphisms, rs1403543 and rs56204867, respectively from AGTR2 and apelin genes in the development of hypertension, and this role is independent of confounding factors.
Recently, we have validated the candidacy of rs56204867 (T-1860C) in the apelin gene for hypertension in both genders among southern Han Chinese, 18 and given that this polymorphism is located in the promoter region, it is expected that if involved, this polymorphism might, by itself or in linkage with another functional locus, be responsible for the transcriptional regulation of the apelin gene. By contrast, we in this study failed to observe any significant association for another promoter polymorphism rs3761581 in the apelin gene, suggesting the obvious regional differences in socio-economic development levels and the dietary habits between northern and southern Chinese. Taking salt consumption as an example, it is well known that a high-salt diet can induce or aggravate hypertension in both animal models and humans.39,40 However, as documented by a national multi-center report, the average urinary sodium excretion in northern Chinese (271 mmol/d) nearly doubled the amount of that in southern Chinese (139 mmol/d), leading to an average 7.4 and 6.9 mmHg increase in systolic and diastolic blood pressure, respectively. 41 We therefore propose that genetic determinants may predispose individuals to develop a disease; however, its incidence is largely dependent on the exposure to a certain environment or lifestyle. Moreover, given the fact that the evolutionary history of linkage disequilibrium patterns will vary significantly in different ethnic populations, 42 there is a need to construct a database of hypertension-susceptibility genes or polymorphisms in each ethnic group.
Interpretation of the present study, however, should consider several limitations. First, the retrospective design of this study has inherent drawbacks and might have introduced survival-related bias. 43 Second, this study of 1765 participants may be inadequate to demonstrate small risk effects, but our findings need to be validated in another independent population of China and other ethnicities. Third, only six common polymorphisms were examined on the X-chromosome, and it is highly encouraged to incorporate other polymorphisms, especially the low-penetrance ones. Fourth, data on the types of antihypertensive drugs and the treatment period were not available for us, which precluded further analyses of drug effects on our estimates. Fifth, the fact that our study participants were of northeastern Han Chinese descent limited the generalizability of our findings to other ethnic groups.
Taken together, our findings demonstrate that genetic defects in AGTR2 and apelin genes by themselves might play an independent leading role in determining susceptibility to hypertension in both genders. As hypertension is a multifactorial complex condition, well-designed longitudinal studies attempting to assess gene-gene and gene-environment interactions, as well as studies seeking to provide biological implications, are warranted in the future.
Footnotes
Conflicts of interest
None declared.
Funding
This study received grants from the Natural Science Foundation of Xinjiang (2012211A075) and the Shanghai Rising Star Program (11QA1405500).