
Abstract
Recent studies on the anatomical and functional organization of GABAergic networks in central auditory circuits of the zebra finch have highlighted the strong impact of inhibitory mechanisms on both the central encoding and processing of acoustic information in a vocal learning species. Most of this work has focused on the caudomedial nidopallium (NCM), a forebrain area postulated to be the songbird analogue of the mammalian auditory association cortex. NCM houses neurons with selective responses to conspecific songs and is a site thought to house auditory memories required for vocal learning and, likely, individual identification. Here we review our recent work on the anatomical distribution of GABAergic cells in NCM, their engagement in response to song and the roles for inhibitory transmission in the physiology of NCM at rest and during the processing of natural communication signals. GABAergic cells are highly abundant in the songbird auditory forebrain and account for nearly half of the overall neuronal population in NCM with a large fraction of these neurons activated by song in freely-behaving animals. GABAergic synapses provide considerable local, tonic inhibition to NCM neurons at rest and, during sound processing, may contain the spread of excitation away from un-activated or quiescent parts of the network. Finally, we review our work showing that GABAA-mediated inhibition directly regulates the temporal organization of song-driven responses in awake songbirds, and appears to enhance the reliability of auditory encoding in NCM.
Introduction
Vocal communication is fundamental to the biology of social vertebrate species, including humans, and hinges both on the adequate production of communicative sound by the sender, and the reliable processing and interpretation of such signals by the receiver. In this review we focus on the brain mechanisms supporting the auditory processing of communication signals, or the receiver end. In particular, we review how GABA, the main inhibitory neurotransmitter in the vertebrate brain, shapes the auditory processing of behaviorally-relevant communication signals, and how GABAergic circuits are anatomically and functionally organized in the avian auditory telencephalon.
Significant advances have been made to our understanding of how learned communication signals are generated and processed by the vertebrate brain. A major fraction of these gains have come from neurobiological studies conducted in songbirds (for reviews, see 1 ). Of the songbirds studied, zebra finches have been the most extensively used species to elucidate the neurobiology of vocal learning, a behavior thought to constitute a substrate for speech acquisition in humans. 1 3 Vocal learning is a rare behavior only found in three groups of mammals (humans, cetaceans and some species of bats) and three avian groups (songbirds, parrots and hummingbirds).1,3 Importantly, this behavior relies on intact auditory processing. Either disruptions in auditory feedback or deafening during a critical period early in development impair the bird's ability to learn their vocalizations from a tutor. Furthermore, intact hearing continues to be important in adulthood as interference with adequate auditory processing leads to a gradual deterioration of learned vocalizations (songs).1,4–9 In addition to learning one's song, the auditory processing of songs in adults is necessary for a series of social behaviors in songbirds. Specifically, ethological studies have revealed that song is used to identify individuals within a flock, to select or assess the fitness of a potential mate, and during establishment and competition for territory.1,10–17 Even though the processing of auditory information is central to a wide array of adult behaviors in songbirds, little is known about the brain mechanisms underlying the auditory and perceptual processing of songs. In fact, a clear understanding of how the adult vertebrate brain encodes and decodes complex, natural communication signals has been one of the greatest challenges in contemporary neurobiology. To systematically and completely address this topic, it is necessary to carefully determine the anatomical, molecular, neurochemical and functional organizations of central auditory circuits. These areas of inquiry have been pursued by a number of research groups, in a variety of experimental models, and have been described elsewhere. 18 25 We will, therefore, not provide a detailed account of these findings here. Instead, we will summarize and interpret recent gains made by our group on the functional organization and contributions of inhibitory transmission in higher order centers of the songbird auditory telencephalon.
GABAergic Transmission as a Central Regulator of Sensory Processing
Neurons that release GABA were once believed to act on the cortex broadly, by controlling inhibitory tone and, therefore, were regarded as a global and/or region specific dampening mechanism for all on-going neural activity. Subsequent work clearly demonstrated, however, that sensory neurons exhibit spatially overlapping excitatory receptive fields (RFs), as well as inhibitory RFs, that compete for functional expression in response to an adequate sensory stimulus. 26 32 It is now very clear that inhibitory mechanisms are activated in parallel with excitation as part of sensory processing and shape neuronal tuning in at least spatial or spectral, as well as temporal domains.30,31,33–39 Computer-based modeling has also revealed that the balance of inhibition and excitation to a large extent determine the dynamic range of individual sensory neurons.40,41 When inhibitory inputs are active, cells enter more functionally rigid states and, when inhibition is less prevalent, neurons are more amenable to undergoing functional reorganization in response to altered sensory drive.42,43 Together these findings have been useful to propose that GABAergic inhibition may play an important role in countering or gating the malleability of central circuits in response to strong excitatory sensory drive. 44
The Ascending Auditory Pathway
The basic anatomical organization of the songbird central auditory pathways is similar to vertebrates in general, and has been carefully detailed previously.20,22,45–49 In the ascending system, auditory information is conveyed from the cochlea to telencephalic centers through a series of pontine, mesencephalic and thalamic structures20,48,49 (Fig. 1A). Thalamic auditory projections target the thalamo-recipient zone Field L2 in the telencephalon which, in turn, primarily project to the caudomedial nidopallium (NCM) and the caudomedial and caudolateral mesopallium (CMM and CLM, respectively)20,50 (Fig. 1B). While contentious, it has been proposed that the songbird Field L2 may be analogous to layer 4 of the primary auditory cortex (A1) in mammals.22,51 NCM and CMM, on the other hand, have been proposed to constitute structures that may be analogous to the supragranular layers of A1, based on anatomical connectivity patterns, or auditory association cortex, based on functional properties.22,50,51 Despite ambiguities on the degree of homology and/or analogy between the songbird and mammalian auditory forebrain structures, it is thought that in both groups such areas are involved in the auditory and perceptual processing of natural communication signals, with each area distinctively contributing to these processes. Of particular interest to our laboratory is NCM, an auditory forebrain area that has selective responses to conspecific (same-species) songs, exhibits forms of experience-dependent plasticity that putatively enable auditory learning, and is a site involved in the formation of auditory memories required for vocal learning. 50 57 Although the focus of this review will be on NCM, we will occasionally refer to findings obtained in the thalamo-recipient Field L, and the main output of NCM, the CMM.

GABAergic Neurons in the Auditory Forebrain
Although it is clear that GABA plays a central role in shaping sensory responses in all vertebrate species studied to date, the organization of GABAergic circuits in the auditory forebrain, and their contributions to the auditory processing of songs in songbirds had not been systematically explored until recently. We review below how our research has focused on determining the anatomical organization of GABAergic circuits, how inhibitory neurons are engaged as a function of auditory experience, and the roles of GABAergic transmission to regulating synaptic physiology and song-driven responses in the auditory forebrain.
Our first efforts centered on determining the distribution and organization of GABAergic neurons in the auditory telencephalon (Fig. 2). To this end, we utilized two independent approaches that reliably allowed for the identification of GABAergic neurons in the zebra finch brain. First, we cloned the zebra finch homologue of the 65 kDa glutamic-acid decarboxylase gene (zGAD65), one of the synthetic enzymes for GABA, and utilized this cDNA to generate zebra finch-specific riboprobes for in-situ hybridization 58 (Figs. 2A–C, G–H). By definition, neurons expressing zGAD65 mRNA are GABAergic. Second, we identified and characterized an antibody that specifically recognizes GABA in zebra finch tissue 49 (Figs. 2D–F, I–J). Importantly, most results obtained with in-situ hybridization for zGAD65 yielded results that were virtually identical to those obtained with immunocytochemistry directed at GABA.

Within all subfields of the thalamo-recipient Field L, GABAergic neurons were detectable at high densities, amounting approximately 39, 54 and 45% of the overall neuronal population, for subfields L1, L2a and L3,49,58 suggesting that inhibition may heavily shape thalamo-“cortical” synaptic physiology. Although GABAergic neurons were homogeneously distributed throughout these sub-structures, identified cells differed significantly in terms of diameter (i.e. from 4 to 22 μm). Cells tended to be slightly larger in L2a (13.2 μm average diameter) than in L1 and L3, (9.4 and 10.8 μm average diameter, respectively). 49 Importantly, Field L neurons were significantly larger than those in NCM and CMM, suggesting that inhibitory neurons may play different functional roles in these areas (see below) 58 (Figs. 2A–C, G).
GABAergic neurons were also highly prevalent in NCM, an auditory forebrain area that receives direct input from Field L and has extensive reciprocal connections with CMM49,58 (Fig. 2A). Interestingly, we found that GABAergic neurons are the prevalent cell type in NCM, accounting for nearly 52% of the overall neuronal population. 59 These findings suggested that inhibitory transmission is central to the physiology of NCM. Importantly, GABAergic cells in NCM could be qualitatively discriminated into two apparent groups (Figs. 2G–J). The first category encompassed GABAergic neurons with small diameters (3.3–10 μm), typically attributed to local circuit inhibitory neurons found in vertebrate sensory cortices. Importantly, this cell type accounted for the large majority of inhibitory neurons within NCM49,58 (Fig. 2A). The second category consisted of uncommonly large GABA-positive neurons, which exhibited diameters of 15–20.8 μm and that were only occasionally found in NCM49,58 (Fig. 2A).
Like NCM, the CMM was also found to be heavily endowed with GABA-labeled cells that comprise a significant fraction of the overall neuronal population (33.8%)49,58 (Figs. 2C, E–F). GABAergic neurons were distributed throughout this structure but showed a strong tendency to appear grouped into clusters of 3–5 cells, a pattern that is characteristic of this area. 49 Most CMM inhibitory neurons tended to be small in diameter (4 μm–12 μm), although some larger neurons (∼15 μm in diameter) were also detectable. Thus, apart from the presence of clusters of GABAergic neurons, the distribution and morphological properties of inhibitory cells in CMM were highly similar to those observed in NCM.
Interestingly, these less numerous large GABAergic neurons detected in NCM and CMM were highly comparable to the inhibitory cell population in Field L2a, which was primarily composed of large GABA-positive neurons 58 (Figs. 2A–B). These qualitative differences raise the intriguing possibility that Field L2a GABAergic cells may be projection neurons that target NCM and/or CMM, while the primarily small diameter population of inhibitory cells in NCM and CMM may participate in local circuit processes. Such a possibility remains to be explored in future studies.
GABAergic Neurons in NCM are Directly Activated by Birdsong Auditory Stimulation in Freely-Behaving Songbirds
Several aspects of the functional organization of the songbird auditory forebrain were uncovered with the use of activity-regulated gene expression. In this regard, the immediate early gene zenk (a.k.a. zif268, egr-1, NGFI-A, krox-24) has been extensively used to map song-responsive neurons in the songbird auditory system50,51,60 The gene zenk encodes a zinc-finger transcriptional regulator (ZENK) that is highly sensitive to neuronal depolarization and that has been repeatedly implicated in central nervous system plasticity. 61 66 Importantly, the mapping of zenk mRNA or protein has been successfully used to reveal song-driven neurons, at single cell resolution, in freely-behaving animals.60,67–70
We recently developed a double-fluorescence in-situ hybridization protocol that enabled us to detect two mRNA species at single cell resolution.58,69,71 This approach enabled us to investigate if inhibitory neurons are directly activated by auditory experience in the auditory forebrain of freely-behaving birds (Fig. 3). To this end, we used riboprobes directed at zenk mRNA to map song-responsive neurons, and a zGAD65 riboprobe to reliably label GABAergic neurons. 58 This method enabled us to conclusively determine that GABAergic cells are directly activated in the auditory forebrain of song-stimulated songbirds (Fig. 3). Specifically, it was found that approximately 42% of song-driven neurons in NCM, as revealed by zenk expression, were GABAergic. 58 Similarly, in CMM, nearly 34% of hearing-activated neurons expressed zGAD65 and, therefore, were by definition inhibitory neurons. 58 These findings suggest that GABAergic neurons directly, and robustly, contribute to NCM's and CMM's response to song in awake-behaving songbirds. Moreover, these observations are consistent with a prevalent role for inhibitory transmission in the auditory processing of behaviorally-relevant vocal communication signals.

Spontaneously Active GABAergic Synapses Regulate the Excitability of NCM's Excitatory Circuit
Our anatomical and molecular biological findings indicate not only that GABAergic neurons are prevalent in the songbird auditory forebrain, but also that they are major contributors to its response to song stimulation. To explore the role of inhibitory transmission in the physiology of auditory circuits, we focused on NCM. Specifically, we generated a slice preparation of NCM where whole-cell patch-clamp recordings could be obtained.58,59
Using this slice preparation we observed that NCM neurons receive high levels of spontaneous synaptic activity. Specifically, a high rate of spontaneous post-synaptic events were detected throughout NCM (average: 3.1 Hz)
59
(Fig. 4A, top panel). Importantly, the large majority (∼90%) of these events were inhibitory, and specifically carried out via type-A GABAergic receptors, as they were largely abolished by bath application of bicuculline (BIC), a selective GABAA receptor antagonist58,59 (Fig. 4A, bottom panel; Fig. 4C). These pharmacologically-isolated spontaneous inhibitory post-synaptic currents (sIPSCs) were observed in nearly all cells recorded, exhibited current magnitudes that averaged 48 pA,
59
and an average decay time constant (τ) of 6.9 ms
58
(Fig. 4B). Such parameters are in close agreement with previously reported decay time constants for GABAA receptors in a variety of preparations.72,73

Interestingly, blockade of these highly prevalent, spontaneously active GABAA-mediated synaptic events with BIC led to the appearance of large bursts of synaptic activity (average: 232 pA) that occurred at very low frequency (average: 0.08 Hz) 59 (Fig. 4D). These events were supra-threshold, as they could elicit action potential generation in current-clamp recording mode (Fig. 4E), and were carried out via non-NMDA glutamatergic receptors as they could be completely blocked by application of DNQX to the recording bath. 59 Together, these data suggest that spontaneously active GABAergic synapses are ubiquitously found in NCM and function to suppress the activity of NCM's glutamatergic circuitry at rest, perhaps to protect this neural circuit against runaway excitation.
Song-Driven Responses are Shaped by Complex Interactions among Inhibitory Circuits in Awake Songbirds
We explored the functional role of GABA during the auditory processing of songs by combining bilateral, multi-electrode extracellular recordings and local pharmacological manipulations in the NCM of adult, restrained zebra finches that were awake. Prior to unilateral infusions of BIC, song-driven neurophysiological responses were detected across most electrodes, and consisted of sustained/tonic spiking activity in response to song syllables (Fig. 5A, blue traces). These responses typically spanned through the inter-syllable intervals and continued after stimulus offset.59,74 During unilateral BIC application, significant changes in the temporal pattern of song-evoked responses were detected. Specifically, stereotypical tonic/sustained NCM neuronal responses were instantaneously converted into highly phasic, on/off-like responses (Fig. 5A). Importantly, the song-evoked response amplitudes (i.e. total voltage change integrated over the period of the played song plus residual firing, minus baseline activity) did not change (Fig. 5B). However, the variability of song-evoked responses increased following BIC treatment, refecting a shift from a tonic-dominant to a primarily phasic response profile (Fig. 5C). Finally, BIC significantly decreased the responsiveness of NCM neurons to certain song syllables. In fact, under BIC treatment, NCM neurons rarely responded to all song elements, unlike observations made under control (pre-drug) conditions
59
(Fig. 5A, grey box). Together, these findings indicated that 1) BIC induced a marked and unexpected temporal reorganization of song-evoked responses in NCM neurons. Unexpected because blockade of inhibition markedly suppresses responses to some song elements; 2) pharmacological disinhibition did not result in dramatically lowered thresholds for neuronal firing as no increase in overall spiking activity was observed, suggesting that BIC did not affect the number of neurons exhibiting responses to song, but rather impacted the temporal responses of previously active NCM neurons; 3) Blockade of GABAA-receptors with BIC decreased the reliability with which song elements (syllables) were encoded in spike trains of NCM neurons. Importantly, this decrease in the encoding fidelity of songs during BIC treatment could not be explained by a decreased ability of NCM neurons to discriminate frequencies, as frequency tuning curves were unaffected by local blockade of GABAA receptors in NCM.
59
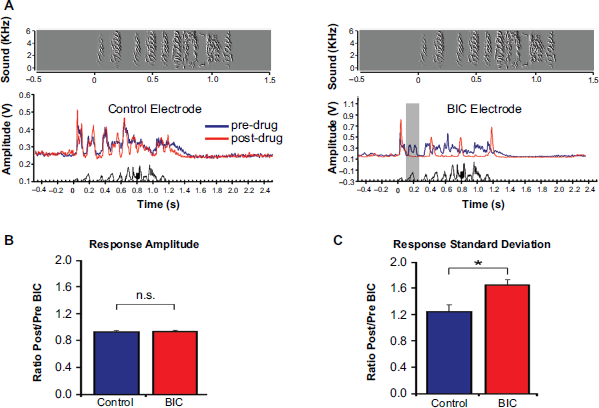
The results described above revealed an unconventional synaptic organization in the songbird NCM, whereby blockade of local inhibition led to an unanticipated suppression (or inhibition) of evoked responses to certain song elements. Although future studies will be required to fully determine the synaptic and neurochemical organization of NCM, our findings indicate that song-driven disinhibitory circuits may be at play. Such a mechanism would be consistent with our patch-clamp findings indicating that spontaneously active GABAergic synapses suppress the NCM's excitatory circuit. Although speculative, one possibility is that such spontaneously active synapses may be suppressed by an independent inhibitory circuit, perhaps GABAA-independent, during song stimulation, consistent with our extracellular electrophysiological findings (Fig. 6). For a full discussion on this topic and potential alternative mechanisms, see.
59

Concluding Remarks
Songbirds rely on the auditory processing of complex, natural communication signals for an array of behaviors, including those that involve individual recognition, as well as for song learning and the maintenance of the learned, stereotyped adult songs. Appropriately tuned auditory responses along the auditory pathway are postulated as a requirement for the optimal auditory and perceptual processing of acoustic features of songs. The findings presented in this review suggest that the highly prevalent GABAergic circuitry in the songbird central auditory pathways may play a central role in regulating both neuronal excitability and the auditory coding of birdsong. Future studies will focus on further exploring the contributions of GABAA-mediated inhibition to the auditory processing of songs in awake songbirds, elucidating the functional organization of inhibition carried out by other GABAergic receptors, and revealing potential interactions between GABAergic networks during the processing of behaviorally-relevant acoustic cues.
Disclosures
The authors report no conflicts of interest.
Footnotes
Acknowledgements
The work described here was supported by NIH/NIDCD, and University of Rochester start-up funds. We thank Dr. C. Mello for insightful discussions on the materials presented here.