
Abstract
Recent studies have shown mitochondrial dysfunction and increased production of reactive oxygen species in peripheral blood mononuclear cells (PBMCs) and endothelial cells from patients with diabetes mellitus. Mitochondria oxygen consumption is coupled to adenosine triphosphate (ATP) production and also occurs in an uncoupled fashion during formation of reactive oxygen species by components of the electron transport chain and other enzymatic sites. We therefore hypothesized that diabetes would be associated with higher total and uncoupled oxygen consumption in PBMCs that would correlate with endothelial dysfunction. We developed a method to measure oxygen consumption in freshly isolated PBMCs and applied it to 26 patients with type 2 diabetes mellitus and 28 non-diabetic controls. Basal (192±47 vs 161±44 pmoles/min, p=0.01), uncoupled (64±16 vs 53±13 pmoles/min, p=0.007), and maximal (795±87 vs 715±128 pmoles/min, p=0.01) oxygen consumption rates were higher in diabetic patients compared to controls. There were no significant correlations between oxygen consumption rates and endothelium-dependent flow-mediated dilation measured by vascular ultrasound. Non-endothelium-dependent nitroglycerin-mediated dilation was lower in diabetics (10.1±6.6 vs 15.8±4.8%, p=0.03) and correlated with maximal oxygen consumption (r = −0.64, p=0.001). In summary, we found that diabetes mellitus is associated with a pattern of mitochondrial oxygen consumption consistent with higher production of reactive oxygen species. The correlation between oxygen consumption and nitroglycerin-mediated dilation may suggest a link between mitochondrial dysfunction and vascular smooth muscle cell dysfunction that merits further study. Finally, the described method may have utility for the assessment of mitochondrial function in larger scale observational and interventional studies in humans.
Introduction
In addition to generating adenosine triphosphate (ATP) via oxidative phosphorylation, mitochondria are important for many other aspects of cellular function, including calcium flux, apoptosis, and intracellular signaling. 1 It is now recognized that altered mitochondrial function contributes to the aging process and many common disease states, including neurodegenerative diseases, cancer, and atherosclerosis.2–5 In light of the ongoing obesity epidemic, there is also considerable interest in mitochondrial dysfunction in the pathogenesis of type 2 diabetes mellitus and its vascular complications.6–11 Recent work has demonstrated increased mitochondrial production of reactive oxygen species (ROS), oxidative stress, and altered mitochondrial morphology and dynamics in peripheral blood mononuclear cells and endothelial cells from patients with diabetes mellitus.12–15
Cellular oxygen consumption is increasingly recognized as a fundamental measure of mitochondrial function.16–18 The basal oxygen consumption rate reflects coupled mitochondrial respiration as well as uncoupled consumption of oxygen to form ROS at mitochondrial and non-mitochondrial enzymatic sites. Coupled and uncoupled respiration can be distinguished by examining the effects of an inhibitor of ATP synthase (oligomycin) and/or complex III (antimycin A). The maximal oxygen consumption rate, provoked by the addition of a mitochondrial uncoupling agent such as carbonyl cyanide p
Study of mitochondrial respiration in humans has previously been limited by a number of technical issues. The Clark electrode has been used to examine oxygen consumption in human leukocytes in suspension,20,21 but its requires a relatively large volume of blood. Attempts to measure oxygen consumption in blood cells using the Seahorse device have been limited by displacement of non-adherent cells during the automated movement of the oxygen sensor. For the present study, we developed a method to immobilize readily available peripheral blood mononuclear cells (PBMCs) that allows us to measure mitochondrial oxygen consumption with the Seahorse XF96 analyzer (Seahorse Bioscience, North Billerica, MA, USA). We used the method to measure mitochondrial function in patients with type 2 diabetes mellitus and non-diabetic controls and related the results to vascular function. Prior studies have shown increased mitochondrial production of ROS in PBMCs, endothelial cells, and other cell types in diabetes, suggesting systemic mitochondrial dysfunction.12,13,22 On this basis, we hypothesized that diabetes mellitus would be associated with higher total and uncoupled oxygen consumption that would correlate with endothelial dysfunction, providing evidence in humans that mitochondrial dysfunction might be relevant to cardiovascular disease in humans.
Materials and methods
Study subjects
Adult patients with type 2 diabetes mellitus and healthy volunteers were recruited at Boston Medical Center by advertisement. Diabetes was defined as fasting glucose ≥126 mg/dl or ongoing treatment for type 2 diabetes mellitus. Healthy volunteers were taking no medications, had a fasting glucose <100 mg/dl, and had never smoked or had stopped smoking for at least a year prior to enrollment. A fasting blood sample was obtained in the morning by venipuncture, and glucose and lipids were measured in the Boston Medical Center Clinical Laboratory. Non-invasive measurement of endothelium and non-endothelium-dependent vasodilator function was then completed, as described below. The study protocol was approved by the Boston Medical Center Institutional Review Board and all participants provided written informed consent.
Materials and reagents
The study used the following materials and reagents: FCCP (Sigma Life Science, St Louis, MO, USA), oligomycin (Sigma Life Science), poly-
Isolation of PBMCs
Venous blood was collected into commercially available cell preparation tubes for PBMCs (BD Vacutainer CPT with sodium citrate; Becton, Dickinson and Company, Franklin, NJ, USA). The tubes were spun at 1650 g for 30 minutes at room temperature and cell layers were collected and transferred into 25 mL of HBSS. This solution was centrifuged for an additional 10 minutes at 250 g at room temperature. The pellet was resuspended in 15 mL of fresh HBSS and the cell count was determined using a glass hemocytometer (Double Neubauer Counting Chamber; Hausser Scientific, Horsham, PA, USA). Fluorescence activated cell sorting (FACS) analysis in the Boston Medical Center Hematology Laboratory of samples from five subjects demonstrated that this isolation method yielded a cell population of 82±4% lymphocytes (CD3 positive cells) and 14±2% monocytes (CD14 positive cells), with the remaining 4% comprised of other mononuclear cells.
Plate preparation
Prior to the addition of PBMCs, XF96 plates (Seahorse Bioscience) were treated with poly-
PBMCs were suspended in DMEM supplemented with pyruvate 1.0 mM and glucose 5.5 mM. The DMEM was warmed to 37°C and the pH was adjusted to 7.4 prior to cell suspension. PBMCs (600,000 per well) were seeded onto poly-
Oxygen consumption measurement protocol
Oxygen consumption rates were measured in accordance with manufacturer instructions (Seahorse Bioscience). The assay media (DMEM) was supplemented with glucose (5.5 mM) and pyruvate (1 mM) and the pH was adjusted to 7.4. In preliminary studies testing 200,000 to 1 million cells per well, we found an optimal signal and uniform distribution of cells on the bottom of the well with the use of 600,000 cells per well. Experiments were replicated in six wells and averaged for each experimental condition. The average coefficient of variation for the basal oxygen consumption rate was 6.4±2.0%.
Oxygen consumption measurements were made approximately every 8 minutes under basal conditions, after the addition of a saturating concentration of oligomycin (5 µmoles/L), 23 and after the addition of FCCP (1 µmoles/L). Basal and post-oligomycin respiration rates were calculated by averaging the last four measurements after achieving a steady state. Coupled respiration was expressed as the percent decrease from basal respiration. Maximal FCCP respiration was taken as the highest measurement after the addition of FCCP.
Assessment of vascular function
We used vascular ultrasound to measure endothelium-dependent flow-mediated dilation of the brachial artery as previously described. 24 Subjects fasted and withheld all medications on the morning of study. Two-dimensional ultrasound images of the brachial artery were recorded before and 1 minute after induction of reactive hyperemia by 5-minute cuff occlusion of the upper arm. We measured Doppler flow velocity at baseline and immediately after cuff release. Non-endothelium-dependent dilation of the brachial artery was assessed by examining the change in brachial diameter induced by sublingual nitroglycerin (0.4 mg). The nitroglycerin portion of the vascular testing protocol was omitted if the participant had systolic blood pressure less than 100 mmHg or had a history of migraine headaches or nitrate intolerance.
Statistical analyses
Clinical characteristics of the diabetic and healthy control groups were compared using the unpaired t-test or chi-squared test for continuous and categorical variables respectively. Group differences (controls versus diabetic patients) in oxygen consumption were compared using the Wilcoxon signed rank test because examination of histograms suggested that these variables lacked a normal distribution. Linear regression analysis was used to determine the relation between log-transformed oxygen consumption rates and diabetes mellitus while adjusting for age, sex, and black race. Changes in oxygen consumption over time according to group (controls and diabetic patients) were assessed using analysis of variance for repeated measures after log transformation of the oxygen consumption variables. We calculated Spearman’s correlation coefficients to test the relation between vascular function variables and oxygen consumption rates. P<0.05 was considered to be statistically significant. Data are expressed as mean±SEM. Statistical analyses were completed using SPSS Version 20 (IBM, Inc.).
Results
Study subjects
We enrolled 54 subjects (26 with diabetes mellitus and 28 controls). Their clinical characteristics are shown in Table 1. The two groups were similar in age and sex. As expected, the patients with diabetes mellitus had higher fasting glucose and body mass index, lower high-density lipoprotein (HDL) cholesterol, and a trend toward higher triglyceride levels (p=0.05). Total and low-density lipoprotein (LDL) cholesterol levels were lower in diabetic patients, likely reflecting the ongoing treatment with cholesterol-lowering drugs. Also as expected, diabetic patients were taking drugs to lower blood glucose and blood pressure (Table 1).
Clinical characteristics.
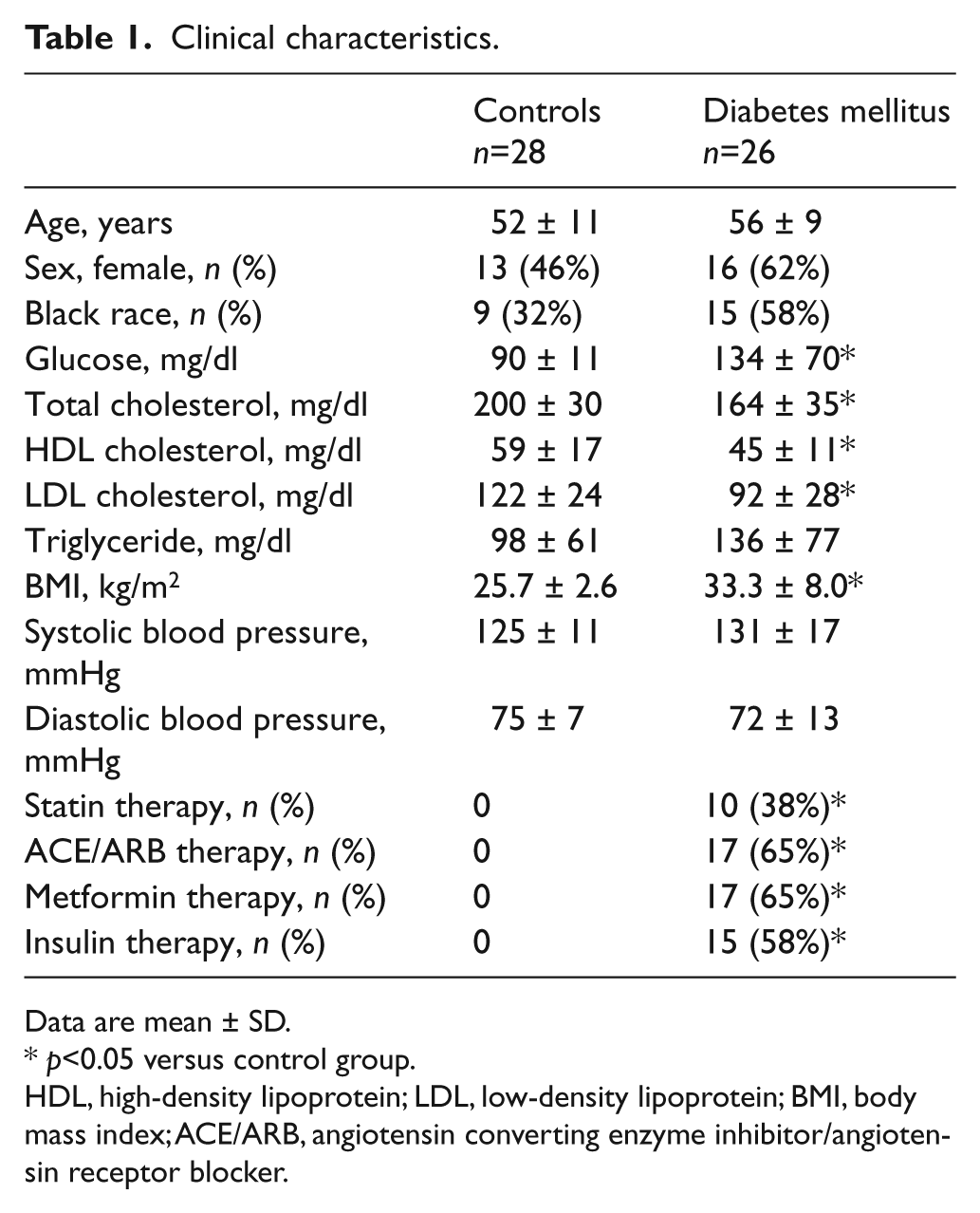
Data are mean ± SD.
p<0.05 versus control group.
HDL, high-density lipoprotein; LDL, low-density lipoprotein; BMI, body mass index; ACE/ARB, angiotensin converting enzyme inhibitor/angiotensin receptor blocker.
Oxygen consumption rates in controls and diabetic patients
Table 2 displays oxygen consumption rates in PBMCs from controls and diabetic patients. As shown, basal, post-oligomycin (uncoupled), and maximal after FCCP oxygen consumption rates were all higher in diabetic patients compared to controls. Interestingly, coupled respiration, calculated as the percent decrease after oligomycin, did not differ between groups. These findings are consistent with our hypothesis that diabetes would be associated with higher uncoupled respiration due to higher production of ROS. The sequence of reagents and time course of oxygen consumption measurements is displayed in Figure 1. As shown, oxygen consumption was consistently higher in diabetic patients.
Diabetes mellitus and oxygen consumption rates in PBMCs.
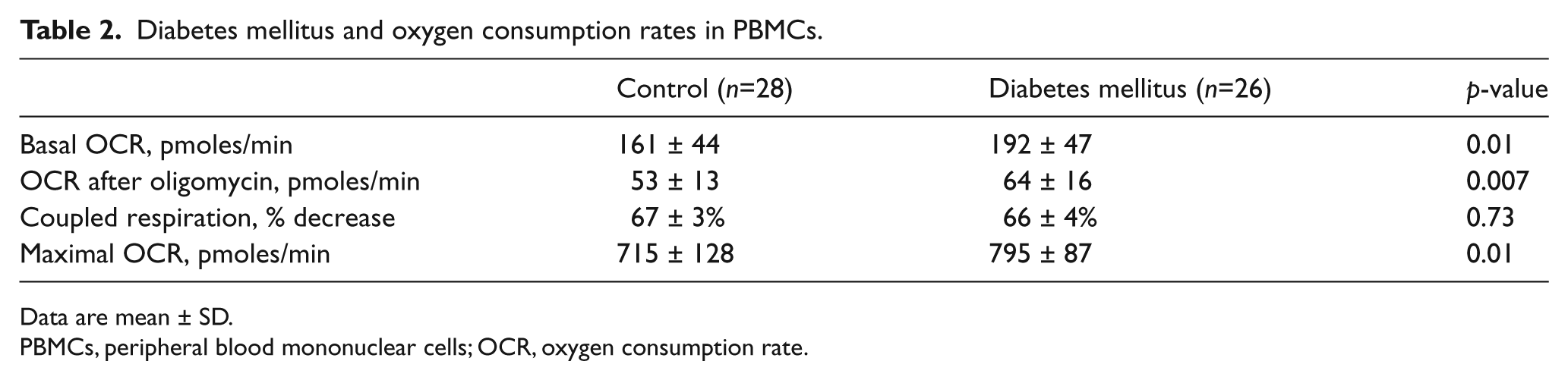
Data are mean ± SD.
PBMCs, peripheral blood mononuclear cells; OCR, oxygen consumption rate.
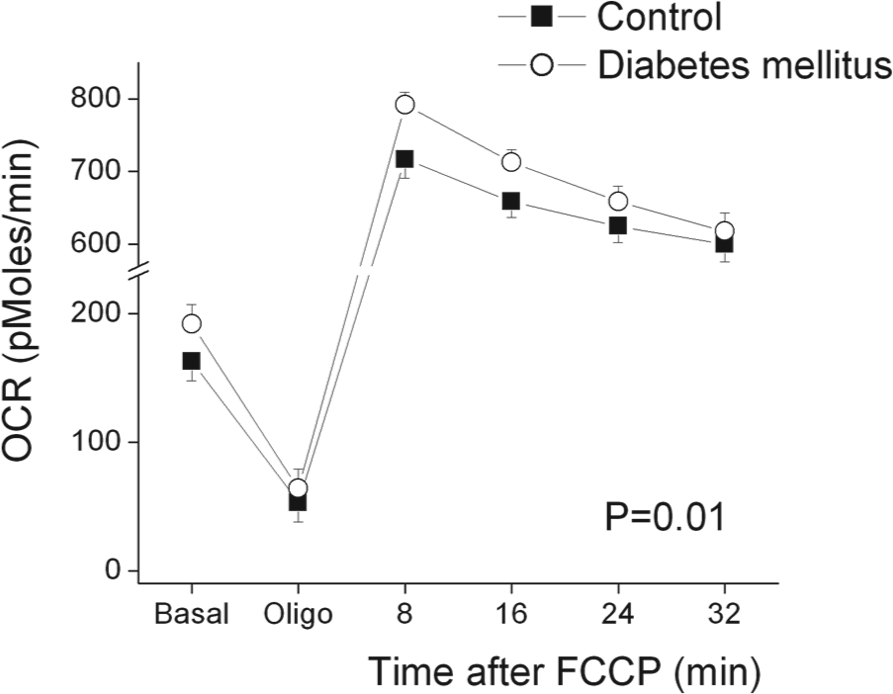
Oxygen consumption rates (OCR) in peripheral blood mononuclear cells (PBMCs) from control subjects (n=28) and patients with type 2 diabetes mellitus (n=26). The OCR was measured approximately every 8 minutes using an XF96 analyzer as described in Methods. Basal and post-oligomycin (5 µmoles/L) rates were calculated by averaging the last four measurements after achieving a steady state. Sequential measurements were also made after the addition of carbonyl cyanide p-trifluoromethoxyphenylhydrazone (FCCP) (1 µmoles/L), which depolarizes the mitochondrial membrane and induces maximal oxygen consumption. As shown, oxygen consumption rates were higher in PBMCs from diabetic patients (p=0.01 for diabetes main effect and p=0.03 for diabetes by time interaction by repeated measures ANOVA).
Other clinical correlates of oxygen consumption rates
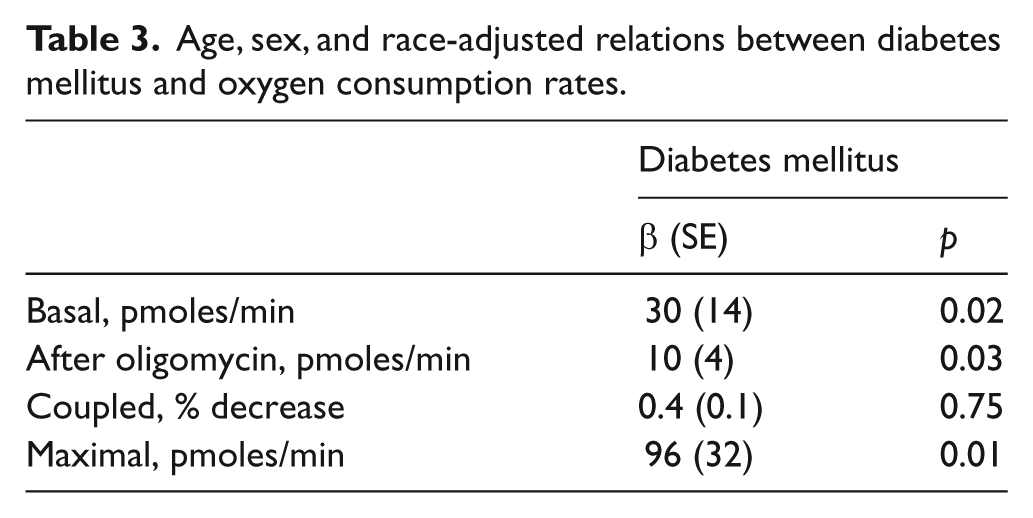
We noted that there were non-significant tendencies for higher age (p=0.13), more females (p=0.27), and more black participants (p=0.06) in the diabetic group (Table 1), although none of these clinical characteristics correlated with oxygen consumption rates. As shown in Table 3, diabetes mellitus was associated with significantly higher basal, post-oligomycin, and maximal oxygen consumption rates in multivariable models adjusting for age, sex, and black race, suggesting that confounding effects of these factors do not explain our findings.
Age, sex, and race-adjusted relations between diabetes mellitus and oxygen consumption rates.
As shown in Table 1, some of the diabetic subjects were taking a statin, angiotensin converting enzyme inhibitor/angiotensin receptor blocker (ACEI/ARB), metformin, or insulin. We compared basal, post-oligomycin, maximal, and coupled oxygen consumption rates according to medication status among the diabetic subjects. Diabetic patients taking insulin had slightly higher coupled respiration (68±4%, n=15) compared to diabetics not taking insulin (64±3%, n=11; p=0.04), possibly suggesting that insulin treatment promotes mitochondrial respiration. Insulin treatment did not relate to the other measures of oxygen consumption (data not shown). Oxygen consumption rates also did not differ in diabetic patients who were and were not taking a statin, ACEI/ARB, or metformin (data not shown).
We explored the other clinical correlates of oxygen consumption in the whole cohort. We observed no relation between basal, post-oligomycin, post FCCP, or coupled respiration with age, sex, blood glucose, blood lipids, body mass index, or blood pressure (data not shown).
Dependence of oxygen consumption rates on fatty acid transport
Type 2 diabetes mellitus is associated with insulin resistance and decreased expression of glucose transporters that might limit entry of glucose into PBMCs. 25 As shown in Table 1, we observed a strong trend toward higher serum triglycerides in our participants (p=0.05). A decrease in glucose transport and higher triglycerides would both be expected to cause a shift away from glucose and toward fatty acids as a source of fuel in mitochondria. We therefore investigated whether differences in fatty acid utilization might account for the higher oxygen consumption rate observed in our diabetic subjects. As shown in Figure 2, inhibiting fatty acid transport into mitochondria with the carnitine O-palmitoyltransferase-1 (CPT-1) inhibitor etomoxir produced a dose-dependent decrease in basal oxygen consumption that did not differ between control and diabetic subjects. This result argues against a disproportionate dependence on fatty acid oxidation in PBMCs in diabetes mellitus.
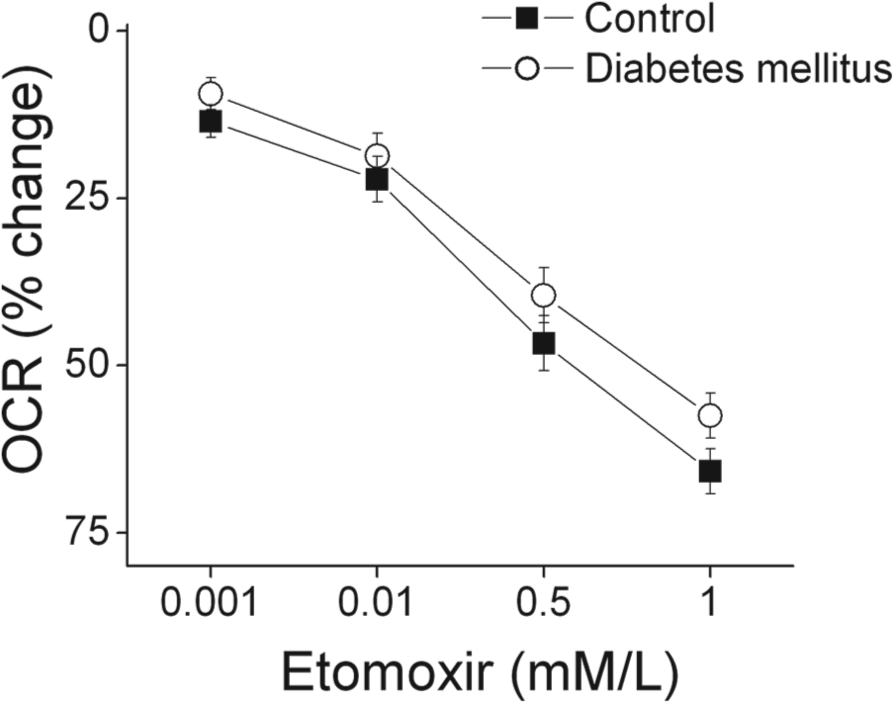
Etomoxir reduces the basal oxygen consumption rate (OCR) to an equivalent extent in diabetic patients and controls. The oxygen consumption rate was measured in peripheral blood mononuclear cells (PBMCs) incubated with the indicated concentrations of etomoxir as described in Methods. As shown, inhibiting fatty acid uptake produced a dose-dependent decrease in oxygen consumption (p<0.001) in both diabetic (n=5) and control subjects (n=5), but the response did not differ between groups (p=0.64 by analysis of variance for repeated measures).
Relation of oxygen consumption rates to vascular function
Table 4 displays vasodilator function in the control and diabetic subjects. As shown, endothelium-dependent flow-mediated dilation was lower in the diabetic subjects, whether expressed as percent change or absolute change (mm). Non-endothelium-dependent dilation of the brachial artery in response to sublingual nitroglycerin was also lower in the diabetic subjects. Finally, we observed a non-significant trend for lower reactive hyperemia, a measure of microvascular function, in the diabetic subjects. Since reactive hyperemia is the stimulus for flow-mediated dilation, reduced hyperemia might have accounted for our finding that flow-mediated dilation was lower in the diabetic subjects. However, diabetics had lower flow-mediated dilation after adjustment for reactive hyperemia, arguing against this possibility and providing additional evidence for impaired conduit artery endothelial dysfunction in the diabetic subjects.26,27
Diabetes mellitus and vascular function.
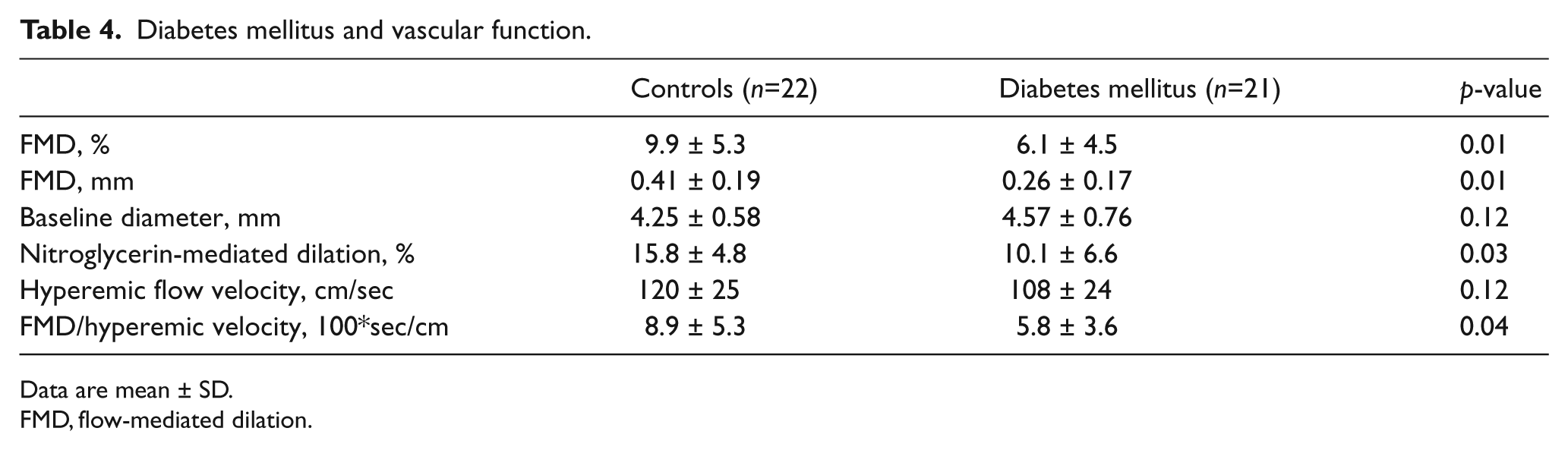
Data are mean ± SD.
FMD, flow-mediated dilation.
We investigated the relationships between oxygen consumption rates and vascular function. There were no significant correlations between brachial artery flow-mediated dilation (Figure 3, lower panel), hyperemic flow velocity, baseline diameter, and the measures of oxygen consumption (data not shown). Interestingly, there was a strong inverse correlation between nitroglycerin-mediated dilation and peak oxygen consumption rate (Figure 3, upper panel).
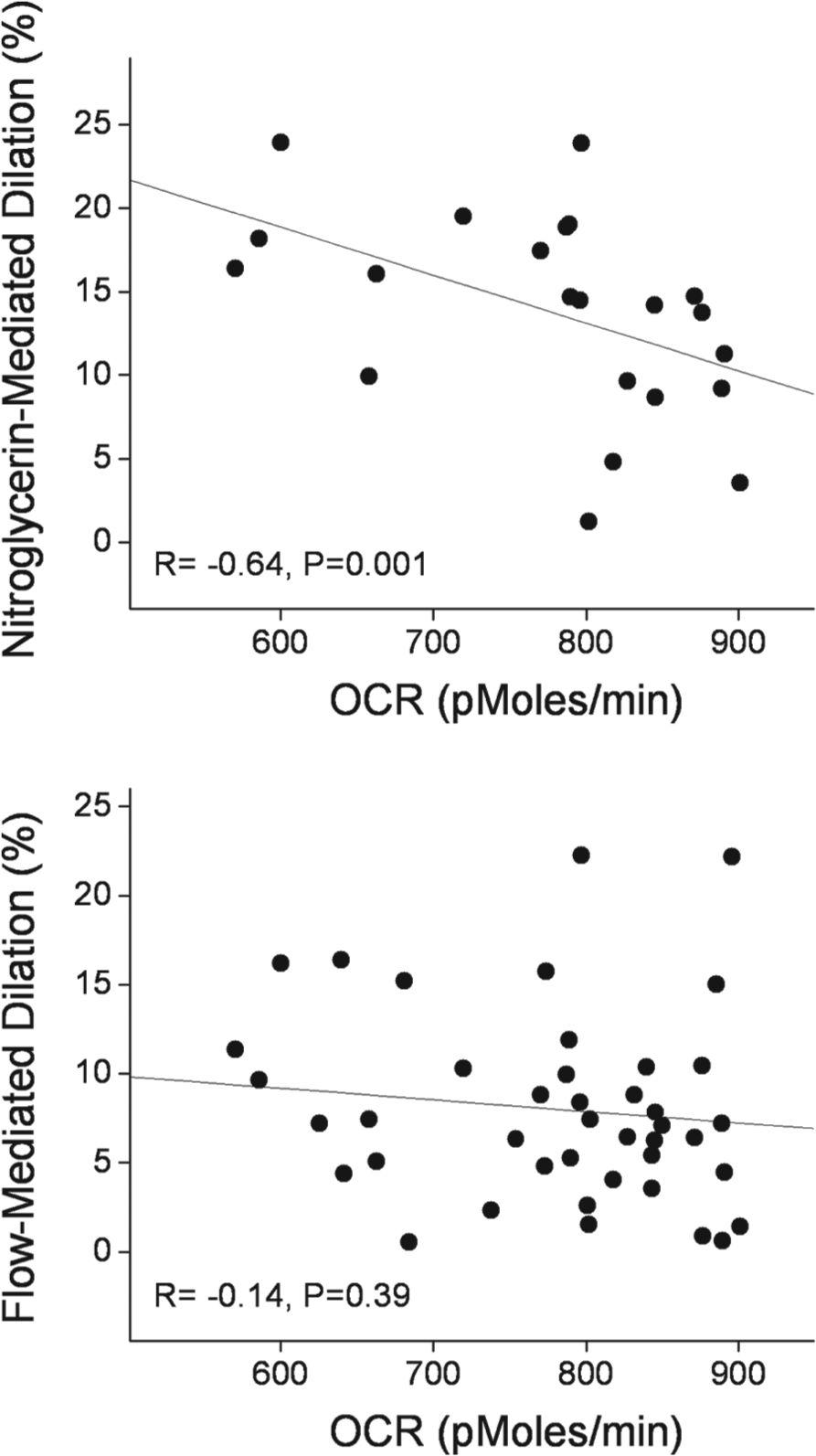
Peak oxygen consumption in PBMCs correlates with nitroglycerin-mediated dilation (upper panel), but not flow-mediated dilation (lower panel).
Discussion
In this study, we developed a method that allowed for measurement of mitochondrial oxygen consumption in PBMCs and related the findings to vascular function in patients with type 2 diabetes mellitus. We observed higher basal, uncoupled, and maximal oxygen consumption in the diabetic group, findings consistent with our hypothesis that increased production of reactive species in diabetes mellitus would lead to an increase in uncoupled respiration. Inhibiting fatty acid transport into mitochondria with etomoxir lowered oxygen consumption to a similar extent in the two groups, suggesting that a shift toward fatty acid utilization does not explain our findings. Contrary to our initial hypothesis, we observed no significant correlation between measures of endothelial vasodilator function and oxygen consumption. There was a significant correlation between nitroglycerin-mediated dilation and maximal oxygen consumption in PBMCs, raising the possibility that mitochondrial function and respiratory reserve might relate to the function of vascular smooth muscle cells. Overall, the study provides evidence that diabetes mellitus is associated with increased uncoupled and maximal respiration in PBMCs that might be relevant to inflammation, altered immune function, and cardiovascular disease.
Our described method offers several advantages over other methods for assessing mitochondrial function in human subjects. Although the Clark electrode can be used to measure oxygen consumption rate in blood cells in suspension, the presented method uses fewer cells (6×105 vs 107), reflecting the greater sensitivity of the Seahorse device.20,21,28 Other approaches for measuring mitochondrial function in humans have included measurement of mitochondrial enzyme activity in biopsied tissue and by magnetic resonance spectroscopy.29,30 Such studies are invasive or require expensive equipment, and are poorly suited to the larger-scale epidemiological or intervention studies that would be required to understand how mitochondrial function relates to disease risk or management.
Prior studies have shown that mitochondrial function is relevant to leukocyte function. For example, mitochondrial membrane hyperpolarization and the production of ROS contribute to immune activation of monocytes and lymphocytes.31,32 Maturation of memory T cells involves mitochondrial biogenesis and an increase in mitochondrial reserve capacity. 33 Several prior studies used a Clark electrode to examine oxygen consumption in PBMCs. Belikova and colleagues demonstrated higher and more uncoupled oxygen consumption in patients with sepsis. 20 Kuhnke and colleagues showed that patients with rheumatologic disease have higher basal oxygen consumption rates that correlate with activity of disease, increase with mitogenic stimulation, and decrease following steroid treatment. 21 It is possible that systemic inflammation and activation of PBMCs in diabetes mellitus might relate to increased oxygen consumption in the present study. It is important to acknowledge that we assessed quiescent cells and a mixed population of lymphocytes and monocytes in the present study. As has recently been emphasized, it would be interesting to examine activated cells and to compare results in monocyte and lymphocyte subsets to gain further insight into the importance of mitochondrial function for leukocyte biology. 34
Several prior studies specifically measured mitochondrial function in other types of blood cells from patients with diabetes mellitus. Hernandez-Mijares and colleagues reported lower oxygen consumption rates measured by a Clark electrode in neutrophils from diabetic patients.
35
Avila and colleagues reported lower oxygen consumption rates in platelets from diabetic patients measured with the Seahorse XF24 analyzer.
36
In that study, Cell-Tak (BD Biosciences (San Jose, CA) was used to enhance platelet adherence to the wells. Cell-Tak was also used in a very recent report examining oxygen consumption rates in human leukocytes and platelets
34
and appears to be a reasonable alternative to poly-
We previously reported higher mitochondrial ROS production, higher membrane potential, a shift toward mitochondrial fission, and lower mitochondrial mass in PBMCs that were isolated from diabetic patients by the method used in the present study. 12 Our new findings add to that prior work and provide additional evidence for altered mitochondrial function in diabetes using an emerging and potentially more clinically relevant approach. Higher ROS production likely accounts for the higher basal, post-oligomycin, and maximal oxygen consumption rates observed in the present study. It is a limitation of our study that we were not able to simultaneously measure ROS production and oxygen consumption rates to investigate this hypothesis. It is notable that we observed higher oxygen consumption despite our prior finding of lower mitochondrial mass. Studies with isolated mitochondria will be needed to differentiate the relative importance of mitochondrial mass, energy demand, coupled respiration, and ROS production for net cellular oxygen consumption in diabetes mellitus. In regard to mechanism, our finding that a CPT-1 inhibitor had similar effects in cells from both groups argues against an alteration in fatty acid utilization. Further study will be required to elucidate the mechanisms accounting for higher oxygen consumption and other mitochondrial abnormalities in diabetes mellitus. Overall, these studies in PBMCs are consistent with a large body of work suggesting altered mitochondrial respiratory capacity and function in other tissues, including skeletal muscle, liver, islet cells, and myocardium in diabetes.8,30,37 Since they are readily available, oxygen consumption in PBMCs may be a useful endpoint to study cross-sectional correlates and the effects of interventions on mitochondrial function in systemic diseases, such as type 2 diabetes mellitus.
Our study also adds to the prior literature by examining the relationship between oxygen consumption in circulating PBMCs and vascular function. We observed that patients with diabetes mellitus had markedly impaired vasodilator function of the conduit brachial artery and a trend toward blunted reactive hyperemia, findings that are consistent with many prior studies (see Tabit et al. for a review). 22 Despite the apparent systemic nature of mitochondrial dysfunction in diabetes and a large body of experimental work implicating these mechanisms in endothelial dysfunction, 5 we found no significant correlation between oxygen consumption rates in PBMCs and endothelial function. We did observe a significant correlation between maximal oxygen consumption and non-endothelium-dependent nitroglycerin-mediated dilation. It is known that mitochondrial ROS contribute to dysfunction of vascular smooth muscle cells in cardiovascular disease. 38 One might speculate that common mechanisms might account for altered oxygen consumption rates in PBMCs and vascular smooth muscle cells in diabetes, but investigation of this possibility goes far beyond the scope of the present study. A more likely explanation for the lack of correlation between oxygen consumption rates and endothelial function is the relatively high degree of variability associated with non-invasive vascular function measurements and the relatively modest sample size. Finally, these results emphasize the importance of direct examination of endothelial cells to understand mechanisms of endothelial dysfunction, and it is notable that such studies are now feasible in humans.13,15
Our study has a number of limitations. First, the sample size was relatively modest and we had limited power to adjust for potential confounders or show correlations between the measured variables. Second, the diabetic patients were taking medications that might influence mitochondrial function and confound the results, although we observed no difference in oxygen consumption rates in diabetic patients who were or were not taking a statin, ACEI/ARB, or metformin. Finally, further studies will be needed to elucidate the mechanisms that account for the findings of this initial report.
In regard to the oxygen consumption methodology, several additional points are worth noting. First, the approach requires study of freshly isolated, live cells, which would make it less useful than a test that could be performed using frozen samples. Second, we used an XF96 Seahorse analyzer and had only two ports available for addition of oligomycin and FCCP to the well. As a result, we could not test the effects of antimycin A and rotenone to distinguish the contributions of proton leak and non-mitochondrial respiration to basal oxygen consumption. We did not have access to an XF24 analyzer, which has additional ports and might be used in future studies; however, the larger well size in that device would require a larger number of cells. These limitations are balanced by the potential utility of this method for clinical research and clinical care because it provides for the measurement of a key aspect of mitochondrial function in readily accessible PBMCs.
In summary, we described a method to evaluate oxygen consumption in freshly isolated PBMCs and used it to examine mitochondrial function in patients with diabetes mellitus. We observed higher basal, maximal, and uncoupled oxygen consumption in the diabetic patients, findings that are consistent with prior work showing increased mitochondrial ROS production in PBMCs. Oxygen consumption in circulating blood cells did not correlate with endothelial vasodilator function, although we did observe a correlation between nitroglycerin-mediated dilation and maximal oxygen consumption that may merit further study. These findings add to our understanding of mitochondrial dysfunction in circulating leukocytes that may be relevant to inflammation, immune dysfunction and cardiovascular disease in diabetes mellitus. Furthermore, our described method might be applied to epidemiological studies and intervention studies, and might eventually prove useful for clinical care.
Footnotes
Acknowledgements
We gratefully acknowledge Seahorse Bioscience, Inc. for the loan of an XF96 analyzer and donation of plates to conduct the described experiments. We also acknowledge the technical assistance of Mai-Anne Carey and Jonathan Aasen with the non-invasive testing of vascular function.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
The project was supported by NIH grants HL081587, HL083801, HL083269, and HL115391 from the National Heart, Lung, and Blood Institute. Drs Vita and Hamburg receive support from the NIH-sponsored Boston University Medical Center Leadership Program in Vascular Medicine (K12 HL083781). Dr Fetterman receives support from T32 HL007224. Dr Hamburg is supported by NIH grant HL102299.