
Abstract
Endothelial microparticles (EMPs) belong to a family of extracellular vesicles that are dynamic, mobile, biological effectors capable of mediating vascular physiology and function. The release of EMPs can impart autocrine and paracrine effects on target cells through surface interaction, cellular fusion, and, possibly, the delivery of intra-vesicular cargo. A greater understanding of the formation, composition, and function of EMPs will broaden our understanding of endothelial communication and may expose new pathways amenable for therapeutic manipulation.
Introduction to extracellular vesicles
Microparticles are submicron vesicles generated from membranes via an exocytic budding process and transported into the circulation (Figure 1). They are irregularly shaped and range from approximately 100 nm to 1 µm in size. Their generation occurs via a range of stimuli leading ultimately to an increase in intracellular Ca++, loss of membrane asymmetry, membrane buckling and vesiculation from the parent cell and release into the extracellular environment (Figure 2). Microparticles are part of a family of extracellular vesicles, including exosomes and apoptotic bodies (Figure 3). Extracellular vesicles may be characterized according to size and range, from exosomes (30–100 nm), through microparticles (100–1000 nm) to apoptotic bodies (up to 4000 nm). Also, exosomes are derived from endoplasmic membranes and apoptotic bodies are derived from endoplasmic membranes. 1 This review focuses on endothelial microparticles (EMPs) and their role in vascular homeostasis and disease, and whether these sub-cellular agents are a therapeutic option for targeting and manipulation.
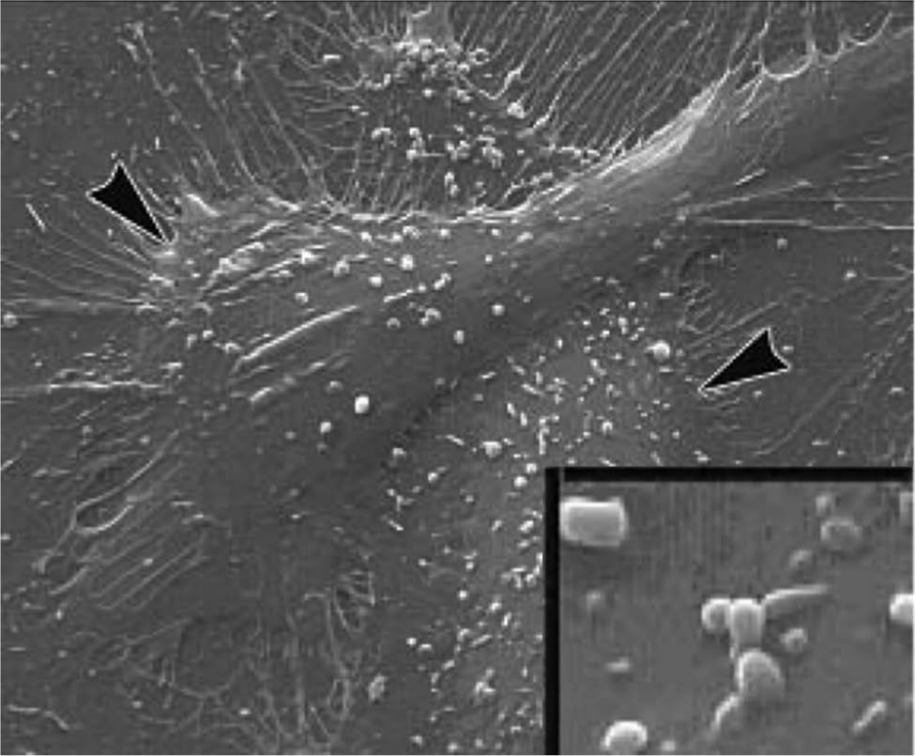
Scanning electron micrographs of cultured human aortic endothelial cells treated with tumor necrosis factor (TNF)α for 24 hours with microparticle formation (<1 µm) evident on the membrane surface. Arrowheads point to areas of microparticle formation.
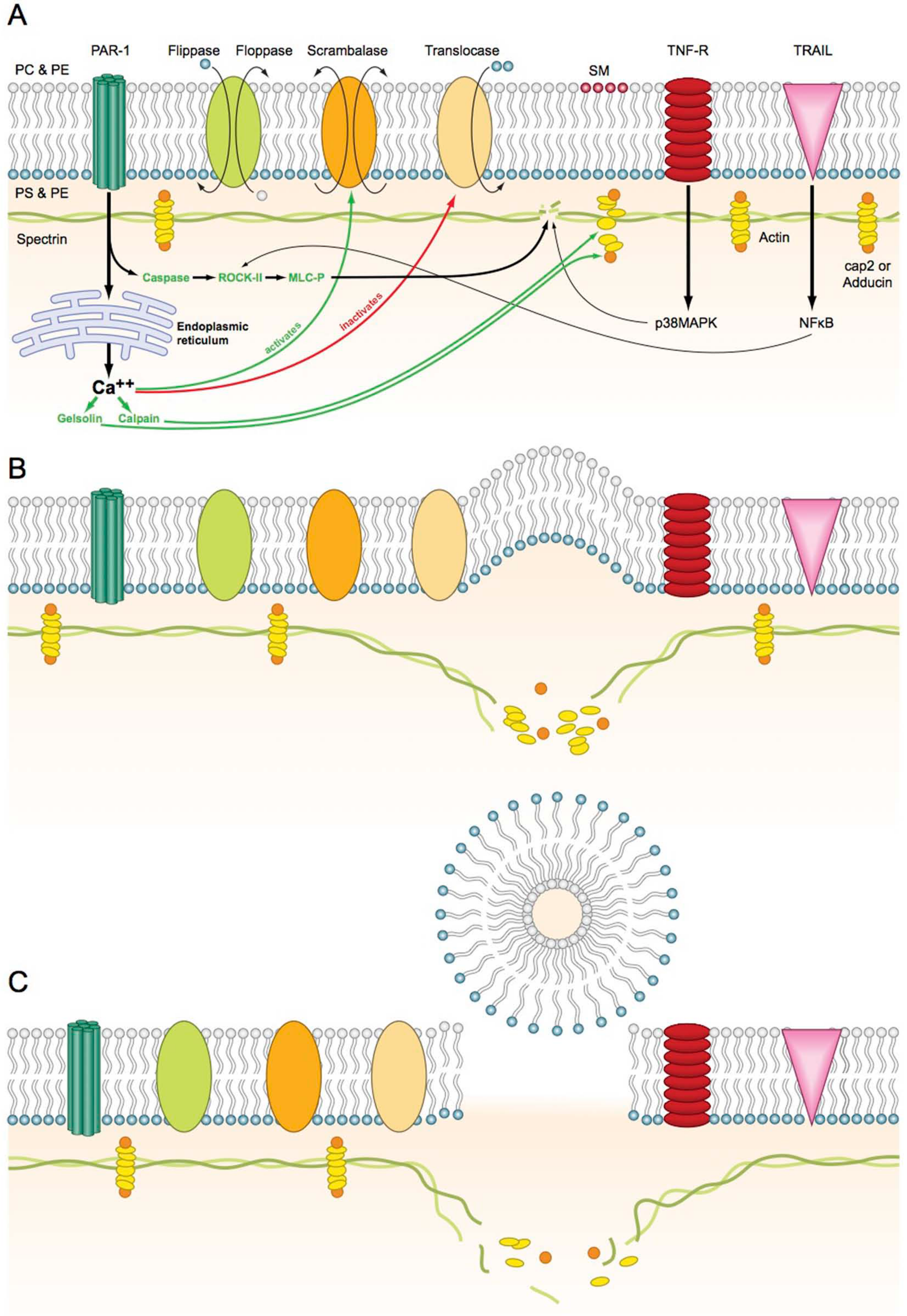
Schematic representation of cell membrane and microparticle formation. In the resting stage, scrambalase is inactive while flippase, floppase and translocase are active and maintain the membrane integrity. (A) Cellular activation: Triggers activating peroxisome proliferator-activated receptor (PPAR) cause release of calcium from the endoplasmic reticulum (ER) and activation of caspases. The caspases lead to activation of Rho kinase II (ROCK II) and eventually mitogen-activated protein (MAP) kinase and the calcium flux leads to activation of calpain, gelsolin and scrambalase, and inactivation of translocase. Activation of tumor necrosis factor receptor (TNF-R) causes the activation of p38 MAP kinase. Activation of TNF-related apoptosis-inducing ligand (TRAIL) by thrombin leads to activation of inflammatory transcription factor nuclear factor-kappa B (NF-κB), which ultimately activates ROCK II. (B) Cytoskeletal disruption: The activation of scrambalase and inactivation of translocase causes loss of membrane phospholipid asymmetry. The MAP kinases cause the disruption of spectrin, while calpain and gelsolin cause disruption of the actin in the cytoskeleton. (C) Microparticle generation: The cell membrane buckles outwards causing the formation of a vesicle that pinches off and is released into the extracellular space.
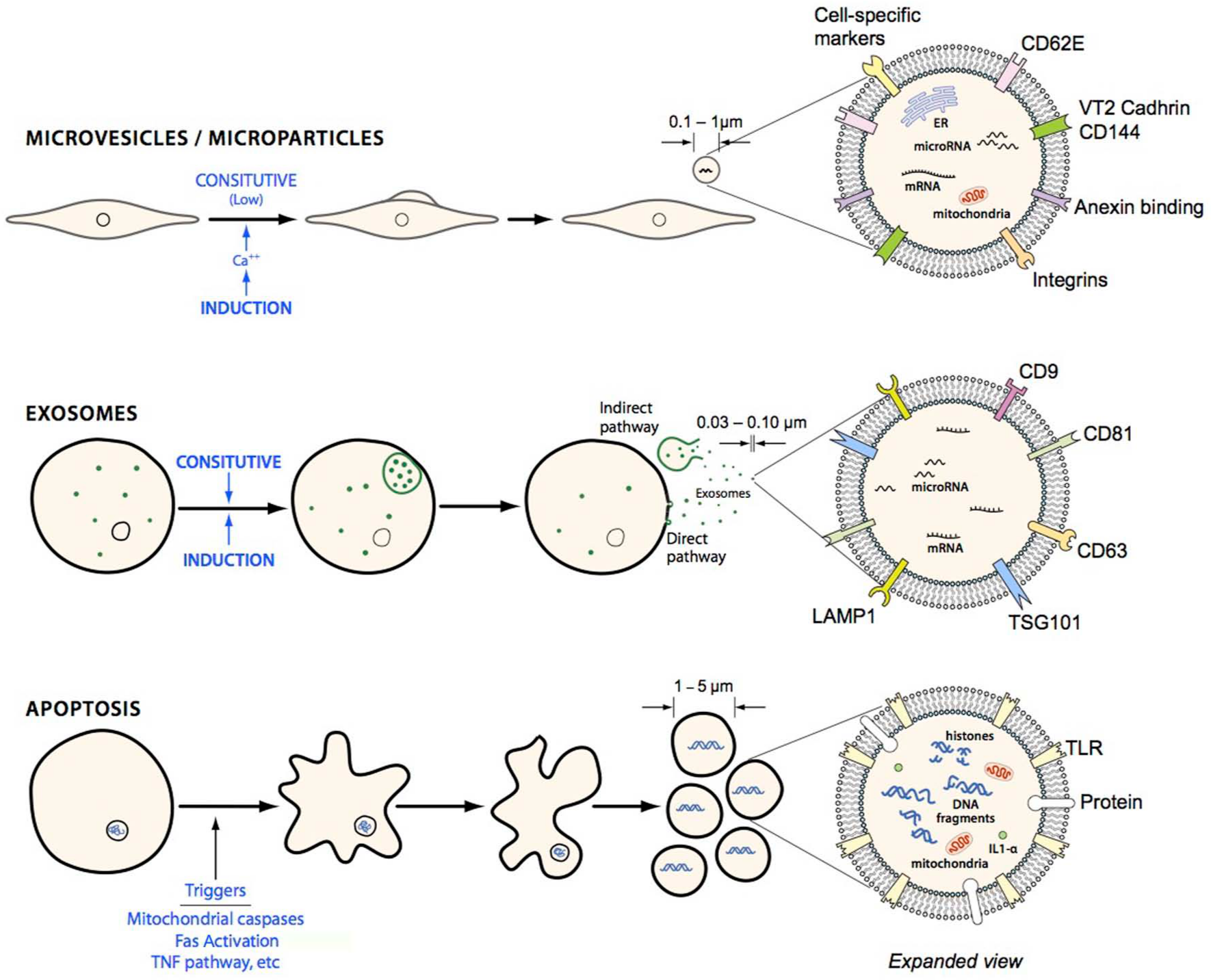
Schematic comparing different types of membrane vesicles: microparticles, exosomes and apoptotic bodies. Exosomes are sized in the approximate range of 30–100 nm and are thus smaller than microparticles and express surface markers such as LAMP1 and TSG101. Exosomes can either be generated from via a direct pathway from the membrane or mature in the cytosol into late endosomes and collect in multivesicular bodies (MVB) and become released via exocytosis. Apoptotic bodies are 1–5 µm in size and are released as cellular blebs as the cell undergoes apoptosis. Apoptotic bodies can be differentiated according to size and the presence of fragmented DNA.
The generation of endothelial microparticles
Biological factors pertinent to vascular health and hemostasis such as tumor necrosis factor alpha (TNFα), 2 thrombin, 3 plasminogen activator inhibitor-1 (PAI-1), 4 angiotensin II, 5 bacterial lipopolysaccharides (LPS), 6 uremic toxins, 7 arsenic trioxide 8 and modified lipids 9 can all induce EMP formation in vitro. Understanding the precise stimuli and associated mechanisms of EMP formation in vivo is still lacking. All hematopoietic cell types can release microparticles, with platelet microparticles being the most numerous, and when identified in the 1960s were termed ‘platelet dust’ and noted to have strong procoagulant capacity. In 1999, elevated numbers of microparticles were first observed in diseased populations, 2 and were thus considered as potential diagnostic and prognostic biomarkers. Barry et al. observed that platelet microparticles exposed to endothelial cells could induce de novo expression of COX-2 and prostacyclin (PGI2), and increase the adhesion of monocytes to endothelial cells,10,11 demonstrating a functional role for microparticles on the endothelium. The field has developed exponentially and today it is recognized that microparticle effects span coagulation, vascular tone, inflammation, angiogenesis, metastasis, regeneration and possibly even shape the innate and adaptive immune response. Microparticles hijack functional receptors, proteins, bioactive lipids and genetic material from the parental cell (Figure 3) and are active sensors, communicators and effectors of the intercellular environment. That microparticles are always formed under deleterious circumstances is perhaps an oversimplification. The challenge that lies ahead is to understand the impact on vascular health and disease of EMPs generated via distinct stimuli.
The biogenesis of EMPs
Enzymatic changes leading to vesiculation involve cytoskeletal re-organization with the inhibition of translocase and activation of scrambalase (Figure 2) and are driven by alterations in intracellular calcium leading to calpain activation and subsequent calcium-dependent proteolysis of cytoskeletal proteins. 12 However, the cell-specific intracellular pathways leading up to these membrane changes are not well defined. Pro-inflammatory, pro-apoptotic and procoagulation factors along with modified lipids and toxins all elicit EMP formation and release in vitro. A correlation exists between low shear stress and EMP number in end-stage renal disease patients, implicating the importance of hemodynamics. 13 Short pulses of second-hand smoke caused induction of EMPs along with repression of endothelial progenitor cells and nitric oxide (NO) in healthy subjects. 14 Thus, the increased generation of EMPs from an endothelium may be considered a hallmark event marking the beginning of endothelial dysfunction.
Evaluating the intracellular transcriptional events leading to EMP formation, microarray analysis on thrombin-stimulated microvascular endothelial cells showed enhancement of genes linked to cytoskeleton arrangement. 3 Cytoskeleton activation was also associated with microparticle formation in mesenchymal stem cells, blocking the formation of microfilaments with cytochalasin B and thus inhibiting their release. 15 In thrombin-stimulated microvascular endothelial cells, Rho kinase (ROCK) II but not ROCK I expression was enhanced, leading to enhanced cleavage by caspase-2, a critical step for thrombin-induced EMP formation. 3 Also, angiotensin II (Ang II)-stimulated EMP formation is through an NADPH and ROCK-dependent pathway. 5 This is provocative given that ROCK activation has been intensely studied in relation to arterial inflammation, remodeling and hypertension 16 and thus it is tempting to speculate that some clinical benefit of ROCK inhibitors may act partly via EMP inhibition. Disruption of lipid rafts by methyl-β-cyclodextrin (MBC) and nystatin abrogated the formation of EMPs, implicating lipid rafts as the membrane site of EMP formation. Thrombin activates the release of soluble TRAIL/ApoL2, a cytokine belonging to the TNFα superfamily. TRAIL signaling through the TRAIL/R2 complex mediates EMP release by downstream activation of the key inflammatory transcription factor nuclear factor-kappa B (NF-κB). The release of soluble TRAIL with thrombin activation is key to EMP formation as supernatants from thrombin-stimulated endothelial cells induced EMP release, whereas supernatants recovered from TRAIL knockdown endothelial cells did not. 17 This pathway eloquently demonstrates the often viscous cascade of inflammation given that EMPs acting on the endothelium induce further inflammation. EMP-mediated anti-inflammatory mechanisms might also be plausible given that p38 MAPK signaling is important for EMP formation, at least in vitro, 18 suggestive also of downstream NF-κB activation. The oral antiplatelet drug clopidogrel inhibits the formation of microparticles from cultures of human umbilical vein endothelial cells (HUVECs), through its action on p38 MAPKs. 19
Endothelial microparticles – a complex milieu of proteins, lipids and nucleic acids
Cell surface protein expression allows for the detection of circulating EMPs by flow cytometry using markers such as VE-cadherin (CD144) and E-selectin (CD62E) (Figure 3). Like many other microparticle subtypes, EMPs present an array of diverse molecules on their surface, with functions encompassing coagulation, cell survival, inflammation, enhanced oxidative stress, adhesion, proteolysis, remodeling, angiogenesis and tumor growth. 20 EMPs contain intracellular proteins, nucleic acids and organelles. A proteomics study of in vitro-derived EMPs reported that cytoplasmic, endoplasmic reticulum, mitochondrion and ribosomal proteins were all detected in significant numbers in these particles. 4 This informs us of the complex processes occurring within the cell ahead of formation of the particle at the surface membrane, and suggests that proteins from other organelles might be chaperoned to the site of microparticle synthesis. Once released into circulation, EMPs can then exert functional effects on cellular targets through surface membrane interactions and/or fusion and delivery of its intra-vesicular cargo. One of the first insights that extracellular vesicles might carry deliverable effective cargo came from a study on the effects of embryonic stem (ES) cell microparticles on the expansion of hematopoietic progenitor cells. 21 The addition of ES microparticles induced early pluripotent and hematopoietic markers along with activation of MAPK p42/44 and AKT. The microparticles were highly enriched for Wnt3 protein and messenger ribonucleic acid (mRNA) for several pluripotent transcription factors as compared to the parental ES cells, and remarkably, the microparticle mRNA was delivered and translated in the recipient hematopoietic cell. mRNA within endothelial progenitor cell (EPC) microparticles was shown to directly fuse with mature endothelial cells and deliver functional mRNA leading to enhanced angiogenesis. 22 The integrins (α4 and β1) on the microparticle surface allowed binding and incorporation within the endothelial cell. Although divergent sets of mRNAs were identified in the microparticles, four of the 78 transcripts were associated with phosphatidylinositol 3-kinase (PI3K)/AKT signaling, and they showed that PI3K and eNOS can both impact on the angiogenic effect of EPC microparticles.
The first evidence of non-coding microRNAs (miRNAs) in extracellular vesicles came from exosomes from human and mouse mast cells. 23 Small RNAs including miRNA were detected in exosomes and its transfer to target cells. Microparticles and exosomes isolated together from plasma of healthy individuals were shown to contain miRNA, although it is not clear from this study to what extent microparticles alone contributed to the miRNA collection. 24 Mesenchymal stem cell microparticles contain ribonucleoproteins involved in RNA storage, suggesting that they may shuttle RNAs from different cytoplasmic compartments including the nascent microparticle structures. 15 Hergenreider et al. demonstrated that endothelial cells expressing the atheroprotective transcription factor KLF2 or sheer flow can enhance the expression of miR-143 and miR-145 in released extracellular vesicles. These annexin V-negative vesicles, distinct from apoptotic bodies, are transferred to smooth muscle cell (SMC) where the miRNAs reduce expression of their targets and prevent SMC de-differentiation and attenuate atherosclerotic lesion formation. 25 Microparticles may therefore serve as biological vectors to shuttle nucleic acid material from one cell to another. That EMPs possess intra-vesicle nucleic acid content is intriguing and more so given the aforementioned data showing horizontal transfer of RNA to recipient cells with functional consequences. Further research is needed to understand which miRNAs are selectively packaged into microparticles under select conditions and how this RNA is transported, sensed and utilized in the targeted cells.
Therapeutic modulation of EMPs
Interpretation of microparticle results from pharmacology studies or human trials are hampered both by the complexity of the system and our ignorance of the influence of confounding variables such as gender, time of sampling, patient age, diet and underlying risk factors. On top of this is the direct effect of drugs on the contributing cell type. For example, platelet number contributes significantly to the levels of platelet microparticles and thus in an uncontrolled trial antiplatelet drug consumption may actually demonstrate a depression of microparticle number in patients with cardiovascular disease. Patients with coronary artery disease under an 8-week regimen of acetylsalicylate (aspirin) 100 mg per day, showed a marked decrease in platelet and EMPs. 26
The effect of HMG-CoA reductase inhibitors (statins) on EMP formation remains somewhat ambiguous. One of the first studies performed indicated that human coronary artery endothelial cell (HCAEC) microparticles derived via TNFα stimulation were sensitive to suppression by fluvastatin. 27 The drug’s ability to inhibit Rho kinase signaling was important. However, a further in vitro study yielded differing results. 28 Using HUVECs and simvastatin, these authors found that physiologically relevant concentrations of this statin increased apoptotic cell detachment and microparticle release. They found that simvastatin promotes microparticle release by repression of protein prenylation, possibly through caspase-8. The authors concluded that increased EMPs with statin treatment may reflect statins’ ability to improve the overall health of the endothelium by facilitating apoptotic cell detachment and microparticle release. A further study on patients with ischemic cardiomyopathy showed atorvastatin decreasing circulating EMPs and increasing EPCs and independent of the concomitant decrease in lipids, oxidized low-density lipoprotein (LDL) and high-sensitivity C-reactive protein (hsCRP). 29
EMPs as active players in vascular disorders
The suggestion that EMPs are causative agents in vascular pathology has arisen from their numerical increase in a range of vascular disorders. Many, if possibly not all, conditions associated with endothelial dysfunction have perturbations in EMPs. One of the primary cellular responses to an activated, damaged or compromised endothelium is vesicular release, placing EMPs as an early marker of vascular dysfunction. Not surprisingly, a range of clinical pathologies affecting the vessel wall, such as acute coronary syndromes, atherosclerosis, stroke, hypercholesterolemia, metabolic syndrome, obesity, diabetes, peripheral artery disease, hypertension and heart failure,30–36 display altered and often elevated numbers of EMPs. Large elevations in EMPs, observed in these conditions, certainly underscore their clinical relevance as a marker and predictor for vascular health and outcomes. The predictive power of EMPs was tested in a prospective study of 350 patients which showed that VE-cadherin (CD144+) EMPs independently predicted future cardiovascular events in high-risk patients (by virtue of established risk factors). 37 EMP counts (>353 events/µl) in untreated patients with pulmonary hypertension were associated with adverse clinical events. 38 Here, only CD62e+ (E-selectin) EMPs and not CD144+ or CD31+/41− were informative, demonstrating the selective dynamics of subsets of EMPs. EPCs contribute to vascular repair, and the balance of EPCs and microparticles is important for the integrity of vascular cells and endothelial function. Studies have shown that the ratio of procoagulant microparticles/EPCs is a useful index to determine vascular health in high-risk patients with underlying complications.33,39
Organ transplant monitoring might utilize EMPs as prognostic biomarkers. In heart transplants, procoagulant E-selectin (CD62E) bearing EMPs released during acute allograft rejection reflect endothelial cell activation and Fas-mediated apoptosis and were an independent marker acute allograft rejection. 40 The endothelium is one of the main targets for allograft rejection; therefore, monitoring and characterization of released EMPs into circulation may prove useful as a real-time monitoring tool of allograft function. 41 Further well-controlled prospective studies are required to establish firmly the prognostic value of EMPs in these conditions.
Notwithstanding their prognostic potential, EMPs modulate their surrounding environment. The endothelium is the site of orchestration of inflammation, coagulation, vasomotor activity, angiogenesis and vascular remodeling. EMPs can impact on all of these biological processes, possibly due to the diverse array of phenotypically distinct subsets of EMPs generated via selective pathways. This insight into molecular pathology differentiates EMPs from functional vascular health measurements, such as flow-mediated dilatation on ultrasound or acetylcholine infusion during angiography or biological markers like soluble proteins, which provide information on singular aspects of endothelial function. 42 The value of EMPs is that they may provide a comprehensive index of endothelial homeostasis, reflective of the parent cellular state. The biogenesis and biology of function on the endothelium are critical aspects. In vitro, selective stimuli and environments produce qualitatively and quantitatively unique subsets of EMPs,4,33,43 but how exactly this will translate to EMPs produced by the endothelium in healthy versus disease conditions remains largely unknown.
EMPs in coagulation and thrombosis
Knowledge of the role of EMPs in coagulation and thrombosis is frustrated by the lack of viable experimental approaches to dissect their contribution in vivo. Tissue factor (TF) is the major cellular activator for the extrinsic clotting cascade 44 and several in vitro studies find that certain agonists induce TF-positive EMPs.2,6,9 EMPs bound to monocytes induce the synthesis of tissue factor. 45 However, in healthy individuals, TF-positive EMPs are difficult to detect with 95% of circulating tissue factor from peripheral blood mononuclear cells and the remaining 5% from monocyte-derived microparticles. 46
Pawlinski et al. demonstrated that selective deletion of tissue factor from endothelial cells had no effect on activation of coagulation in a mouse model of endotoxemia. 47 Nevertheless, TF-positive EMPs have been detected in sickle cell patients, 48 leaving open the possibility that TF expression may be restricted to certain diseases and possibly to subsets of EMPs that have yet to be characterized. 49 The presence of anionic phospholipids, particularly phosphatidylserine (PS) on the surface of EMPs, contributes to its procoagulant potential. EMPs also act as a docking station for the assembly of components of the clotting cascade. This attraction occurs via the interaction with positively charged γ-carboxyglutamic acid (GLA) domains in some clotting proteins. 49
Endotoxemia and sepsis carry increased thrombotic risk including disseminated intravascular coagulation and lower levels of endothelial, platelet and leukocyte microparticles, which were associated with poorer outcomes and mortality in these conditions. 50 It appears that the microparticle pool, including those derived from endothelial cells, is necessary to prevent hypo-reactivity, and thus propagation of the shock. The microparticles therefore function in a cardio-protective capacity via delivery of thromboxane A2 (TxA2) to the vessel wall. 51 The in vitro interaction of EMPs with monocytes induced TF-dependent procoagulant activity from the monocytes. 45 This manifested through the interaction of EMPs by surface intercellular adhesion molecule 1 (ICAM-1) and its counter receptor β2 integrins on monocytes. Microparticles also harbor proteases such as TACE/ADAM17, on their surface, cleaving transmembrane proteins on target cell surfaces. This can induce shedding of endothelial protein C receptor (EPCR) on endothelial cells, enhancing in situ thrombogenicity. 52 In short, these findings illustrate the broad scope and multitude of mechanisms by EMP in hemostasis and their beneficial effects in certain pathological situations.
Are EMPs fueling the fire of inflammation?
Inflammatory mediators such as TNFα, interleukin (IL)-1β and thrombin enhance the generation of EMPs and these microparticles are believed to act in a paracrine fashion, promoting further vascular inflammation adding to the cyclical inflammatory milieu. 1 It is conceived that EMPs not only reflect the activation status of the cell but also confer further systemic inflammatory activity, implicating them as both a cause and consequence of inflammation. There is a direct correlation between EMP number and soluble IL-6 and CRP concentration in healthy young men, indicating that low-grade inflammation and EMP formation are correlated. 53 Certainly the increase in EMPs witnessed in many consensus inflammatory-type pathologies such as atherosclerosis, diabetes and autoimmune conditions such as multiple sclerosis and systemic lupus erythematosus (SLE) is suggestive that chronic inflammation drives the release of EMPs in vivo. What remains unanswered is whether these EMPs are active participants in the inflammatory feedback loop and if so how? In vitro EMPs induce inflammation in naïve endothelial cells by way of induction of ICAM-1 message and protein. 18 The direct transfer of ICAM-1 message and protein to the target cell is plausible, but receptor ligand interaction between the particle and cell, delivery of transcription factors or modulation by microvesicle miRNA might also be at play. Plaque microparticles harboring ICAM-1 were able to transfer this molecule directly onto endothelial cells, resulting in increased monocyte adherence. 54 Plaque microparticles did not affect other adhesion factors such as VCAM-1 and E-selectin or IL-6 and IL-8, indicating selectivity. EMPs injected intravenously into mice lead to increased systemic and pulmonary levels of IL-1β and TNFα, correlating with increased neutrophil recruitment to the lung. In relation to the secretion of IL-1β from cells, a cytokine which lacks a secretory signal peptide, microparticle shedding appears critical in the extracellular transport of IL-1β from ATP-stimulated macrophages. 55 Macrophage microparticles contain the inflammasome NALP3 and caspase-1, both required for IL-1β processing from cells. 56 Whether these properties are specific just for immune cell microparticles or extend to EMPs has yet to be determined. A study sought to understand if IL-1α, which is constitutively present in endothelial cells, is passaged out via microparticles. 57 Apoptotic bodies were distinguished from cell-derived microparticles via DAPI and histone staining and it was found that only apoptotic bodies contained IL-1α. Also, only apoptotic bodies and not microparticles induced chemokine production from HUVEC through the IL-1α pathway. As there are now better markers and methods to separate the different classes of extracellular vesicles, it is clear that further studies are needed to clearly delineate the separate contribution of apoptotic bodies, microparticles and exosomes in inflammatory pathways.
EMPs and angiogenesis
As discussed previously, microparticles from EPCs can have angiogenic properties. Angiogenesis is the formation of new capillaries from the pre-existing vasculature and is controlled through growth factors, such as vascular endothelial growth factor (VEGF) and signaling pathways (Figure 4). Microparticles from a number of different cell types impact on angiogenesis and this has been reviewed in detail elsewhere. 58 The angiogenic effect of microparticles is highly dependent on the surface expression of molecules on the microparticle. For example, it is reported that T-lymphocyte-derived microparticles (TMP) are pro-angiogenic through the increase of both endothelial cell adhesion molecules and expression of pro-angiogenic factors. 59 In contrast to this, one study showed that a population of TMPs has opposing effects, suppressing angiogenesis and endothelial cell viability through increased generation of reactive oxygen species. 60 The majority of studies suggest that EMPs inhibit angiogenesis; however, the underlying mechanisms by which this occurs remain unclear. The role of reactive oxygen species may also be important as in vitro-derived EMPs from HUVECs impair angiogenesis which can be restored with a cell-permeable superoxide dismutase (SOD) mimetic. 61
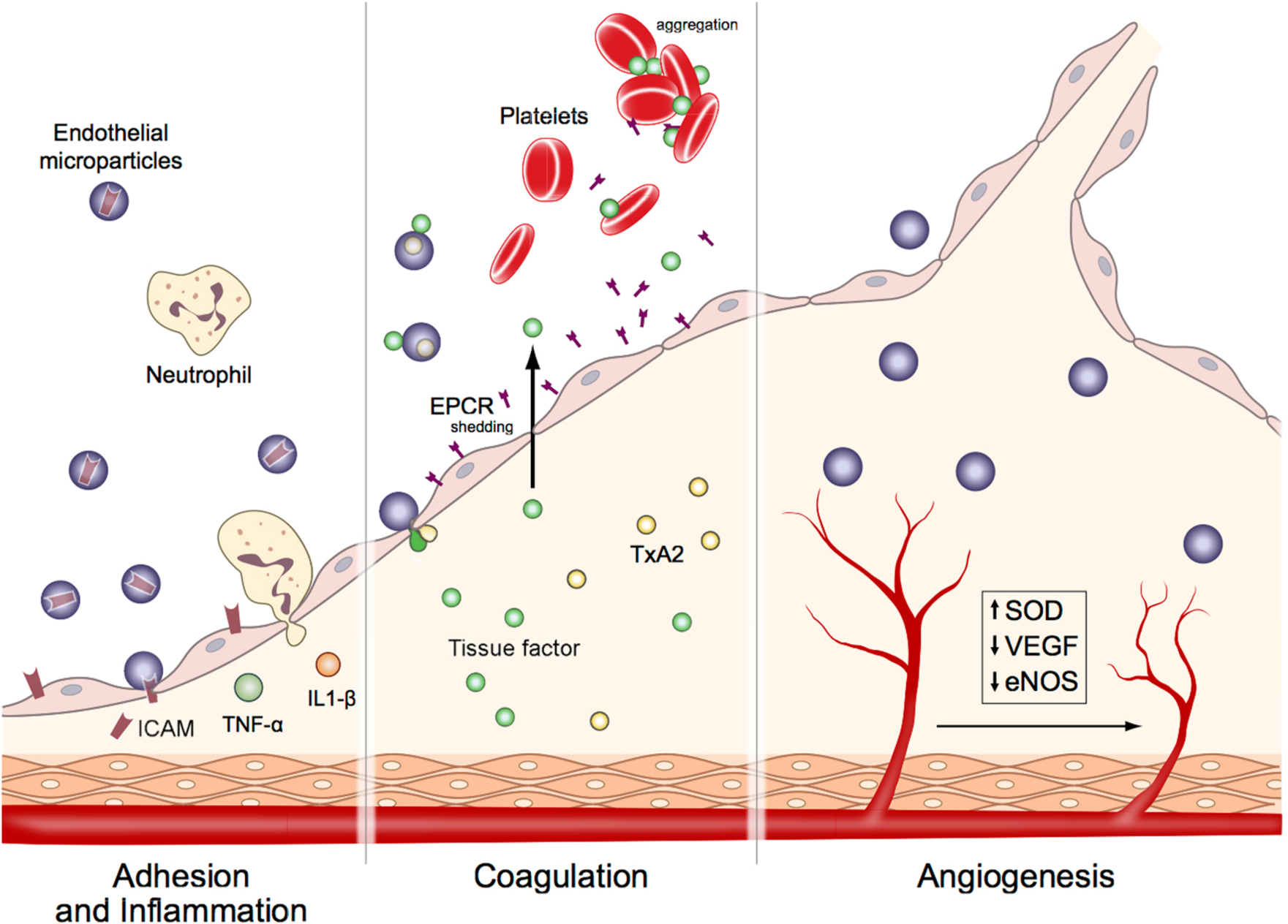
Schematic showing major effects of endothelial microparticles (EMPs) on cellular functions. (A) Adhesion and inflammation: EMPs transfer ICAM-1 to endothelial cells and cause activation of inflammatory cascade resulting in neutrophil chemotaxis and induction of TNFα and IL-1β. (B) Coagulation: EMPs induce the synthesis of tissue factor (TF), deliver thromboxane A2 (TxA2) and cause shedding of endothelial protein C receptor (EPCR), which result in initiation and amplification of thrombosis. (C) Angiogenesis: EMPs inhibit angiogenesis by down-regulating vascular endothelial growth factor (VEGF) and nitric oxide synthase (eNOS), and inducing superoxide dismutase (SOD) enzyme systems.
Microparticles from human microvascular endothelial cells (HMEC) were shown to serve as a surface for the generation of plasmin and EMP plasmin generation affected in vitro tube formation. This effect was entirely dose dependent as low concentrations of EMPs supported tube formation whereas high concentrations inhibited it. 62 Physiological and pathophysiological concentrations of in vitro-derived EMPs were injected into Ldlr−/− mice on a high-fat diet. 63 Both concentrations of EMPs inhibited angiogenesis in the hearts of these mice with effects on eNOS and NO generation. In patients with type 2 diabetes mellitus, circulating EMPs increase with disease severity. 33 A recent study analyzed the effect of plasma microparticles from both healthy and type 2 diabetic patients on endothelial tube formation. They observed that tube networks were longest and most stable from endothelial cells incubated with microparticles from healthy controls, whereas no tube formation occurred with microparticles from diabetic patients with coronary artery disease. 64 Overall, it would appear that circulating EMPs can inhibit angiogenesis, and this may be detrimental in conditions of failed angiogenesis such as diabetes mellitus. However, within an atherosclerotic plaque, their role may differ. Leroyer et al. demonstrated that CD40 ligand+ microparticles can enhance endothelial proliferation, promoting in vivo neovessel formation and favoring intraplaque hemorrhage. 65 Ligation of CD40 ligand with CD40-expressing endothelial cells appears to modulate VEGF and PI3K/AKT activation and cell proliferation. Subsets of plaque microparticles are of endothelial origin, with the majority of leukocyte origin. Similarly, although other subsets of microparticles have been classified in cancer, the specific role of EMPs in cancerous conditions has yet to be properly established. 66
Effect of EMPs on target cells and therapeutic potential
EMPs can exert functional effects on target cells through a number of currently known mechanisms. These mechanisms include surface membrane and ligand-receptor interactions, enzymatic cleavage of receptors on target cells by microparticle-born enzymes and proteases, fusion and importation of microparticle cargo into the target cell and also indirectly by providing a docking site or signal for other molecules or cells to interact with the target cell or tissue.
MPs can negatively impact on the endothelium by modifying NO production, bioavailability and signaling in the target cell. In acute lung injury (ALI), EMPs generated with human plasminogen activator inhibitor (PAI-1) could induce significant lung injury at physiological concentrations. 67 This was due to inhibition of endothelium-mediated vasodilation and NO generation from eNOS. Endothelial cell cultures co-incubated with EMPs showed a decrease in eNOS phosphorylation at Ser1179. The signaling by eNOS was also compromised in another study where increased phosphorylation of the inhibitory site Ser495 of eNOS was observed after incubation of endothelial cells with microparticles from patients with metabolic syndrome. 68 EMP formation by CRP is also dependent on NO. Wang et al. described how CRP-mediated induction of EMPs from HUVECs was related to impaired tetrahydrobiopterin (BH4)-dependent NO production. 69
Currently, a number of clinical trials are measuring microparticles either as a biomarker or direct effector of thrombosis, angiogenesis, tumor spread, endothelial dysfunction and inflammation (reviewed in ref. 70). Such results will reveal the potential of EMPs as clinical biomarkers and offer further insight into the potential of targeting these vesicles for therapeutic benefit. As discussed, a number of pharmacological agents can alter the number of circulating EMPs; however, caution should be exercised when interpreting these findings. Changes in circulating EMPs with a therapeutic agent may be due to a direct effect of the drug on EMP generation, an indirect effect due to the general improvement in vascular health or due to drug-induced enhanced clearance of the circulating microparticles.
A further avenue of therapeutic control may lie in the area of preventing the interaction of deleterious EMPs with the target tissue. A possible role for β3 integrin antagonists was utilized in preventing SMC microparticles from interacting with the endothelium in a model of mouse aorta vaso-reactivity. 71 This type of therapeutic mechanism could be explored specifically for the prevention of EMP interactions with a cell type of interest. Specific agents that target the generation of EMPs could also be of value, such as agents that inhibit calpain, Rho kinase activity or cytoskeleton disruption. However, not all subpopulations of EMPs are deleterious and there appears to be a fine balance between EMPs that may be helpful versus harmful. The ubiquitous nature of some of the signaling molecules involved and the possible redundancy in EMP generation might also make effective targeting of disease causing EMPs difficult.
A fruitful avenue may still lie in the ability of EMPs to act as autocrine and paracrine effectors in cell-to-cell communication. How EMPs communicate with target tissues could be used as a model to understand and interrogate a novel system of cell-to-cell communication. For example, Deregibus et al. investigated the mRNA content within EPC microparticles that elicited an angiogenic effect. 22 From this, they were able to package that RNA content into artificial vesicles using lipofectamine and elicit the same effect. In essence, new avenues and targets for therapeutic control may be realized through a better understanding of how these vesicles operate under homeostasis and disease.
Challenges for microparticle research
Although the field of microparticle research has advanced considerably in the last decade, this area of research is still fraught with a number of challenges. Many of the challenges are technical and another involves bioinformatics. There have been numerous efforts to standardize the enumeration of microparticles.48,72 Microparticles are close to the size limit of detection on even the most advanced flow cytometers, making it difficult to reliably enumerate microparticle subsets. Variability is introduced with differences in sample collection and handling methods, which hampers comparisons of results between studies. 73 Whether EMP generated in vitro can be compared to in vivo-generated EMPs is still unclear. Studies that analyze the effect of in vivo-derived microparticles isolated from plasma may be hampered due by pre-analytic alterations introduced by the collection method. 74 The amount of data generated from flow cytometry measurement of microparticle subsets is extremely large and unbiased bioinformatics methods such as the newly applied cytometric fingerprinting 75 are needed for reproducible and relatively rapid generation of results.
Conclusions
There is now accumulating evidence on the multifaceted biology of EMPs as active intercellular communicators. These vesicles are no longer considered as passive products but active players regulating the intracellular environment. The dysregulation of EMPs may be an early marker of vascular dysfunction and as such provide useful markers of endothelial dysfunction. It is hoped that addressing cardiovascular risk factors will favorably modify production of proatherogenic microparticles and improved survival. The current clinical trials that are ongoing will provide the needed clarification as to how informative these vesicles are in large-scale multicenter clinical trials. The multiple roles of EMPs make interpretation of in vitro studies difficult. The function of EMPs is highly dependent on the stimulus used to produce the EMPs, the concentration of EMPs and the target tissue analyzed. Although this area is technically challenging, a fuller understanding of these vesicles, their formation, effect and role in physiology and deregulation with disease is required. This research will undoubtedly uncover new areas of biology and provide novel insights into the world of intracellular communication.
Footnotes
Declaration of conflicting interest
Drs Mohler, Rogers and Moore are founders of a company called Cytovas that is commercializing an assay that in part measures microparticles.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.