
Abstract
Commonly used neuroimaging approaches in humans exploit hemodynamic or metabolic indicators of brain function. However, fundamental gaps remain in our ability to relate such hemo-metabolic reactivity to neurotransmission, with recent reports providing paradoxical information regarding the relationship among basal perfusion, functional imaging contrast, and neurotransmission in awake humans. Here, sequential magnetic resonance spectroscopy (MRS) measurements of the primary inhibitory neurotransmitter,
INTRODUCTION
Magnetic resonance imaging (MRI) has been widely applied to obtain a more thorough understanding of the central nervous system. In addition to being an excellent tool for structural tissue characterization, functional MRI (fMRI) can be used to measure regional and global cerebral hemodynamics and to make inferences regarding neuronal activity. Functional MRI, which most commonly exploits blood oxygenation level-dependent (BOLD) contrast, 1 does not require exogenous contrast agents and therefore is an invaluable tool for obtaining repeated functional measurements, assessing longitudinal changes in brain function, and for probing pathophysiologic mechanisms in clinical scenarios where contrast agents may be contraindicated. As a result, BOLD fMRI has emerged as the most popular tool for assessing brain function in humans, with a wide variety of clinical, pharmacological, and neuroscience applications to date.
However, the BOLD fMRI signal is only an indirect marker of neuronal activity, arising from complex neurochemical, metabolic, and hemodynamic modulations that occur concurrent to both ongoing and stimulus-evoked neuronal activity. Specifically, in stimulus-evoked experiments, BOLD contrast arises owing to a greater increase in cerebral blood flow (CBF; 20% to 50%) relative to cerebral blood volume (CBV; 5% to 30%) and the cerebral metabolic rate of oxygen consumption (0% to 20%), leading to an increase in diamagnetic oxy-hemoglobin relative to paramagnetic deoxy-hemoglobin in capillaries and veins.
While much progress has been made in understanding the hemodynamic and metabolic contributions to BOLD contrast, important gaps remain in our ability to relate BOLD signals to underlying neuronal activity and neurotransmission. These gaps significantly hinder BOLD interpretability in many applications, as substantial variability in BOLD responses exists between even healthy individuals, partly accounting for why BOLD data remain largely qualitative in nature. Understanding the physiologic sources of this variability is fundamental to using BOLD as a tool for identifying quantitative differences in brain function between individuals and conditions and for gauging functional response to disease and treatment.
The critical barrier to characterizing BOLD signal is that contrast is fueled by a variety of sources, with few direct observables. Alternative fMRI approaches specifically sensitive to individual hemodynamic parameters (e.g., CBF or CBV) can be applied in sequence with BOLD fMRI for more comprehensive investigations.2, 3, 4, 5, 6 Yet, the majority of multimodal studies in humans have focused on understanding hemodynamic and metabolic changes, with comparatively less information available on the neurochemical precursors to these changes. However, coordination between excitatory and inhibitory neurotransmission, facilitated by glutamate and
Much elegant work, primarily from the animal literature, has provided important insights into the neuro-energetic mechanisms of excitatory and inhibitory processes.
7
For instance, glucose taken up from the blood is converted into glutamate in neurons (glutamate oxidation; CMRglc(ox)), after which glutamate binds to postsynaptic neurons, and subsequently is converted into glutamine in astrocytes, and finally transported back to neurons (rate of glutamate/glutamine cycling:
To relate these findings to functional imaging, it is also necessary to understand how CBF varies with excitation and inhibition. While there is general agreement that changes in CBF correlate with oxygen usage during periods of elevated neuronal activity, 19 increases in CBF have been reported to extend beyond the locus of oxygen utilization, 20 which suggest that CBF is controlled by additional factors beyond signals related to energy production (e.g., CO2, H+) alone. Evidence has been provided for glutamate-evoked Ca2+ influx in postsynaptic neurons activating the production of nitric oxide, arachidonic acid, and adenosine, which in turn produce vasodilation and related increases in CBF.21, 22 Largely owing to less available data, more controversy exists with regard to how inhibition contributes to brain energy use, CBF, and thus functional imaging signals. In cortex, exogenous GABA has been shown to increase CBF and CBV, 23 yet this observation was not replicated in cerebellum. 24 It has also been reported that inhibition does not contribute to BOLD signals, partly because the fraction of inhibitory cells is smaller relative to the fraction of excitatory cells 25 and also because mouse astrocytes exposed to glutamate initiate uptake of glucose, whereas no uptake is observed during GABA exposure. 26 Therefore, despite elegant work on the neurochemical sequelae of energy production, it remains unclear how some of these findings translate to large-scale changes in CBF, metabolism, and therefore functional imaging signals, in a relatively large (e.g., 3 × 3 × 3 mm) fMRI voxel.27, 28
An additional complication is that neurotransmitter investigations are frequently conducted under pharmacological manipulation in sedated animals,8, 29 or with tissue biopsy or autopsy procedures, with the obvious caveat that
To gain additional information in awake humans, it is possible to measure
Unexpectedly, a positive trend has been reported between cerebral blood flow-weighted reactivity using pulsed arterial spin labeling (ASL) and baseline GABA concentration in visual cortex. 35 In a separate study using a different ASL technique, 38 no relationship between CBF and GABA was observed, despite clear trends between temporal features of the evoked BOLD time course and GABA. A lack of relationship between CBF and GABA is unlikely given the reproducible correlations between BOLD and GABA reported. Importantly, a lack of relationship between these parameters casts doubt on the relevance of the BOLD/GABA correlations outlined above (which are frequently reported in relatively small volunteer groups). Alternatively, demonstration of a relationship between GABA and CBF would underscore the relevance of these relationships, and furthermore provide a greater degree of quantitative interpretability.
Here, we aim to better characterize these relationships by performing careful measurements of CBF and GABA in human occipital lobe using improved experimental and statistical methods that account for variations in blood velocity and voxel tissue fraction. Importantly, ASL measurements are sensitive to both CBF and arterial arrival time (AAT: time for the labeled blood water to reach the capillary exchange site). Therefore, AAT, not CBF, may positively correlate with GABA and this relationship could explain the ASL–GABA correlation. This would provide support for blood water velocities, likely at the arteriolar level, being associated with GABA. Alternatively, the positive correlation between ASL and GABA may be driven by CBF. Thus, the inverse correlation between BOLD and baseline GABA is likely due to a positive correlation between cerebral metabolic rate of oxygen consumption reactivity and GABA. Here, we systematically investigate these hypotheses by sequentially measuring AAT, CBF, and GABA in the occipital lobe of an age-restrictive cohort of healthy adult male volunteers. We also administered a standard neurocognitive task adapted from an established paradigm, 39 as cognitive impairment may confound the relationship between GABA and CBF.
MATERIALS AND METHODS
Patient Anonymity, Informed Consent, and Ethics
Anonymity of all volunteers was assured by removing the volunteers' names from data and figures. This study includes experiments on human subjects; procedures were followed in accordance with the ethical standards of the Vanderbilt University Institutional Review Board (IRB Study #101567), the Vanderbilt University Human Research Protection Program, as well as with the Helsinki Declaration of 1975 (and as revised in 1983). Informed, written consent was obtained from all volunteers and all components of this study were in compliance with the Health Insurance Portability and Accountability Act. No animal studies were conducted as part of this work.
Volunteer Demographics
Healthy volunteers (
Neurocognitive Task
The purpose of this component of the study was simply to ensure that all participants exhibited unimpaired cognitive capacity for their age and demographic. Participants completed a paradigm modeled after Dickerson
Imaging and Spectroscopy Protocol
Each volunteer underwent a 3.0T MRI (Philips Medical Systems, Best, The Netherlands) using an 8-channel SENSE coil for reception and quadrature body coil for radiofrequency transmission. The MEGA-PRESS MRS acquisition was performed in the same scan session, except with a birdcage coil for both radiofrequency transmission and reception to increase B1 performance. The MRI protocol consisted of (1) a 3D structural T1-weighted MPRAGE (repetition time/echo time=5.4/2.5 ms; turbo gradient echo factor=160; spatial resolution=1 × 1 × 1 mm3), and (2) a gradient-echo single-shot echo planar imaging (factor=35) multiinversion time (TI) pseudo-continuous arterial spin labeling (pCASL) sequence. The pCASL sequence (repetition time/echo time=4,350/18 ms; spatial resolution=3.43 × 3.43 × 6 mm3; slices=15) utilized a Hanning pulse train for blood water labeling with pulse duration=0.5 ms and total labeling duration=1,200 ms; postlabeling delay values=200, 500, 800, 1,100, 1,400, 1,700, 1,900, and 2,300 ms (12 averages per TI), dual adiabatic background suppression for concomitant suppression of gray-matter and white-matter static tissue signal. The pCASL labeling duration was reduced slightly (duration=1,200 ms) relative to more conventional labeling duration of ∼1,500 to 2,000 ms41, 42 to sensitize the approach to AAT, and the TI range was sampled with a temporal resolution of 300 ms over the approximate range of expected AATs. Finally, an equilibrium magnetization image (M0) was acquired with identical spatial resolution and readout as the pCASL scan, but with repetition time=20 seconds and all labeling and background suppression pulses removed.
For MRS, the 3-p.p.m. GABA peak ([GABA+], which reflects GABA+macromolecules) was measured using a previously published J-difference MEGA-PRESS approach43, 44 with repetition time/echo time=2,000/73 ms, 320 spectra with 2,048 samples each. The editing pulses were toggled between 1.91 and 7.4 p.p.m. on alternate scans and the editing pulses had a bandwidth of 64 Hz. Care was taken to place the spectroscopy voxel (voxel dimensions: 25 × 30 × 22 mm) similarly in the occipital cortex between all volunteers and with minimal partial voluming between tissue and sagittal sinus. To provide an additional level of control regarding voxel placement, gray- and white-matter voxel fractions were recorded and included in analysis, as outlined below. Optimization was performed before the spectroscopy acquisition to determine the pulse angle for improved water suppression. Four Regional Saturation Technique slabs using hyperbolic secant pulses targeted to the lipid resonance were placed orthogonally in the anterior–posterior and inferior–superior directions around the voxel to suppress the signal from lipids and to eliminate signal contamination from the scalp lipids. The REgional Saturation Technique slabs had a width of 30 mm, which was chosen to improve the profile relative to much larger REgional Saturation Technique slabs (50 to 100 mm) that are frequently used in other applications. While this approach proved effective to minimize conspicuous lipid contamination, residual lipids upfield of 1.9 p.p.m. were further mitigated by restricting the postprocessing quantification window to 1.9 to 4.0 p.p.m. The orientation of the spectroscopy relative to the pCASL and T1-weighted scans were recorded in all volunteers for subsequent coregistration and tissue volume segmentation.
Analysis: Neurocognitive Assessment
For the face/name task, recognition accuracy scores for each subject were calculated using
Analysis: Preprocessing
First, gray matter, white matter, and cerebrospinal fluid were segmented from the
Second, pCASL data were pair-wise subtracted and a single-compartment kinetic model was applied to simultaneously quantify CBF and AAT: 46
where
for tissue water
Third, for MRS analysis, the edited and unedited spectra were first corrected for phase and frequency variations between the 320 acquisitions using Creatine (Cr) as a reference peak. Since the editing pulse is asymmetrically disposed about the edited resonance, a differential phase results from this procedure. This effect was mitigated by applying a small differential zero-order phase correction to the respectively aligned, odd and even spectra. The corrected mean difference spectrum, obtained then by subtraction of the averaged unedited spectra from the averaged edited spectra, was processed using LCModel 50 to obtain a final estimate of the [GABA+]/[NAA–NAAG] ratio. To enforce spectral quality control, a Cramer-Rao Lower Bound (CRLB) threshold for [GABA+]/[NAA–NAAG] was utilized. Cramer-Rao Lower Bounds compare the known theoretical basis spectra and noise to predict an upper limit on the precision of the output concentrations obtained from LCModel. Larger percentages reflect poorer precision; in this study, CRLBs were restricted to 15% to ensure that data of poor quality were excluded from analysis. Finally, many studies separately normalize [GABA+] to [Cr]. Therefore, we additionally investigated the relationship between [GABA+]/[NAA–NAAG] and [GABA+]/[Cr]. For this purpose, the ratio of [NAA–NAAG]/[Cr] was independently estimated from only the even spectra. Multiplication of [GABA+]/[NAA–NAAG] by [NAA–NAAG]/[Cr] provided the value for [GABA+]/[Cr] ratio.
Data Analysis and Statistical Considerations
The primary objective is to assess the correlation between the MRI measurements (i.e., baseline CBF and AAT) and the MRS measurement (i.e., [GABA+]/[NAA–NAAG]). To achieve this, the GABA voxel was superimposed onto the quantified CBF and AAT (Δt) maps and CBF and AAT were recorded only for common cortical regions that overlapped between the CBF, AAT and MRS regions. A measure of error was calculated as the standard deviation (STD) of the CBF and AAT over the common region-of-interest or as the value of the CRLB of the [GABA+] measurement in the J-edited spectra. This analysis pipeline ensured that the same regions were being compared for the MRS and MRI comparisons. Next, normal probability plots and Pearson's
Additional secondary analyses were performed as well. Importantly, gray-matter fraction within the voxel may vary between volunteers, and therefore we additionally performed multiple regression analysis whereby the gray-matter voxel fraction, calculated from the
Finally, it is important to note that the AAT may contribute to final interpretation in two ways. First, AAT may correlate with [GABA+]/[NAA–NAAG], which was tested here using the multi-TI pCASL approach and sequential measurement of [GABA+]/[NAA–NAAG]. Alternatively, there may be no correlation between these two parameters, however, CBF quantified from single-TI pCASL data may show greater variability when uncorrected for AAT (e.g., a constant AAT assumption simply adds more uncertainty to the CBF measurement). Therefore, we repeated the above analyses when only the postlabeling delay=1,700 ms pCASL data were used for CBF quantification and the AAT was fixed at 0.8 second (calculated mean AAT over visual cortex).
RESULTS
For the neurocognitive task, all subjects performed within a normal range on this task with an average
Figure 1 shows the subject-averaged (A) pCASL difference images for different postlabeling delay times, whereby the changes in contrast between these postlabeling delay times are due to inflow of labeled blood water into the imaging slice. By applying the general kinetic model (equation (1)), it is possible to calculate (B) AAT and (C) CBF. All AAT and CBF calculations were performed in the native space of the pCASL images.
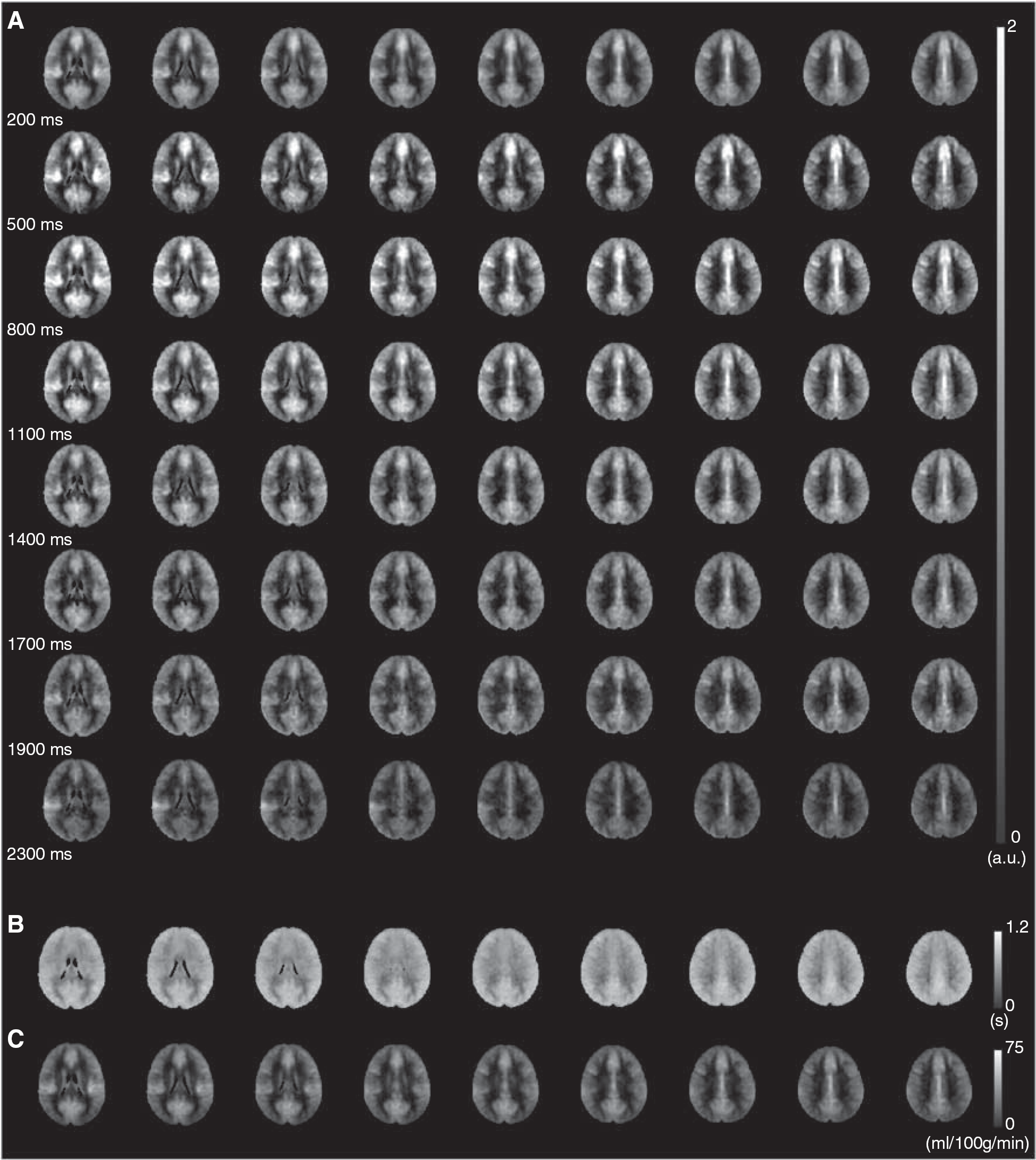
Quantification of cerebral blood flow (CBF) and arterial arrival time (AAT) is achieved from acquisition of pseudo-continuous arterial spin labeling (pCASL) data with differing postlabeling delays. (
Figure 2 shows a typical location for the MRS voxel. This region was used for analysis of both [GABA+]/[NAA–NAAG] (from the MRS data) and CBF (from the pCASL MRI data). The fractional gray matter and white matter within the voxel are shown as well (B–D). Gray-matter fraction within the voxel was found to be 36.3±2.0%. Therefore, the analysis region contains substantial contributions from white matter, and, to a lesser extent, cerebrospinal fluid. This gray-matter fraction is smaller than is typically reported in fMRI voxels (70% to 90%) due to the much larger size of the MRS voxel relative to typical fMRI voxel dimensions (e.g., 3 × 3 × 3 mm). However, the range of variability in gray-matter concentration was small (±2.0%), suggesting that bias from partial volume effects was similar in all volunteers.
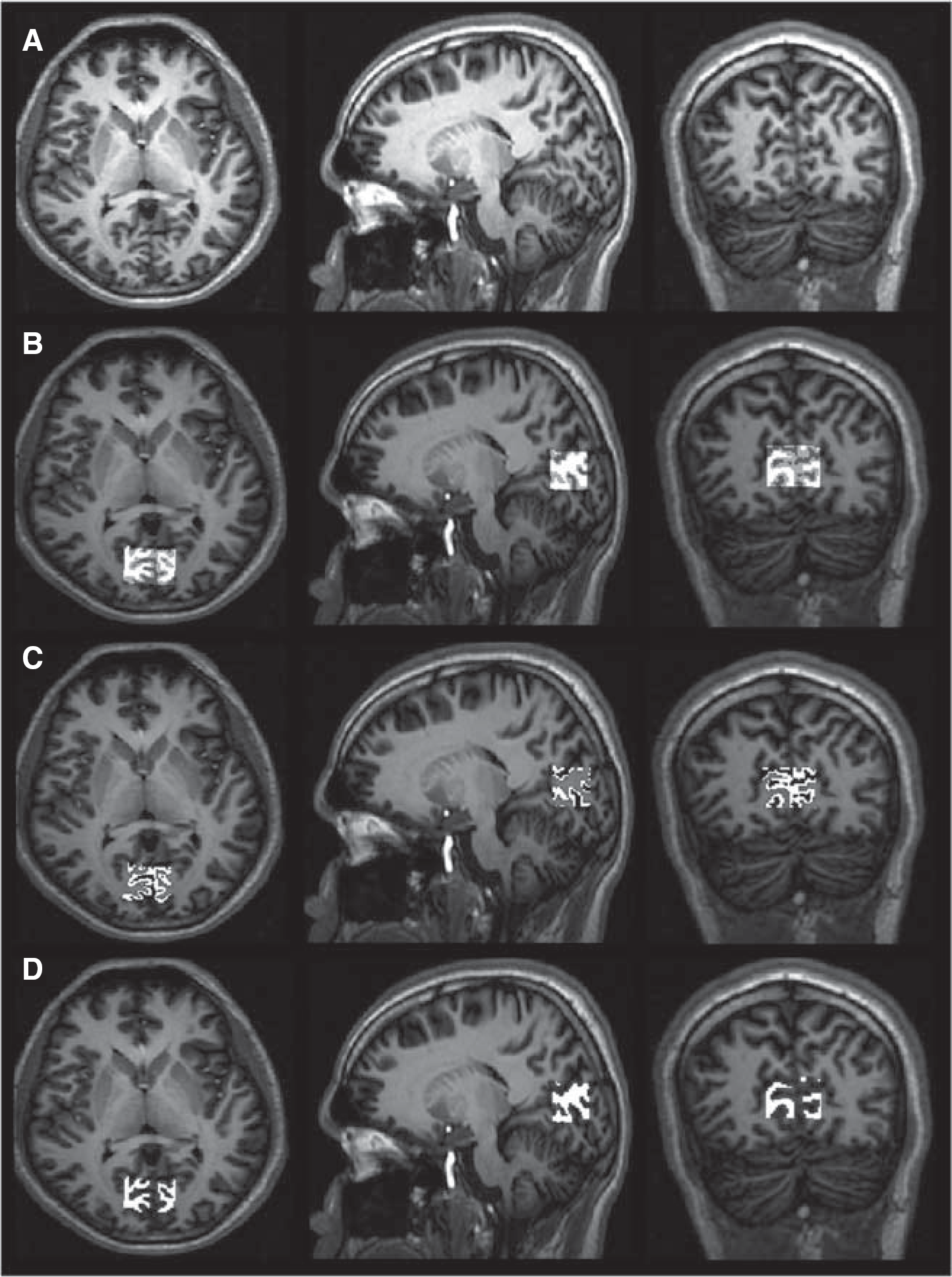
Region-of-interest procedure. The location of the
Figure 3 shows representative J-edited spectra from two subjects (A), along with the LCModel fit of these two spectra (B). In both cases, the GABA doublet at ∼3.0 p.p.m. is clearly visible; all volunteers were required to have CRLBs15% and therefore the quality of spectra from other subjects is similar to the quality of these presented cases. It is clear that Subject 7 shows a reduced GABA peak relative to Subject 8, and this volunteer was found also to have higher occipital CBF (C).
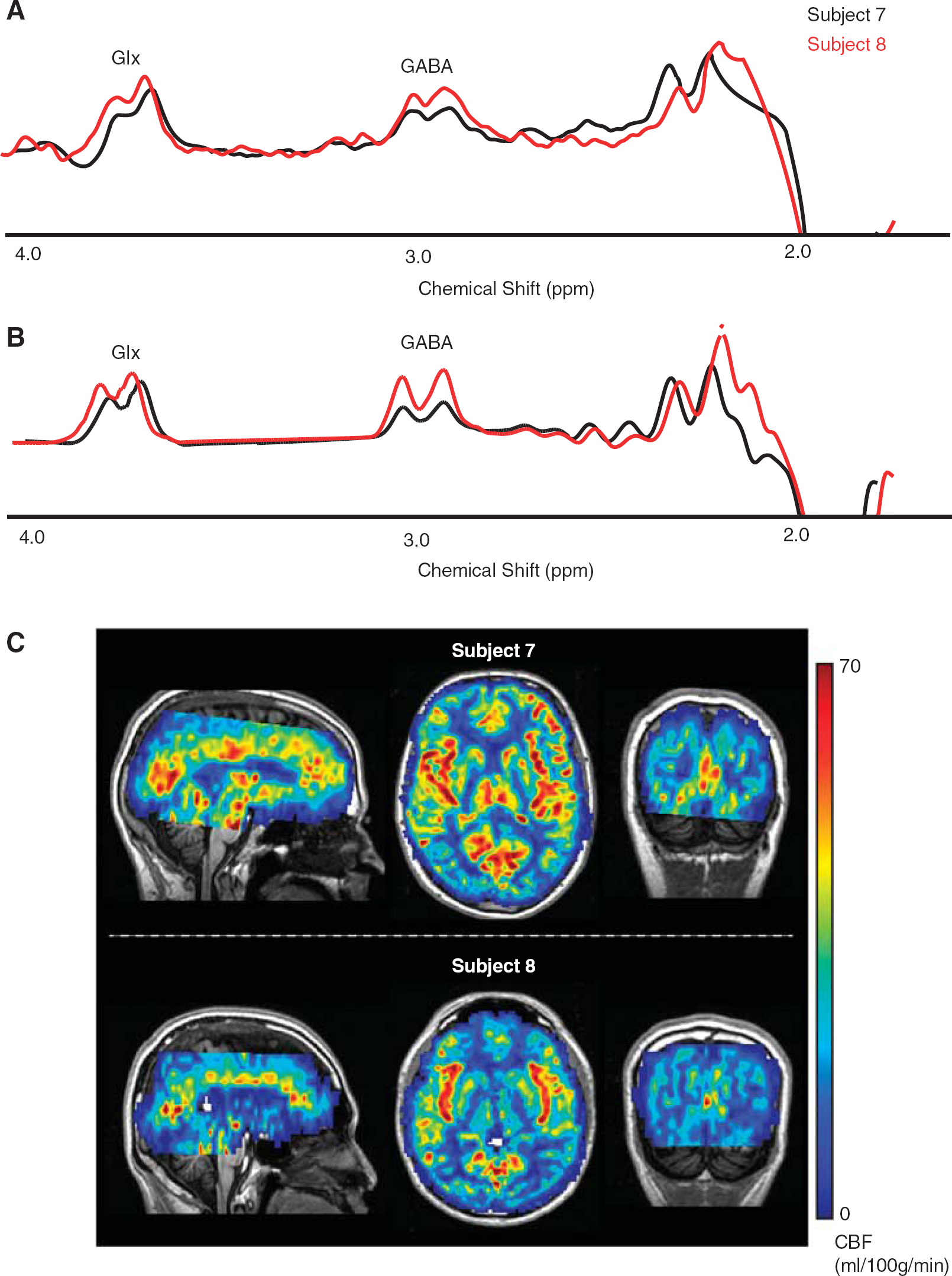
Figure 4 shows scatter plots from all subjects and data for (A) CBF (multi-TI) versus [GABA+]/[NAA–NAAG] (
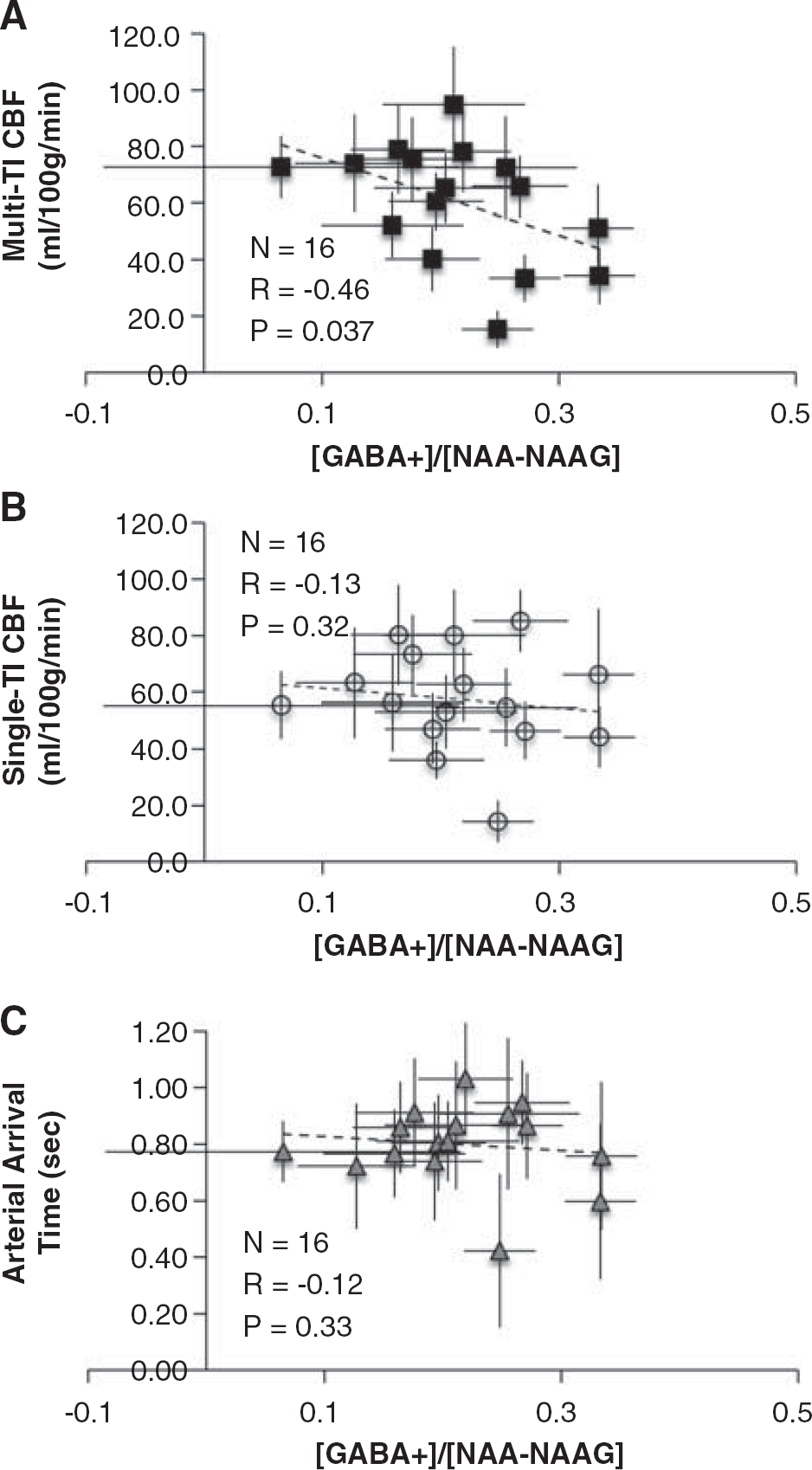
Scatter plots describing the relationships between [GABA+]/[NAA–NAAG] and the hemodynamic parameters. (
Figure 5 shows the results of the multiple regression analyses whereby multi-TI CBF (dependent variable) is investigated relative to [GABA+]/[NAA–NAAG], gray-matter voxel fraction, and AAT for the multi-TI fitting procedure. The adjusted
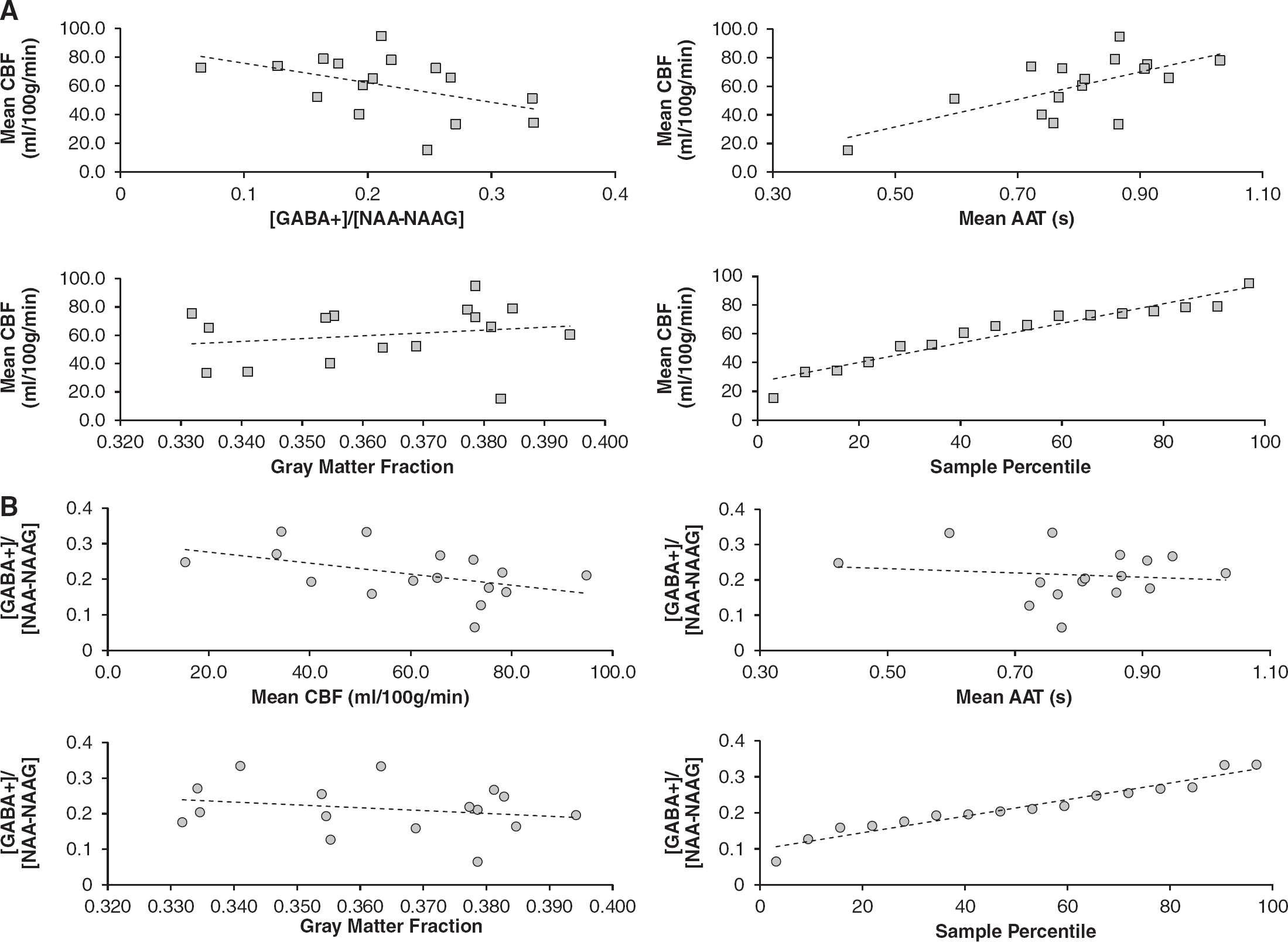
Multiple regression performed on the multiinversion time (multi-TI) imaging and spectroscopy data. Data points are shown along with the multiple-regression predicted-model fit (dashed line). (
Finally, we performed multiple regression analyses between [GABA+]/[NAA–NAAG] and CBF and gray-matter fraction, separately for the CBF calculated from the multi-TI versus single-TI fitting procedure. Previous correlative studies between GABA and CBF have only utilized single-TI procedures, which may be suboptimal when transit time varies between volunteers. In this regression, AAT was not included as a regressor as this metric is only available in the multi-TI fitting procedure (see Figure 5). Here as well, the multi-TI CBF described the [GABA+]/[NAA–NAAG] best (
In this study we chose to normalize [GABA+] by [NAA–NAAG], however a popular choice is also [Cr]. Therefore, we also evaluated whether [GABA+]/[NAA–NAAG] values were similar to [GABA+]/[Cr] values. It was found that [GABA+]/[NAA–NAAG] and [GABA+]/[Cr] correlated extremely tightly (
Table 1 provides CBF, AAT, and gray-matter concentration for all volunteers.
Measurements in all volunteers
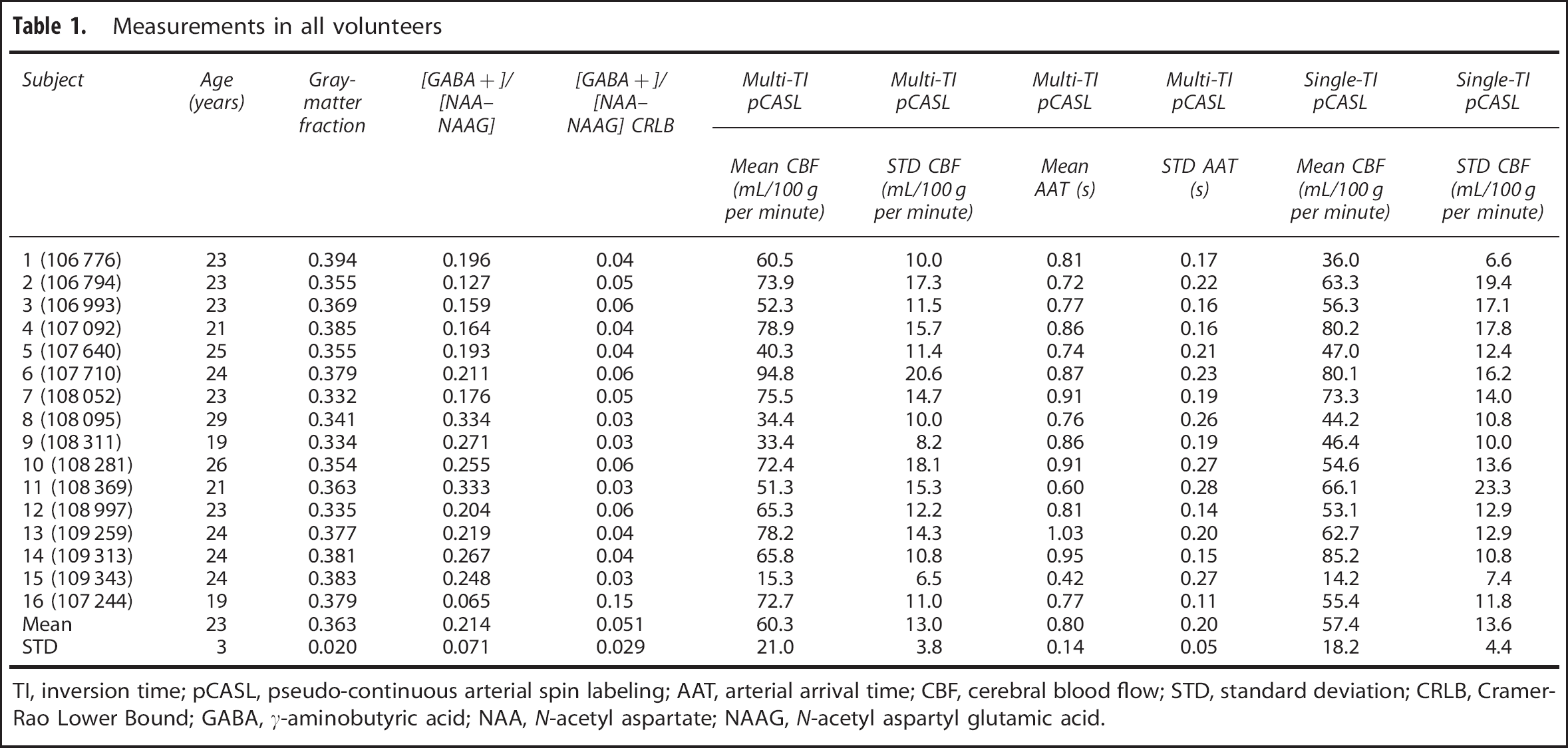
TI, inversion time; pCASL, pseudo-continuous arterial spin labeling; AAT, arterial arrival time; CBF, cerebral blood flow; STD, standard deviation; CRLB, Cramer-Rao Lower Bound; GABA,
DISCUSSION
The primary finding of this work is that measurements of occipital GABA ([GABA+]/[NAA–NAAG]) using J-edited MEGA-PRESS spectroscopy inversely correlate with CBF in the same region. This finding provides some physiologic basis for the recently reported inverse relationships between evoked BOLD responses and GABA. A secondary finding, which is important for explaining the prior inconsistencies between ASL-measured CBF and GABA, is that while blood arrival times do not appear to correlate strongly with [GABA+]/[NAA–NAAG], failure to account for arrival time in measurements of CBF can reduce or even eliminate the detectability of relationships between CBF and GABA.
It is important to consider these findings in the context of the AAT measurement. Recent work has performed multiecho ASL measurements and quantified T2, which is dependent on vascular compartment, to show that following spin labeling in feeding cervical vessels, blood water leaves the vasculature and enters tissue in less than 2 seconds. 51 Importantly, in ASL measurements where only a single postlabeling delay time is sampled (which is common given time constraints in many applications), it is not possible to measure the time that the blood water bolus reaches the imaging slice and rather a common value for this parameter is assumed. This metric has been used as a marker in steno-occlusive cerebrovascular disease 52 and has been implicated in memory disorders, 41 where blood velocity is reduced secondary to vascular stenosis, or alternatively, arrival time is increased owing to increased collateralization. Here, we observe that by not accounting for intersubject AAT variability, the relationship between ASL-measured CBF and [GABA+]/[NAA–NAAG] is considerably weakened. It is also important to note that the AAT measure is similar, yet fundamentally different from more common measures of blood arrival time using dynamic susceptibility contrast MRI with exogenous paramagnetic contrast agents. Here, either time-to-peak (time for the tracer to maximally attenuate signal in a voxel) or mean transit time (ratio of CBV/CBF or approximate time for tracer to traverse the capillary bed) is reported. Recent reports53, 54 have reviewed quantitative comparisons of ASL and dynamic susceptibility contrast in the context of different patient and healthy populations.
It was unexpected to find that the [GABA+]/[NAA–NAAG] measurement correlated only very weakly with the amount of gray matter in the voxel, as it is well known that GABA levels are higher in cortex than in white matter. A similar finding has been reported previously by other groups as well. 36 The likely reason for this pertains to the small range of variability in gray-matter fractions across volunteers, which simply does not supply sufficient range to observe this finding. It is likely that studies with larger voxel volumes, placed less consistently, may reveal this trend.
Regarding the phase correction procedure implemented in processing the MEGA-PRESS spectra, a recently published study has confirmed that alignment reduces variance and demonstrates that the quantification procedure should include additional corrections for optimal accuracy. 57 In this work, a differential phase between odd and even spectra is used to approximate the differential phase; however, an optimal method would be to use prior knowledge for the correction. Optimization of such correction procedures in future work is warranted.
These findings should also be considered in light of their potential biologic and clinical relevance to studies of behavioral control disorders, as well as poststroke plasticity mechanisms. First, it is well known that excitation-inhibition imbalance is implicated in a range of neuropsychological disorders. In practice, measuring CBF, with magnetic resonance, single photon emission computed tomography, or positron emission tomography, is considerably simpler than measuring GABA, and therefore it would be convenient if CBF represented a surrogate marker of GABA. While our results lend support for an inverse relationship between GABA and CBF, even in our strict volunteer cohort, this trend was not of sufficient statistical strength (
Animal studies have suggested direct roles of neurotransmitters after postischemic injury, and the relevance of poststroke GABA levels in humans has been suggested using pharmacological manipulation of GABA.
58
After stroke, there is a decrease in inhibitory activity, and some data have shown that in well-recovered patients increasing inhibition through GABAA agonists causes prior stroke symptoms reemerge.
58
Reduced GABA in M1 after ischemic stroke, and a correlation between GABA reduction and functional recovery has recently been reported.
59
Therefore,
In terms of functional imaging, it is important to note that BOLD signals are only indirect markers of neuronal activity. Recent correlative findings among neurotransmission, behavior, and BOLD responses are being presented more frequently, yet few efforts have been made to our knowledge to describe these relationships as more than qualitative correlations. Therefore, as more information regarding correlative relationships between
In conclusion, we measured basal GABA and CBF
Footnotes
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
The authors are grateful to David Pennell, Leslie McIntosh, Kristen George-Durrett, Donna Butler, Kevin Wilson, and Chuck Nockowski for experimental support.