
Abstract
Pharmacologic inhibition of nitric oxide production inhibits growth of coronary collateral vessels. Dimethylarginine dimethylaminohydrolase 1 (DDAH1) is the major enzyme that degrades asymmetric dimethylarginine (ADMA), a potent inhibitor of nitric oxide synthase. Here we examined regulation of the ADMA-DDAH1 pathway in a canine model of recurrent myocardial ischemia during the time when coronary collateral growth is known to occur. Under basal conditions, DDAH1 expression was non-uniform across the left ventricular (LV) wall, with expression strongest in the subepicardium. In response to ischemia, DDAH1 expression was up-regulated in the midmyocardium of the ischemic zone, and this was associated with a significant reduction in myocardial interstitial fluid (MIF) ADMA. The decrease in MIF ADMA during ischemia was likely due to increased DDAH1 because myocardial protein arginine N-methyl transferase 1 (PRMT1) and the methylated arginine protein content (the source of ADMA) were unchanged or increased, respectively, at this time. The inflammatory mediators interleukin (IL-1β) and tumor necrosis factor (TNF-α) were also elevated in the midmyocardium where DDAH1 expression was increased. Both of these factors significantly up-regulated DDAH1 expression in cultured human coronary artery endothelial cells. Taken together, these results suggest that inflammatory factors expressed in response to myocardial ischemia contributed to up-regulation of DDAH1, which was responsible for the decrease in MIF ADMA.
Keywords
Introduction
Coronary angiogenesis and collateral vessel growth are chronic adaptations to myocardial ischemia that can restore blood flow and salvage myocardium in ischemic regions. Growth of the small pre-existing interarterial anastomotic channels to form effective collateral vessels (arteriogenesis) involves the addition of new vascular components (endothelium, smooth muscle and fibroblasts) via mitosis, migration, tissue remodeling, matrix degradation and differentiation. 1 Consequently, regulation of translation, transcription and protein modifications are expected to be important in the development of an effective collateral circulation.
Arginine methylation is a common post-translational modification of proteins that is catalyzed by protein arginine methyltransferases (PRMTs). Arginine methylation is implicated in many cellular processes including transcription, RNA processing and transport, translation, signal transduction, DNA repair and apoptosis. 2 Degradation of these proteins containing methylated arginine residues results in release of symmetrically and asymmetrically methylated arginines. Among these, asymmetric dimethylarginine (ADMA) is a potent endogenous inhibitor of nitric oxide (NO) synthesis.
There is evidence that NO is important in collateral vessel development. Thus, blockade of NO synthase (NOS) activity by the NOS inhibitor
ADMA is cleared principally through metabolism by dimethylarginine dimethylaminohydrolase 1 (DDAH1) to
In the dog, brief repetitive periods of myocardial ischemia result in remarkable growth of collateral vessels. Thus, Weihrauch et al. 10 reported that 3 weeks of repetitive 2-minute coronary occlusions resulted in the development of collateral vessels sufficient to maintain normal resting tissue flow and contractile function in the ischemic zone during sustained total coronary artery occlusion. The present study was performed to examine regulation of the ADMA-DDAH1 pathway during this period when coronary collateral vessel development is known to occur. We used a canine model of recurrent myocardial ischemia to assess the regulation of myocardial interstitial ADMA levels and to examine spatial and temporal expression patterns of DDAH1, DDAH2, PRMT1, inflammatory factors, and protein arginine methylation in left ventricular (LV) myocardium. In addition, we used cultured human coronary artery endothelial cells (HCAEC) to define the influence of hypoxia and the inflammatory factors interleukin (IL)-1β and tumor necrosis factor (TNF)-α on endothelial DDAH1 expression. Our results provide new data suggesting a novel role for inflammatory factors in regulation of myocardial DDAH1 and ADMA in response to ischemia.
Methods
Surgical instrumentation and coronary occlusion protocol
Studies were performed in nine adult mongrel dogs (25–30 kg). As detailed below, four of these animals underwent intermittent coronary occlusions for 5 days (Group 1) and five underwent occlusions for 21 days (Group 2). In addition, four normal dogs were studied. All studies were carried out in accordance with the ‘Position of the American Heart Association on Research Animal Use’ and approved by the Animal Care Committee of the University of Minnesota. Animals in the repetitive occlusion groups were anesthetized with sodium pentobarbital (30–35 mg/kg IV), intubated and ventilated with 1.5% isoflurane. A left thoracotomy was performed and a hydraulic occluder was placed around the proximal left anterior descending coronary artery (LAD). A perforated intramyocardial catheter was introduced into the midwall of the LV in the perfusion territory of the LAD to allow sampling of interstitial fluid in the ischemic region, as previously described. 3 In animals in Group 2, a Doppler velocity probe was placed around the proximal LAD for monitoring of blood flow to insure total coronary occlusion during inflation of the occluder.
We used an intermittent coronary artery occlusion protocol to produce myocardial ischemia. 11 Beginning 10–14 days after surgery, eight 2-minute coronary occlusions were performed every 15 minutes for 2 hours, 5 days each week, by inflating the hydraulic occluder. Animals rested quietly in the laboratory and showed no evidence of distress during the 2-minute coronary occlusions. Animals in Group 1 were euthanized after 5 days of occlusions with an overdose of sodium pentobarbital and myocardium was harvested from the anterior ischemic LV region and from the posterolateral remote myocardial region. For animals in Group 2, intermittent occlusions were performed for 3 weeks and the artery was then permanently occluded. Collateralization is sufficient at this time to maintain myocardial viability during permanent coronary occlusion.3,12 After 5 days of permanent coronary occlusion, LV tissue from the ischemic and remote regions was harvested as described above.
Collection of interstitial fluid and tissue
Myocardial interstitial fluid (MIF) samples were collected from the animals in Group 2 each morning prior to beginning the daily coronary occlusions as previously described. 3 Samples were obtained by flushing 3 mL of isotonic saline into one end of the intramyocardial catheter as 3 mL of aspirate was withdrawn from the opposite end. Interstitial fluid samples were stored at −80°C for later analysis.
Measurement of ADMA
ADMA content in MIF was measured using the ADMA ELISA kit from DLD Diagnostika (Hamburg, Germany).
Western blot
Duplicate myocardial specimens from the ischemic zone and remote zone were quickly divided into three transmural layers representing subepicardium, midwall and subendocardium of the LV. The tissues were homogenized in lysis buffer containing 20 mM Tris·HCl, 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 1% Triton X-100, 2.5 mM sodium pyrophosphate, 1 mM β-glycerophosphate, 1 mM Na3VO4, 1 μg/mL leupeptin, 2 μg/mL aprotinin, and 1 mM PMSF. The homogenates were clarified by centrifugation at 12,000 g at 4°C for 10 minutes, and equal amounts of protein (80 μg) were subjected to 10% SDS-PAGE, transferred to HyBond nitrocellulose membrane (GE Healthcare Life Science, Pittsburgh, PA, USA), incubated with primary antibody followed by horseradish peroxidase-labeled secondary antibody, and detected by enhanced chemiluminescent substrate (Amersham; GE Healthcare Life Science, Pittsburgh, PA, USA) as previously described. 9 Signal intensities were quantified using ImageJ (US National Institutes of Health, Bethesda, MD, USA) and are presented as the arbitrary ratio to the signal intensities of sarcomeric α-actin. Primary antibodies included DDAH1 (1:2000 dilution; gift from Dr Kimoto), DDAH2 and dimethylarginine (1:1000, Abcam; Cambridge, MA, USA, ab87064), PRMT1 (1:3000, Sigma-Aldrich; St. Louis, MO, USA, P1620), α-sarcomeric actin (1:5000, Sigma-Aldrich, A2172) and β-actin (1:5000, Sigma-Aldrich, A1978), endothelial NOS (eNOS) (1:1000, Transduction Laboratories, BD Biosciences; San Jose, CA, USA, 610297), monomethylarginine (1:1000, Cell Signaling Technology; Danvers, MA, USA, 8015), dimethylarginine (1:1000, Abcam, ab-413), inducible NOS (iNOS) (1:200, Santa Cruz Biotechnology; Dallas, TX, USA, SC-651), HIF-1α (1:200, Santa Cruz Biotechnology, SC-10790), vascular endothelial growth factor (VEGF) (1:100, Santa Cruz Biotechnology, SC-507), IL-1β (1:100, Santa Cruz, SC-7884) and TNF-α (1:100, Santa Cruz Biotechnology, SC-8301).
Cell culture
HCAEC were purchased from Lonza (Allendale, NJ, USA), and cultured with EGM2-MV media; passages of p3-p6 were used in the studies. IL-1β and TNF-α were from R&D Systems (Minneapolis, MN, USA).
Statistical analysis
Results are presented as mean ± standard deviation unless otherwise indicated. Significance testing between groups was performed using Student’s t-test for unpaired data. Testing within groups was performed using Friedman’s ANOVA for repeated measures with post hoc testing using the Student–Neuman–Keuls method. Statistical significance was considered to be p< 0.05.
Results
ADMA and VEGF in interstitial fluid
As shown in Figure 1A, ADMA in MIF was 0.072±0.020 µmol/l before occlusions, decreased to 0.045 ± 0.016µmol/l at the first day after occlusions (p<0.05), and thereafter remained significantly below the baseline pre-occlusion level until day 21 at which time the experiment was terminated. There was a tendency for MIF ADMA levels to decrease further after day 1 of occlusions, but this was not significant. The low ADMA concentrations we found in MIF before occlusion, as compared to reports for other tissue fluids such as normal serum, plasma and aqueous humor,13,14 are due, at least in part, to dilution of the interstitial fluid with saline during the collection procedure. VEGF in MIF peaked at day 1 after occlusion, and waned during the following days of coronary occlusions (Figure 1B). Matsunaga et al. 3 reported that VEGF in MIF peaked at day 3 after initiation of repetitive occlusions in the same canine model of repetitive myocardial ischemia. The discrepancy between these two studies could be due to the technical variations. Nevertheless, both studies agree that expression of VEGF in the MIF peaked early during ischemia but waned as collateral growth progressed.
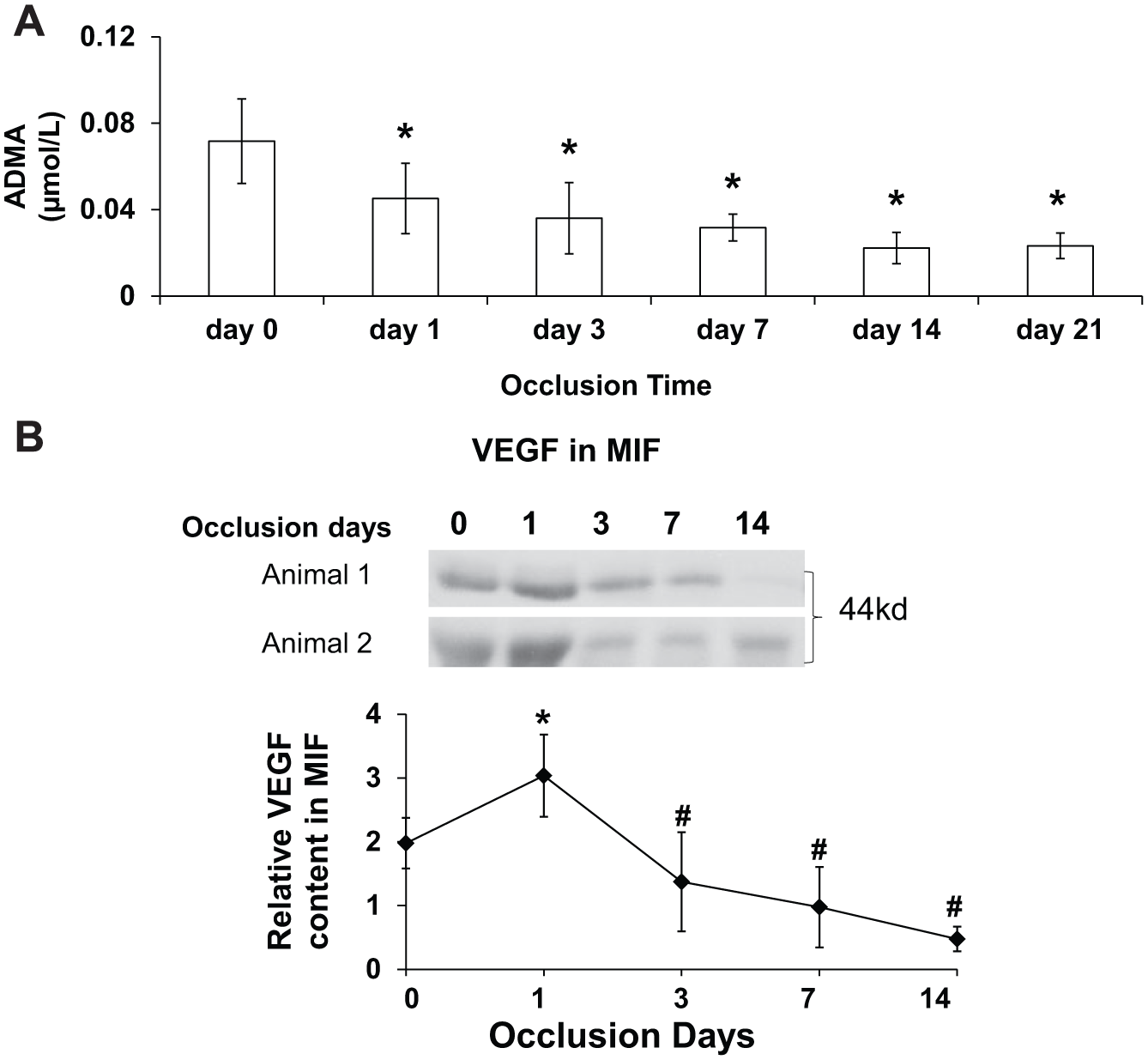
(A) ADMA in MIF is expressed as mean ± standard error (n=5). The ADMA level decreased on the first day after beginning coronary occlusions, further declined at day 3, and remained low through day 21 of occlusions when the experiment was terminated. (B) VEGF content in MIF. VEGF peaked at day 1 of occlusions and waned during subsequent occlusions.
Myocardial DDAH1, DDAH2 and PRMT1 expression
Methylation of arginines in myocardial proteins is catalyzed by protein arginine methyltransferases; the subsequent catabolism of these proteins results in liberation of free ADMA. In the heart, PRMT1 is responsible for arginine methylation while DDAH1 is the enzyme principally responsible for degrading ADMA. Therefore, we studied the protein expression levels of DDAH1, DDAH2 and PRMT1 to determine whether the decrease in ADMA levels that occurred during repetitive ischemia could be ascribed to decreased PRMT1 with decreased arginine methylation, or to increased DDAH1 and increased ADMA degradation. We first compared protein levels of DDAH1 and PRMT1 in three transmural layers of myocardium across the LV wall in four normal control animals. We found DDAH1 expression most abundant in the subepicardial myocardium, less in the midmyocardium, and least in the subendocardium (Figure 2A and 2B). PRMT1 expression was uniform across all three transmural layers. We then assayed myocardial tissue samples from the Group 1 dogs in which tissue was harvested after 5 days of coronary occlusions (Figure 2C and 2D). We found a similar gradient of DDAH1 across the three transmural layers with values highest in the subepicardium, while PRMT1 was again distributed uniformly across the LV wall. DDAH1 expression levels in the subendocardium and subepicardium were not different between the remote zone and ischemic zone, but DDAH1 expression was selectively increased in the midmyocardium of the ischemic zone and significantly higher than in the remote zone (Figure 2D). Since the position of the intramyocardial catheter from which MIF samples were obtained represented the midwall of the LV, further studies were performed using the midmyocardial tissue specimens.
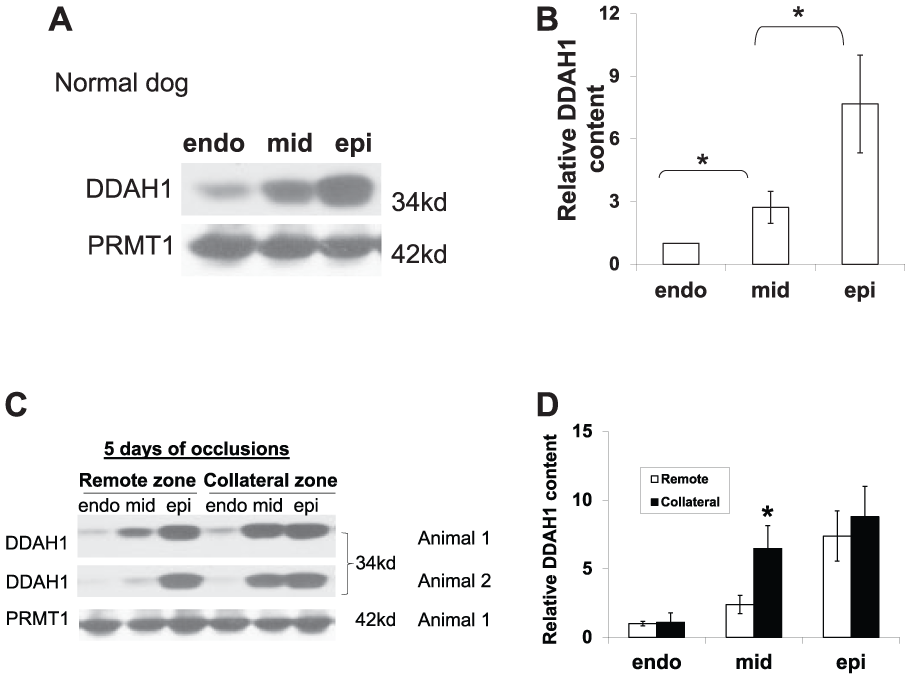
Expression of DDAH1 and PRMT1 in the subepicardium, midmyocardium and subendocardium of normal dogs (four animals) and dogs after 5 days of coronary occlusions (n=4). (A) Representative western blots of normal dogs. (B) Density quantification of western blots showed a gradient of distribution of DDAH1 in the three layers of LV myocardium of normal dogs with greatest abundance in the subepicardium, less in the midmyocardium, and least in the subendocardium. (C) Representative western blots of myocardium from dogs after 5 days of coronary occlusions. (D) Density quantification of western blots showed a gradient of distribution of DDAH1 in the three layers of LV myocardium from dogs after 5 days of coronary occlusions with increased DDAH1 expression in the midmyocardium from the ischemic zone as compared to the remote zone.
Western blots of the midmyocardial tissue lysates obtained after 3 weeks of coronary occlusions (five animals) demonstrated that the protein contents of DDAH2 and PRMT1 were similar to normal animals in both the ischemic and remote zones. However, DDAH1 content was significantly increased in the ischemic zone compared to both the remote zone and to normal hearts (Figure 3A and 3B).
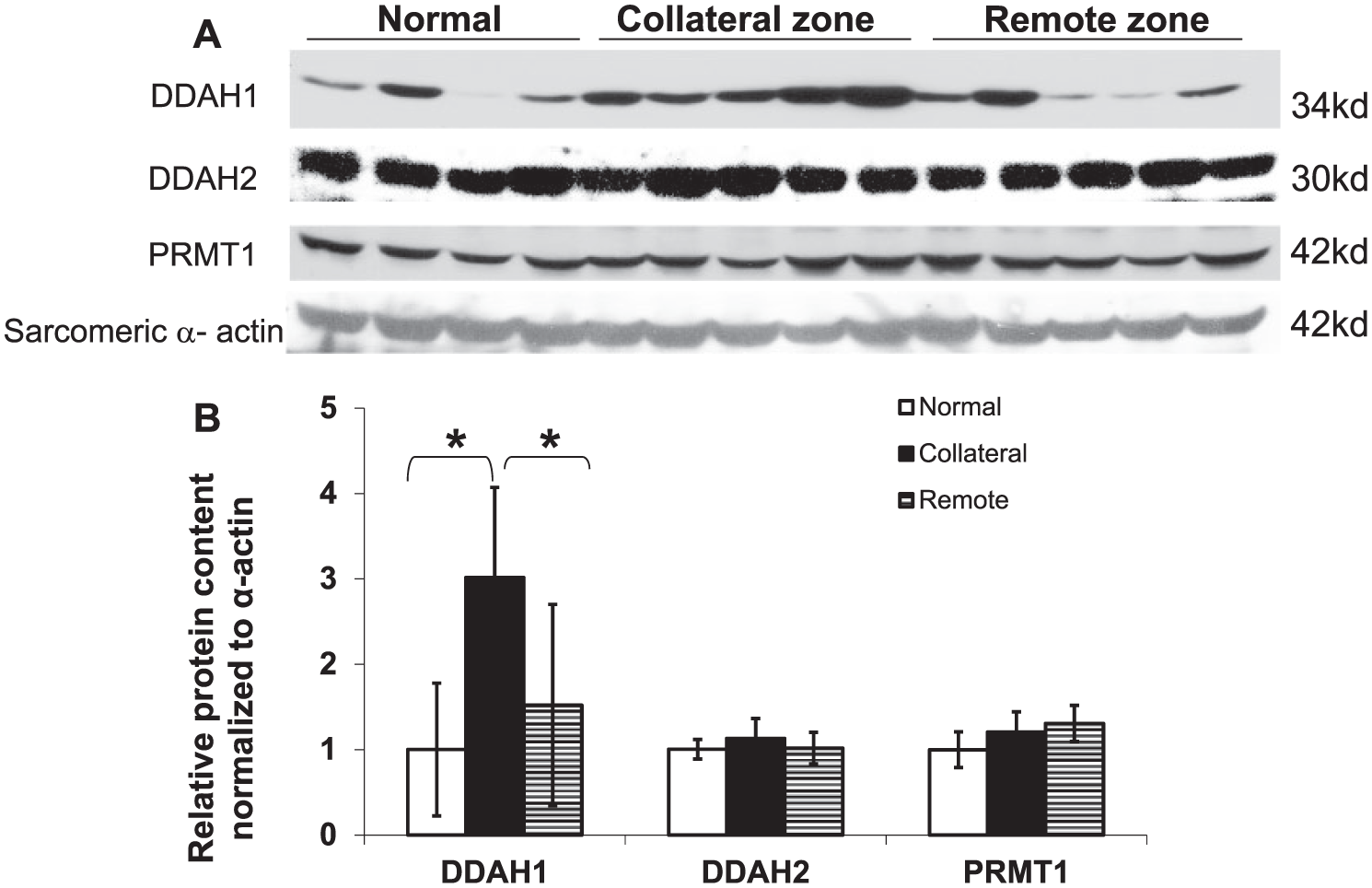
Expression of DDAH1, DDAH2 and PRMT1 in the midmyocardium after 3 weeks of coronary occlusions. (A) Representative western blots. (B) Density quantification of the western blots showed increased DDAH1 expression with no significant change of DDAH2 or PRMT1 in the ischemic zone as compared to either the remote zone or the midmyocardium from normal dogs (data from four normal dogs and five dogs with coronary occlusions,
Proteins with methylated arginine in the midmyocardium
Although we found that PRMT1 protein levels were similar in myocardium from the remote and ischemic zones (Figure 2C), and not significantly different from normal tissue (Figure 3B), this does not rule out the possibility that differences in PRMT1 activity might account for the observed differences in interstitial ADMA levels between the ischemic and remote zones. PRMT1 activity is regulated by formation of different macromolecular PRMT1 complexes that change accessibility of the enzyme to the substrate as well as through other post-translational modifications.2,15 Consequently, we examined the amount of proteins containing methylated arginines as an indicator of PRMT1 activity. Using antibodies to monomethylarginine and dimethylarginine, we found that, compared to normal animals, monomethylarginine-modified proteins were significantly increased in both the ischemic and remote zones of animals exposed to 3 weeks of coronary occlusions, with no difference between the ischemic and remote zones (Figure 4A and 4B). In contrast, the content of proteins with dimethylarginines was significantly increased in the ischemic zone as compared with both the remote zone and the midmyocardium from normal dogs (Figure 4C and 4D).
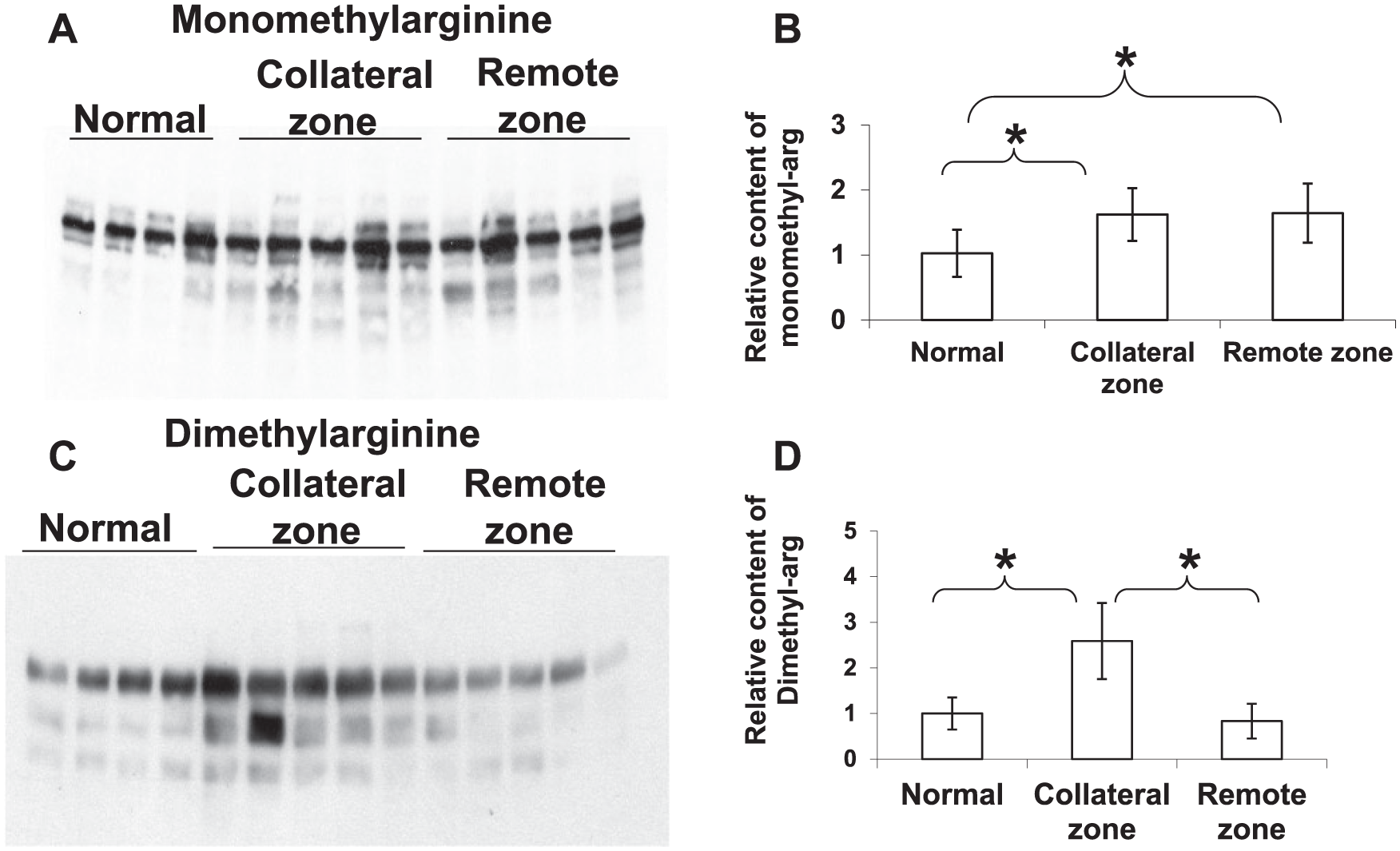
The levels of proteins with monomethylarginines and dimethylarginines in the midmyocardium of normal dogs, and from the remote and ischemic zones of dogs after 3 weeks of coronary occlusions. (A) Representative western blots using the monomethylarginine antibody. (B) Density quantification of the western blots showed that proteins with monomethylarginines (monomethyl-arg) were increased in both the ischemic and remote zones. (C) Representative western blots using the dimethylarginine antibody. (D) Density quantification of the western blots showed that proteins with dimethylarginine (dimethyl-arg) were selectively increased in the ischemic zone (data from four normal dogs and five dogs with coronary occlusions)
Inflammatory factors in the midmyocardium
Inflammatory processes are activated during collateral vessel development16–18 with inflammatory cells, including T cells, monocytes and natural killer cells in areas of collateral growth. 1 Mechanical stress can promote myocardial production of pro-inflammatory cytokines such as TNF-α and IL-6. 19 Since most previous studies examining inflammatory factors during the period of active coronary collateral vessel growth were done using plasma assays, we decided to study tissue levels of inflammatory factors in the midmyocardium of the hearts exposed to repetitive ischemia. In comparison to normal hearts, both TNF-α and IL-1β were significantly increased in the ischemic zone (Figure 5A–C). Furthermore, both TNF-α and IL-1β were significantly higher in the ischemic zone than in the remote zone. IL-1β was also significantly increased in the remote myocardial region compared to normal myocardium, while TNF-1α was not significantly different from normal.
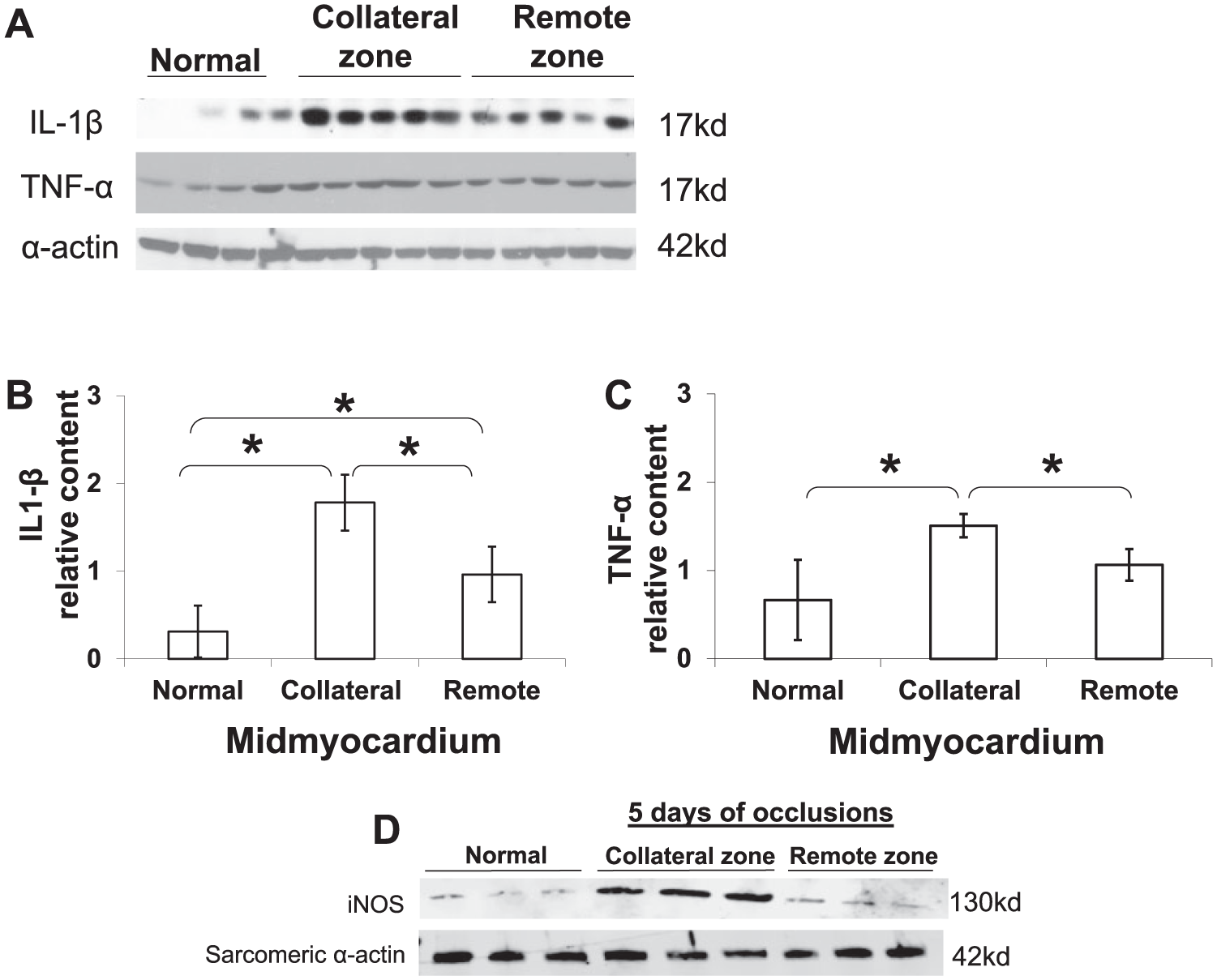
Protein levels of IL-1β and TNF-α in the midmyocardium from normal dogs and from the ischemic and remote zones after 3 weeks of coronary occlusions (A–C). (A) Representative western blots of IL-1β and TNF-α. (B) Density quantification of the western blots showed IL-1β was increased in the ischemic zone as compared to both the remote zone and to normal hearts. (C) Density quantification of the western blots showed TNF-α levels were increased in the ischemic zone as compared to both the remote zone and to myocardium from normal dogs (data from four normal dogs and five dogs with coronary occlusions). (D) Representative western blots of iNOS expression in the midmyocardium after 5 days of coronary occlusions; expression was increased in the ischemic zone compared to normal animals and to the remote zone of the hearts with coronary occlusions.
iNOS in the midmyocardium
Since the ischemia and its recruited inflammatory response can induce NOS expression, 19 and since NO is critical for coronary collateral development, 3 we evaluated the level of iNOS in the midmyocardium after 5 days of occlusions (Figure 5D). iNOS expression was increased in the collateral zone compared to both normal animals and the remote zone. We previously reported that iNOS expression and iNOS activity were increased in the collateral zone after 3 weeks of occlusions, and the coordinated expression of iNOS and eNOS acted to maintain collateral vasodilation to optimize the O2 supply–demand relationship and protect the collateralized myocardium from ischemia. 20 Therefore, iNOS expression is important for both collateral vessel development and collateral vasodilation.
Hypoxia and DDAH1 expression
Tissue hypoxia can be a driving force for vessel growth. The main mechanism of hypoxia-induced angiogenesis involves an increase in hypoxia-inducible factor (HIF) proteins. Consequently, we studied the effect of induction of HIF-1α on DDAH1 expression in cultured cells. In human coronary artery endothelial cells (HCAEC), treatment with CoCl2 (100 µM, 18 hours) robustly induced HIF-1α expression, but DDAH1 expression was not changed (Figure 6). In a separate study using neonatal rat cardiomyocytes, hypoxia also did not change the expression of DDAH1 (data not shown). These data indicate that hypoxia alone is not sufficient to increase DDAH1 expression.
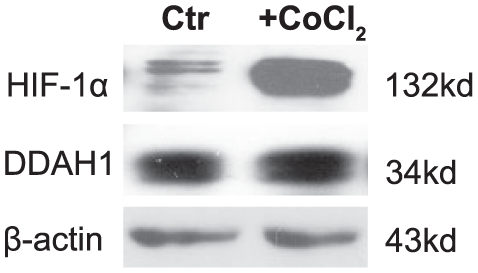
Representative western blots of the effect of hypoxia induced by cobalt chloride (CoCl2) on the expressions of DDAH1 and HIF-1α in HCAEC (n=3). Treatment with CoCl2 (100 µM, 18 hours) robustly induced HIF-1α expression while DDAH1 expression was not changed. Cells without CoCl2 treatment were control (Ctr). DDAH1, dimethylarginine dimethylaminohydrolase 1; HIF, hypoxia-inducible factor; HCAEC, human coronary artery endothelial cells; kd, molecular weight kilodalton.
Inflammatory factors and DDAH1 expression
IL-1β has profound effects on gene expression of endothelial cells 21 and plays an important role in angiogenesis. 22 Since we found that the content of both IL-1β and DDAH1 were increased in the ischemic zone, we hypothesized that IL-1β might regulate the expression of DDAH1 in endothelial cells. As shown in Figure 7A, treatment with IL-1β (10 units/ml) for 1–3 days significantly increased DDAH1 expression in HCAEC, while eNOS expression was unchanged. This result was consistent with an earlier report 23 that IL-1β induced DDAH1 expression in cultured rat smooth muscle cells. When we treated HCAEC with different concentrations of IL-1β for 1 day, we found that 0.01–10 units/mL of IL-1β caused dose-dependent increases of DDAH1 expression. However, further increases of IL-1β to 20–200 units/mL caused decreases of DDAH1 expression (Figure 7B). Thus, IL-1β has divergent effects on DDAH1 expression, with low concentrations increasing DDAH1 expression and high concentrations inhibiting DDAH1 expression. Concentrations of 100–200 units/mL IL-1β also caused morphologic changes in HCAEC with elongation of the cells to resemble the shape of fibroblasts (data not shown). HCAEC eNOS content was not significantly changed with low-dose IL-1β treatment (0.01–10 unit/mL), but decreased significantly with high IL-1β concentrations (20–200 units/mL) (Figure 7B). We found similar results when HCAEC cells were treated with TNF-α (Figure 8). These results suggest that low concentrations of IL-1β or TNF-α have effects that could act to enhance endothelial cell NO production, but that higher amounts of IL-1β or TNF-α exert deleterious effects by depressing both eNOS and DDAH1 expression.
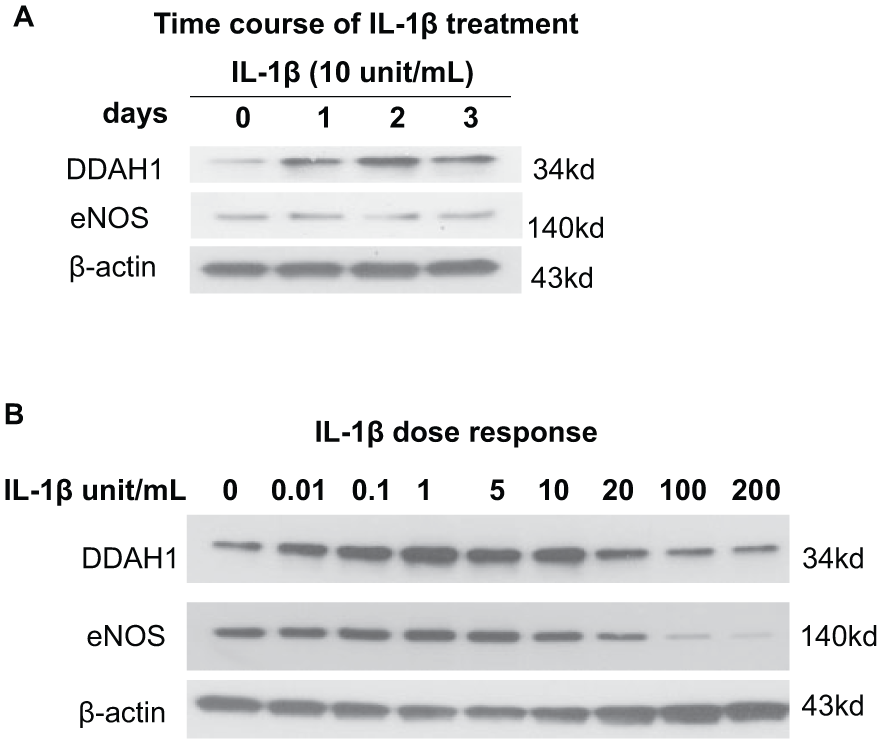
IL-1β-regulated DDAH1 and eNOS expression in HCAEC. (A) Time course of IL-1β (10 units/mL) on the expression of DDAH1 and eNOS (1–3 days). DDAH1 expression was increased from day 1 to day 3. (B) Dose–response to IL-1β (0.01–200 units/mL) on the expression of DDAH1 and eNOS after 24 hours of treatment. IL-1β had divergent effects on DDAH1 expression, with low concentrations causing increased DDAH1 expression and high concentrations suppressing DDAH1 expression (n=3). IL-1β, interleukin; DDAH1, dimethylarginine dimethylaminohydrolase 1; eNOS, endothelial nitric oxide synthase; HCAEC, human coronary artery endothelial cells; kd, molecular weight kilodalton.
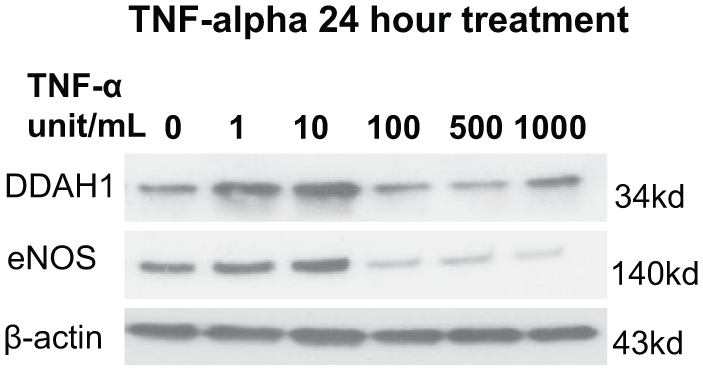
Dose–response of TNF-α (1–1000 units/mL) on the expression of DDAH1 and eNOS in HCAEC after 24 hours of treatment. TNF-α had divergent effects on DDAH1 expression, with low concentrations causing increased DDAH1 expression and high concentrations inhibiting DDAH1 expression (n=3). TNF-α, tumor necrosis factor; DDAH1, dimethylarginine dimethylaminohydrolase 1; eNOS, endothelial nitric oxide synthase; HCAEC, human coronary artery endothelial cells; kd, molecular weight kilodalton.
Discussion
Because DDAH1 regulates ADMA levels, NOS activity, and endothelial cell growth and function, we examined the spatial and temporal expression of myocardial DDAH1 in left ventricular myocardium in response to brief repetitive periods of myocardial ischemia that are known to induce collateral vessel development.3,10,12 We found that myocardial DDAH1 expression is significantly increased and interstitial fluid ADMA levels are decreased in the ischemic zone at a time that corresponds to active collateral vessel growth. 10 In addition, our combined in vivo and in vitro investigations into mechanisms of DDAH1 regulation identified a role for the inflammatory factors IL-1β and TNF-α in up-regulation of endothelial DDAH1 expression. Up-regulation of DDAH1 by inflammatory factors to remove ADMA during myocardial ischemia would be expected to augment NO generation and thereby promote collateral vessel development.
The endogenous NOS inhibitor ADMA is a major risk factor in patients with coronary artery disease (CAD).24,25 ADMA is functionally related to decreased NO bioavailability, eNOS uncoupling 26 and activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, the latter two of which result in increased oxidative stress. 27 Although collateral blood flow was not measured in the present study, using this experimental model myocardial blood flow during coronary occlusion was reported to be about 10% of normal zone blood flow at day 1 of coronary occlusions and increased to 81–88% of normal zone flow at day 21. 10 In this study, we found that ADMA in MIF was significantly decreased on the first day after beginning occlusions and remained low for the subsequent 3 weeks of occlusions. This reduction in ADMA levels would be expected to augment NO production during this phase of rapid collateral vessel growth.
ADMA is derived from the degradation of proteins containing methylated arginines. The expression of PRMT1 in myocardium from animals exposed to repetitive ischemia was not different from normal animals in either the remote zone or the ischemic zone. However, since the activity of PRMT1 can also be regulated, 2 we indirectly examined the activity of PRMT1 by measuring levels of methylated arginine containing proteins in myocardial lysates. We found that, in comparison to normal hearts, the level of dimethylarginine proteins was increased in the ischemic zone (but not the remote zone) of the hearts exposed to repetitive coronary occlusions, and levels of monomethylarginine proteins were significantly increased in both the ischemic and remote zones. This suggests that arginine methylation, and especially arginine dimethylation, is an important post-translational modification associated with recurrent myocardial ischemia. It might reasonably be expected that the increase in methylated arginines would result in increased ADMA production. Our finding that myocardial interstitial ADMA levels were decreased during recurrent ischemia could be explained by either decreased degradation of methylated proteins or increased metabolism of ADMA in the collateralizing hearts. Our finding that DDAH1 expression was increased in the midmyocardium of the ischemic zone (corresponding to the region from which the interstitial fluid was sampled) suggests that increased metabolism resulted in the lower ADMA levels in the collateralizing hearts.
DDAH1, the principal enzyme that degrades ADMA, is highly expressed in endothelial cells
28
where it can exert effects on endothelial function.
9
We found that DDAH1 expression was increased in the midmyocardium of the ischemic zone both after 5 days and 3 weeks of repetitive coronary occlusions. Because ADMA inhibits NOS activity, this increase in DDAH1 expression may help preserve NO-dependent collateral vessel development and dilation1,3 under conditions in which protein arginine methylation is increased. In support of the concept that increased DDAH1 expression increases eNOS activity and NO formation, a prior study from our lab found that inhibition of NOS activity with L-NG-Nitroarginine (
Interestingly, we found that DDAH1 expression in the LV wall is non-uniform, with the highest levels in the subepicardial myocardium, intermediate levels in the midwall, and lowest levels in the subendocardium. In response to repetitive myocardial ischemia, DDAH1 expression was specifically up-regulated in the midmyocardium. The mechanism for the specific increase of DDAH1 in the midmyocardium is not clear, but our in vivo findings, combined with our in vitro findings using cultured endothelial cells, suggest a role for inflammatory factors in the DDAH1 up-regulation. In myocardial tissue, both IL-1β and TNF-α were elevated specifically in the midmyocardium, similar to DDAH1. In vitro treatment of endothelial cells with low doses of IL-1β or TNF-α increased DDAH1 expression, suggesting a potential mechanism for the observed increase in DDAH1 expression in this region. Similarly, Ueda et al. found that IL-1β (1–100 units/mL) induced DDAH1 expression and decreased ADMA in vascular smooth muscle cells. 23 Inflammation is believed to play an important role in collateral vessel development.16–18 Our finding that IL-1β and TNF-α promote DDAH1 expression suggests a mechanism by which inflammation regulates NO signaling that could influence collateral vessel growth. Interestingly, hypoxia did not increase DDAH1 expression in cultured endothelial cells. In models involving prolonged total coronary artery occlusion, inflammatory processes are activated that could contribute to increased DDAH1 expression. However, when we performed hematoxylin and eosin staining of myocardium in this study, we observed no evidence of inflammatory cell infiltrate in either the ischemic or the remote myocardial regions (data not shown). Previous investigators have shown that cardiac myocytes can produce pro-inflammatory cytokines when triggered by ischemic tissue injury or mechanical stress. 31 Thus, it is likely that the increased expression of IL-1β and TNF-α, observed in the ischemic region in the present study, originated from cardiac myocytes or other cells resident in the myocardium rather than from infiltrating inflammatory cells.
In contrast to the DDAH1/NO promoting effects of low-dose IL-1β and TNF-α treatment, high concentrations of IL-1β and TNF-α decreased both DDAH1 and eNOS expression. Previous cell culture studies have shown that TNF-α decreases eNOS expression by reducing the half-life of the mRNA encoding eNOS.32,33 It has also been reported that human eNOS promoter activity is reduced by TNF-α and that the reduced activity is restored by co-transfection of dominant negative IKKβ (inhibitor of nuclear factor Kappa-B kinase subunit beta). 34 IL-1β also has been shown to decrease eNOS mRNA levels in endothelial cells. 35 The high doses of IL-1β and TNF-α that we used in this study are in the same range as the aforementioned studies that demonstrated down-regulation of eNOS by IL-1β and TNF-α in endothelial cells. Thus, our finding of decreased eNOS expression under high-dose IL-1β or TNF-α treatment are consistent with those previous studies. To the best of our knowledge, our findings of the biphasic effects of IL-1β and TNF-α treatment on DDAH1 expression are novel. Determining the mechanism of this regulation will be an interesting topic for further study.
In summary, brief repetitive episodes of myocardial ischemia caused increased expression of DDAH1 in the midwall of the LV that was accompanied by decreased interstitial fluid ADMA levels. Our observation that the up-regulation of DDAH1 expression was associated with elevated IL-1β and TNF-α in the LV midwall of the collateral zone, and that low doses (but not high doses) of IL-1β and TNF-α were sufficient to increase DDAH1 expression in isolated endothelial cells, identifies a relationship between inflammation and DDAH1 expression that could act to enhance collateral vessel growth. It has been suggested that strategies directed at stimulation of vessel growth by activation of inflammatory cells might be a therapeutic target for patients with ischemic heart disease. 36 Considering the importance of DDAH1 and ADMA in regulation of NO production, our findings provide an important aspect to be considered in applying such therapeutic strategies.
Limitations
Results from clinical trials of interventions to enhance collateral vessel development have demonstrated that age, hypercholesterolemia, genetic susceptibility, diabetes and smoking (frequently present together) all adversely affect the capacity for collaterals to develop. 37 The present study was conducted in healthy canines that were free from these risk factors. It is possible that the presence of these risk factors in patients with coronary disease could influence the responses of DDAH1 that we observed in the healthy animals in the present study. We have previously found DDAH1 transcript and protein levels in failing human myocardium from patients with congestive cardiomyopathy. 38 However, the effect of ischemia on DDAH1 in the human heart has not been studied. Thus, the present results must be interpreted with caution; the finding that intermittent brief bouts of ischemia increased DDAH1 expression in canine myocardium cannot be directly extrapolated to the human heart.
Conclusions
Repetitive episodes of ischemia caused by brief coronary occlusions resulted in an increase in DDAH1 protein in the midwall of the ischemic myocardium as well as decreased ADMA in MIF. The increased DDAH1 expression likely resulted, at least in part, from the up-regulation of IL-1β and TNF-α in the ischemic region, since low concentrations of these inflammatory mediators, but not hypoxia, caused an increase of DDAH1 expression in coronary artery endothelial cells. The up-regulation of DDAH1 to remove ADMA may be an important mechanism to augment NO generation in the ischemic region and thereby promote coronary collateral vessel development.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this study was supported by US Public Health Service Grants HL20598, HL21872 and HL71790 from the National Heart, Lung, and Blood Institute. Drs Zhang, Fassett and Hu are recipients of Scientist Development Awards from the American Heart Association.