
Abstract
The serotonergic precursor tryptophan and the dopaminergic precursor tyrosine have been shown to be important modulators of mood, behaviour and cognition. Specifically, research on the function of tryptophan has characterised this molecule as particularly relevant in the context of pathological disorders such as depression. Moreover, a large body of evidence has now been accumulated to suggest that tryptophan may also be involved in executive function and reward processing. Despite some clear differentiation with tryptophan, the data reviewed in this paper illustrates that tyrosine shares similar functions with tryptophan in the regulation of executive function and reward, and that these processes in turn, rather than acting in isolation, causally influence each other.
Keywords
Introduction
Serotonin (5-HT) has long been established as an important modulator of mood, particularly with respect to the pathophysiology of depression. A substantial amount of this evidence has come from studies employing the acute tryptophan depletion (ATD) procedure, where 5HT levels are lowered by reducing the availability of its precursor, tryptophan. 1 Despite this well-established role, 5HT has also implicated in the regulation of executive functions, specifically response inhibition and its relation to impulse control2-8 (with the dopaminergic system [DA] also having an important involvement). 9 Additionally, over the last decade several investigations in both animals and humans have implicated 5-HT in reward processing and goal-directed behaviour,10-16 domains normally attributed to be under the control of the DA system. 17
The purpose of this review and its related novelty is therefore to bring together research findings in which tryptophan and the dopaminergic precursor tyrosine were manipulated in the context of executive function and reward processing in humans. By comparing and contrasting tryptophan and tyrosine studies, we will be able to isolate the influences that presynaptic manipulations of serotonergic and dopaminergic activity have on these processes.
The evidence reviewed for both tryptophan and tyrosine will come from the following three sources: (1) ATD and acute phenylalanine and tyrosine depletion (APTD) studies (2) tryptophan and tyrosine loading and (3) polymorphisms in genes encoding tryptophan hydroxylase (TPH1 and TPH2). To the best of the author’s knowledge, there have been no investigations on the association between genetic polymorphisms in tyrosine hydroxylase (TH) and executive function/reward processing (of note, TH polymorphisms have been linked to bipolar disorders, 18 personality traits, 19 schizophrenia, 20 alcohol dependence 21 and suicide attempts) 22 and therefore TH studies will not be reviewed.
This review consists of the following four parts: (a) an evaluation of the literature on the serotonergic and dopaminergic systems (broadly) and their modulation of mood (depression in particular), executive function and reward processing; (b) an overview of the mechanisms of the ATD and APTD procedures (as most of the reviewed evidence relies on the ATD/APTD literature); (c) a summary of the findings which relate to the role of tryptophan and tyrosine in executive function; (d) and reward processing.
PubMed (https://pubmed.ncbi.nlm.nih.gov) was used to search for all articles. The publication selection criteria with respect to (c) and (d), consisted of the following: (1) the articles had to be original, peer-reviewed manuscripts, published in English from 1990 to 2020; (2) the studies must have been conducted in healthy volunteers but not in healthy and older individuals; (3) must have used cognitive tasks that measure executive function and reinforcement; (4) the articles were retrieved using the following search terms: ‘tryptophan’ ‘tryptophan depletion’, ‘tyrosine’, ‘tyrosine depletion’, ‘executive’, ‘reinforcement’, ‘reward’, ‘learning’, ‘TPH1’, ‘TPH2’. In order to minimize the risk of missing potentially relevant articles, the reference list of every retrieved paper in which the above search terms were used was extensively examined. Studies that failed to meet the above criteria were excluded. A summary of all the reviewed studies for section (c) and (d) has been reported in Tables 1–4.
Effects of tryptophan manipulations on executive function related tasks.
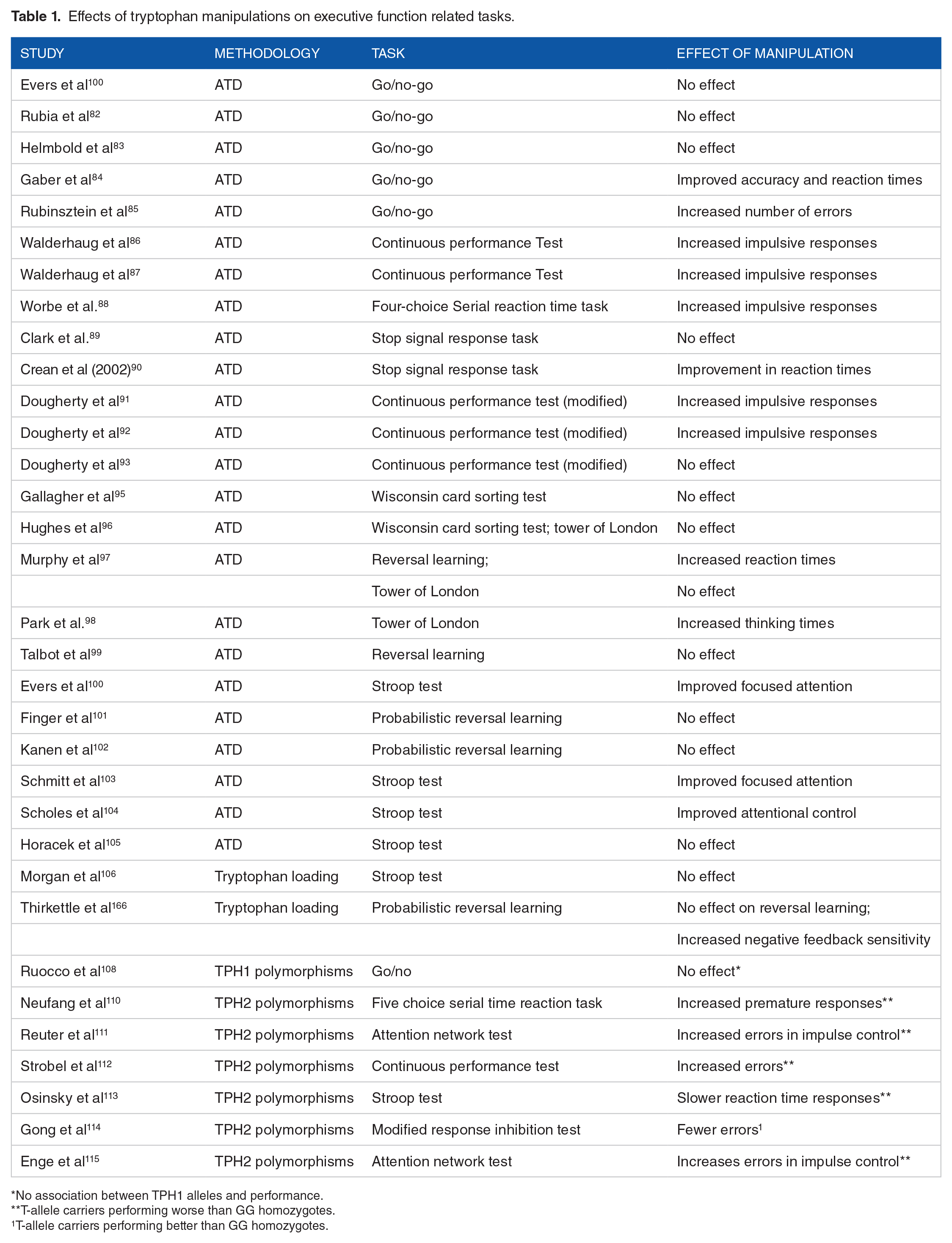
No association between TPH1 alleles and performance.
T-allele carriers performing worse than GG homozygotes.
T-allele carriers performing better than GG homozygotes.
Effects of tyrosine manipulations on executive function related tasks.
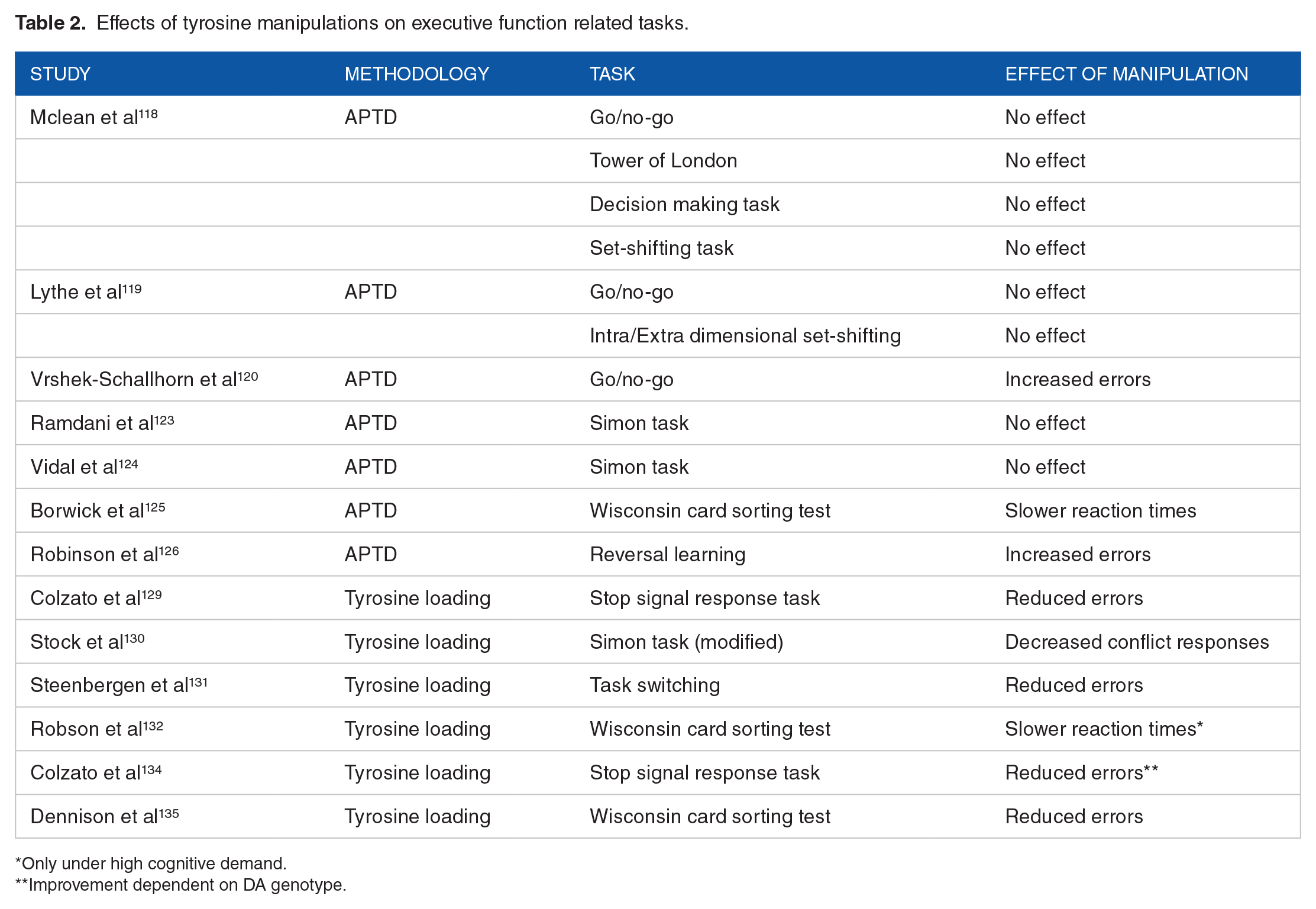
Only under high cognitive demand.
Improvement dependent on DA genotype.
Effects of tryptophan manipulations on reward processing related tasks.

Effects of tyrosine manipulations on reward processing related tasks.

Only in COMT val/val homozygous.
The serotonergic and dopaminergic neuronal circuitry and their modulation of depression, executive function and reward processing
Neuroanatomical characteristics of the serotonergic and dopaminergic systems
5-HT is primarily produced in the cell bodies of neurons located in the dorsal raphe nucleus (DRN) and medial raphe nucleus (MRN). 23 Whilst approximately 50% of DRN cells express 5-HT, this number drops to 5% for MRN neurons. 24 Both medial and dorsal parts of the raphe nucleus display extensive and widespread connections with sub-cortical and cortical regions of the brain. In particular, the DRN/MRN send projections to the amygdala, hypothalamus, thalamus, superior colliculus, medial prefrontal cortex (mPFC), sensorimotor cortex, caudate putamen and septum. 25 5-HT neurons often co-express other neurotransmitters, most notably GABA, glutamate and nitric oxide. 26 DA neurons, on the other hand, whilst also mostly localized in the midbrain, can be found in the ventral tegmental area (VTA) and substantia pars compacta (SNc). These 2 regions give rise to the mesolimbic, mesocortical (originating from the VTA) and the nigrostriatal (from the SNc) pathways. More specifically, VTA DA neurons innervate the nucleus accumbens (NAc), amygdala, hippocampus and mPFC, whereas the SNc DA neurons have axonal projections with the caudate and putamen of the striatum. 23 DA neurons also co-release nitric oxide, glutamate and GABA.
Functional interactions between the serotonergic and dopaminergic system
5-HT neurons innervate DA neurons in both the VTA and SNc but also in DA terminal fields (ie, NAc, mPFC and amygdala). 23 In addition to their anatomical interactions, administration of 5-HT agonists/antagonists, serotonin reuptake inhibitors (SSRI) and 5-HT lesioning, has been shown to modulate DA neurons activity. The response pattern of DA neurons to 5-HT is very complex and not yet fully understood, but a brief summary of the available evidence is here reported.
Early investigations in which VTA/SNc DA neurons were recorded in anaesthetized rats showed that application of 5-HT resulted in a weak inhibition of VTA DA firing and a more robust inhibition of SNc DA cells.27,28 Electrical stimulation of DRN cells, on the other hand, has been reported to inhibit SNc DA cells with a low firing rate, but excite others,29,30 whilst in VTA DA cells, the inhibition was dependent on whether these neurons were projecting to the NAc, whereas other VTA DA neurons were excited. 31 Using optogenetic techniques, stimulation of 5-HT terminals in the VTA and SNc produced excitatory postsynaptic potentials that were larger in magnitude for SNc DA than VTA DA cells, and critically, this effect was dependent on the co-release of glutamate. 32 5-HT excitatory effects on DA neurons have also been shown to be indirectly modulated by GABA mechanisms. 33 Synaptic increase in 5-HT by SSRI administration into the VTA (but not in the SNc), has been shown to result in reduced firing rates of VTA DA cells.34,35 However, these results appear to be dependent upon the type of SSRI administered, the route of administration (eg, systemic vs intracerebral), dosage, and whether the SSRI was given acutely or chronically. The effects of 5-HT lesions on VTA/SNc DA cells have produced mixed results, with both no changes in the firing rates of VTA/SNc DA cells, excitations and inhibitions.36-38 In addition to the complexities cited above, there is an abundant literature on the effect that different 5-HT receptors have on DA release, which demonstrate that DA signalling is affected by 5-HT receptor subtype (eg, 5-HT1a, 5-HT1b, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6, 5-HT7), DA target (eg, VTA vs SNc) and their reciprocal afferent/efferent connections, reviewed extensively elsewhere 23 and outside the scope of this paper.
Regulation of mood, reward processing and executive function by the serotonergic and dopaminergic system
As reported in the previous section, administration of SSRI affects DA cells activity. Using a variety of animal models of depression, accumulating evidence points to a shared role of 5-HT and DA in the aetiology of this mood disorder. For example, in the chronic mild stress test which attempts to mimic the symptoms of anhedonia, administration of antidepressants that are selective for 5-HT2c receptors, ameliorated performance and resulted in increased DA release in the NAc.39,40 Critically, when D2/D3 receptors were blocked, the beneficial effects of 5-HT2c stimulation on anhedonia were nullified. 41 In studies of monoamine transporter knockouts for both dopamine (DAT) and serotonin (SERT), impairments have been reported in measures of behavioural despair (ie, tail suspension test and forced swimming test) and anhedonia (ie, sucrose preference test).42,43 Furthermore, the involvement of the mesolimbic DA pathway in regulating mood and depressive-like behaviour outside of interactions with 5-HT has been well documented in the context of the transcription factor CREB, the opioid receptor dynorphin, the protein BDNF, hypothalamic peptides and circadian genes, as extensively reviewed by Nestler and Carlezon. 44
Given that many depressive symptoms as defined by the DMS-5 could be described as impairments in reward processing (ie, a lack of motivation to obtain rewards; inability to experience pleasure; deficits in reinforcement learning), 45 it is perhaps not surprising that both 5-HT and DA would regulate its function. Deficits in the ability to predict and anticipate a reward (a core aspect of reinforcement learning), as seen in depressed patients, have been ascribed to dysfunction within the mesolimbic DA pathway.46-51 Similarly, SSRI intake in healthy volunteers has been shown to diminish effort cost (ie, it increased motivation) which in turn mediated the ability to obtain more rewards. 52 Importantly, this SSRI effect might have been driven by increasing DA activity in the ventral striatum, a brain region known to be involved in reward processing. 53
In addition to the functional connection between mood disorders and reward processing as modulated by the serotonergic and dopaminergic systems, several lines of evidence suggest that those suffering from depression display impairments in cognition and, in particular, executive function. A meta-analysis by McDermott and Ebmeier, 54 reported significant negative correlations between increased depressive symptom severity and decreased executive function performance, in domains that included cognitive flexibility, planning, set-shifting and working memory.
In healthy populations and based on animal studies, the serotonergic system has been implicated in the regulation of response inhibition and reversal learning (mostly through the orbitofrontal cortex), whilst the dopaminergic system in measures of cognitive flexibility which include set-shifting (via the medial prefrontal cortex). 3
The acute tryptophan depletion procedure (cellular mechanisms)
One of the first attempts to lower 5-HT in humans was performed using an inhibitor of TH (PCPA). The authors found that administration of PCPA induced relapse in depression in patients who had reported improvements in depression severity following antidepressant treatment (by a MAO inhibitor and a tricyclic compound), 55 suggesting that PCPA may have counteracted the increasing 5-HT function of the antidepressants. 56 At around the same time, a version of the current ATD procedure (in humans) was initially tested in rats. In this investigation, the animals consumed a diet that was deficient in tryptophan. This manipulation resulted in lower plasma and brain tryptophan as well as serotonin and 5-hydroxyindoleacetic acid (found in the cerebrospinal fluid [CSF] and a marker of brain serotonin synthesis). 57 Further investigations revealed that tryptophan depletion was dependent on protein synthesis, since administration of a protein synthesis inhibitor could block the fall in tryptophan depletion levels by the ATD procedure.58,59 In humans, the first demonstration that tryptophan levels could be lowered was shown by the Concu group, 60 who gave participants a tryptophan free mixture which resulted in reduced serum tryptophan and increased anxiety compared to a group who consumed a tryptophan balanced mixture. Several studies following the original Concu investigation have confirmed that the ATD procedure lowers serotonin synthesis in the brain, 61 and additionally, the rate of 5-hydroxyindoleacetic acid in the CSF.62-66
What remains to be established, at the very least in humans, is whether the ATD procedure results in reductions in 5-HT release (in rats using microdialysis these reductions were dependent upon the coadministration of 5-HT reuptake inhibitors). 67 It is not clear, for example, whether decreases in 5-HT synthesis would correspond to linear decreases in 5-HT firing and/or release. One factor that is unrelated to 5-HT synthesis in the regulation of 5-HT firing is behavioural arousal and/or motor activity. 68 It is also likely that the degree of 5-HT release/firing altered by the ATD procedure would vary by brain location.56,61 Unrelated to 5-HT release, the ATD procedure may result in effects that are not directly related to 5-HT, including changes in BDNF, 69 amino acid balance, 70 melatonin, 71 and nitric oxide synthase.72,73
The acute phenylalanine and tyrosine depletion procedure (cellular mechanisms)
It was again the Biggio group in 1976 who were the first to attempt to lower DA in animals in what we now describe as the APTD procedure. Using an amino acid mixture lacking tyrosine and phenylalanine, they demonstrated in rats that tyrosine in serum and in the brain (basal ganglia specifically) could be lowered by 73% 2 hours post ingestion. 74 The exclusion of phenylalanine is required as this amino acid can be converted to tyrosine or tyrosine hydroxylase. 75 M and J Fernstrom replicated Biggio’s findings two decades later. 76 The authors demonstrated that serum tyrosine and hypothalamic tyrosine levels could decrease significantly for 60 to 180 minutes due to the APTD. A year later, it was Sheehan and colleagues to confirm in humans that tyrosine plasma levels could be lowered by up to 50% over a 5-hour period by the APTD. 77 As for the ATD procedure, there is lack of evidence in humans that the APTD can result in reductions in DA release. Using microdialysis in rats, however, McTavish and colleagues showed that the APTD decreased extracellular DA release in the striatum when amphetamine was co-administered. 78 Interestingly, DA release did not change under basal conditions (ie, when comparing APTD to control), in a manner that resembles the 5-HT release by the 5-HT reuptake inhibitors, 67 suggesting that there are specific concentration thresholds of DA and 5-HT that are necessary in order to see declines in extracellular release of these neurotransmitters under APTD and ATD. The McTavish study was also particularly important as it revealed that noradrenergic release was not affected by the APTD-amphetamine combination, indicating that the APTD preferentially affects DA neurons despite tyrosine and phenylalanine acting as molecular precursors to DA, noradrenaline and adrenaline.
The role of tryptophan in executive functions
ATD studies
The bulk of the evidence regarding the role of tryptophan in executive functions has been revealed by ATD studies. The three main cognitive domains that have been investigated are response inhibition, selective attention/cognitive interference and reversal learning. As one might expect, not all studies have administered the same cognitive task to assess a particular cognitive domain. In response inhibition studies, for example, the tests used include the Go/Nogo task, the Stop Signal task, the Continuous Performance Test, the 5-choice serial reaction time task, the Simon task, the loss avoidance in a decision-making task. Although one can assume a degree of similarity in the way all of these tasks are encoded by fronto-striatal neural circuitry, 79 it is plausible that there might be subtle differences in neural activation (not precisely captured by functional neuroimaging studies) as a result of these tasks which may account for diverging outcomes. Most of the studies reviewed here only reported behavioural effects, however there were a handful of examples in which both neuroimaging and behavioural data was gathered, which are going to help us define the neuronal networks involved.
In a study assessing response inhibition using the Go/Nogo task, the authors found that the ATD had no impact on brain activity in relation to response inhibition, however, the ATD did reduce dorsolateral prefrontal cortex activity during performance monitoring. 80 In the Go/Nogo task, response inhibition is measured as the percentage of correct responses during Go and Nogo trials. Performance monitoring, on the other hand, is assessed by measuring the reaction times on a correct response following an error, a well-known form of post-error slowing, 81 which can also be measured by administering a reversal learning task. In contrast to the above findings, Rubia and colleagues, also using the Go/Nogo task, did report a neural effect of the ATD during response inhibition with reduced right orbitoinferior (OFC) prefrontal and increased superior and medial temporal cortices activation. 82 Evers and colleagues speculated that the reasons for the contrasting findings may relate to task difficulty of the Go/Nogo task, performance feedback
provided during the task and gender differences between their study and that of Rubia et al. Behaviourally, both reports did not find an effect of the ATD on response inhibition and performance monitoring. This is collaborated by an additional report in which no behavioural effect was found, however, during the Nogo (punishment) trials healthy female volunteers had reduced medial OFC and anterior cingulate cortex (ACC) activation compared to controls. 83 In contrast to these findings, a body-weight adjusted ATD experiment found improvements in accuracy and reaction times in the same task by reductions in tryptophan levels, 84 and opposite findings have been reported in another. 85 Overall, it would seem that whilst several of these prefrontal regions process stimuli that relate to the Go/Nogo task, their activation is not causally related to performance. Moreover, the behavioural evidence is mixed with a combination of null findings, improvements and impairments following ATD administration.
Behaviourally, two studies (by the same research group) that adopted the Continuous Performance Test to measure response inhibition have both found that the ATD impaired performance86,87: a similar impairment was reported when the 5-choice serial reaction time task was adopted. 88 In contrast, two studies that assessed response inhibition using the Stop Signal task found either no effect of the ATD on performance, 89 or even an improvement on the stop reaction time by the ATD. 90 These data again indicate that different cognitive tasks measuring response inhibition (underpinned by similar but not identical neurobiological mechanisms) likely contribute to the relative heterogeneity of the findings.
Using a modified version of the Continuous performance task, Dougherty and colleagues, showed that those in the ATD conditions had significantly impaired performance compared to baseline (ie, prior to ATD) and to those in the tryptophan loading group 91 . This finding was also replicated in a follow up study both in reference to baseline and to placebo. 92 In a third study by the same group, the authors investigated the interaction between alcohol consumption and ATD and found that the ATD combined with alcohol did not increase/decrease response inhibition. 93
In a separate study measuring ‘Reflection Impulsivity’, that is, making decisions before having gathered enough information, the Trevor Robbins’ lab showed that the ATD did not affect reflection impulsivity, but rather it promoted avoidance of small, immediate (reward) losses, 94 and hence appeared to have a more specific role in aversive processing.
In a study assessing set-shifting (Wisconsin Card Sorting test), ATD administration had no impact on performance. 95 The same research group replicated these findings in a separate investigation, including no effect of the ATD on planning abilities (as measured by the Tower of London test).96,97 This is interesting given that set-shifting is a subset of cognitive flexibility and suggests a possible separation of tryptophan effects on different measures of cognitive flexibility. Nevertheless, a separate research group demonstrated that the ATD can negatively impact planning abilities in the Tower of London test but only in those familiar with the task, suggesting a specific detrimental effect on performance retrieval rather than acquisition learning. 98
The ATD has also been reported to negatively impact reversal learning, 97 but a later study failed to replicate the reversal learning impairment. 99 The effect of tryptophan on probabilistic reversal learning (PRL), a form of reversal learning which can assess negative feedback sensitivity (a measure of switching behaviour during the task when misleading feedback has been provided) has also been tested in three additional studies. None of these studies reported a significant effect of ATD on PRL performance.100-102
Using the Stroop test, three investigations reported improvements in selective attention following ATD administration,100,103,104 with the ATD increasing the BOLD signal in the anterior cingulate cortex (ACC) and precuneus. A fourth study did not find a behavioural effect but did report increased activation in the ACC and PFC. 105
Taken together, the evidence reviewed suggests a potential role for tryptophan and the prefrontal cortex (PFC) (as a whole) in modulating some forms of response inhibition, which is in line with studies that have identified serotonin, more generally, and the prefrontal cortex in the regulation of response inhibition. Furthermore, tryptophan seems to modulate cognitive interference/selective attention but not reversal learning.
Tryptophan loading studies
There has been a real scarcity of tryptophan loading studies assessing executive function in healthy (several more studies have been published in clinical samples) participants. To the best of the author’s knowledge, this was a study by Morgan and colleagues who administered tryptophan and found decreased activations in prefrontal regions with no behavioural effect on a cognitively demanding Stroop task. 106 The second study was a recent investigation by our research group in which we reported an increase in negative feedback sensitivity but no change in reversal learning as a result of tryptophan loading (compared to a placebo group).
TPH1 and TPH2 studies
Studies for TPH1 and THP2 genes have been grouped together with the caveat that TPH1 and TPH2 are differentially expressed in the brain, 107 and therefore careful interpretation of the data needs to be applied.
In a study using the Go/Nogo task, the authors tested the hypothesis that single nucleotide polymorphisms in the TPH1 gene, would affect brain activation and performance on the task. The authors found no behavioural evidence for this but did show that those with the ‘risk’ allele for TPH1 had reduced medial prefrontal cortex activity during response inhibition. 108 Importantly, the ‘risk’ allele for TPH1 has been linked to lower serotonin turnover. 109 A different research group, instead, looked at the effect of single nucleotide polymorphisms in the TPH2 gene on the 5-choice serial reaction time task and brain activity. Here, the authors found that the risk allele (T/T homozygous on the 703 polymorphism) resulted in a greater number of premature responses (a form of inhibitory control and/or ‘waiting impulsivity’) which was related to diminished ventromedial prefrontal cortex activity. 110 Deficits in response inhibition and impulse control have also been reported on the same T/T genotype of TPH2 by 3 separate research groups,111-113 despite a contradictory finding by a fourth in which however the T/T and G/T genotypes were grouped together and compared to the G/G. 114 Using EEG recordings, Enge et al were able to reveal some of the putative mechanisms by which response inhibition performance is superior in the G-allele versus T-allele homozygotes in that those with the G-allele displayed increased event-related gamma-band activity during the task. 115 Critically, gamma-band activity has been reported to modulate top-down attentional selection by amplifying relevant sensory inputs and inhibiting attention to irrelevant ones,116,117 hence facilitating response selection.
The role of tyrosine in executive functions
Tyrosine depletion studies
The first use of the APTD procedure to understand the impact of reductions in putative dopaminergic synthesis on executive function in healthy volunteer was a study conducted by McLean et al. Using a parallel group design (crossover repeated measures can be problematic with the APTD due to the unpalatability of the amino acid drinks and the potential for a high withdrawal rate between repeated sessions), the APTD did not affect measures of planning, decision making and set-shifting compared to the balanced (control) group. 118 These findings were largely replicated by Lythe and colleagues (this time using a crossover repeated measure design) as neither set-shifting nor response inhibition nor impulsivity were significantly affected by the APTD. 119 Response inhibition was negatively affected by the APTD as reported in a later study, 120 however, it is worth noting that the statistical analyses contained multiple comparisons (ie, a total of 14 measures for the response inhibition task were taken) which did not appear to control for false discovery rate such as by the application of the Benjamini-Hochberg adjustment, as recommended elsewhere. 121 Response inhibition as measured by the Simon task was unaffected by the APTD in two additional studies,122,123 nevertheless, changes in the N-40 component (an event related potential which has been proposed to represent response selection) 124 by the APTD were noted but these neurophysiological changes were unrelated to behavioural performance. In a recent study by our research group, we reported a negative effect of the APTD on task-switching using the Wisconsin Card Sorting Task, an effect which could be ‘rescued’ by anodal (ie, excitatory) transcranial direct current stimulation (tDCS) of the dorsolateral prefrontal cortex, 125 highlighting a potential neuronal network which may underpin APTD effects on task-switching. On the contrary, Robinson and colleagues demonstrated that the APTD had a beneficial effect on reversal learning in healthy female participants but not in males. 126 This improvement in females was driven by increased sensitivity in punishment processing following an error during reversal trials. These results parallel some of the findings reported in the ATD literature.
Tyrosine loading studies
In contrast to the APTD studies where the overall pattern of results would suggest that being tyrosine depleted does not affect executive function (but see), 124 tyrosine loading studies have been characterized by more consistent results. A plausible explanation for this difference may lie in the degree to which the APTD suppresses DA availability in contrast to the increases in DA availability by tyrosine loading. That is, both theoretical and experimental evidence support the view of an inverted-U shaped function between DA and cognitive function, whereby too low or too high levels of DA may lead to detrimental performance, with an optimal level occurring somewhere in the middle and providing the most (cognitive/behavioural) benefits. 127 Whilst APTD studies have used largely similar amino acids concentrations, the same cannot be said for tyrosine loading studies where dosages have varied from as little as 500 mg to 12 g. 128 Therefore, it is possible that the 50% reductions in plasma tyrosine levels by the APTD as reported by Sheehan and colleagues, 77 may not always be sufficient to create a dip in DA availability that is on the left-bottom side of the inverted-U curve. On the contrary, the combination of more varied dosages in tyrosine loading studies and the fact that tyrosine conversion to dopamine is limited by the competition from other endogenous amino acids and by the TH enzyme, means that even when higher dosages of tyrosine have been administered, these are unlikely to result in DA availability that is on the right-bottom side of the inverted-U curve. 128
Two studies by the Colzato group demonstrated an improvement in two separate measures of response inhibition by tyrosine supplementation when compared to a placebo group.129,130 Similar results were obtained when measuring task switching. 131 Interestingly, our research group found that when task switching was measured under cognitively demanding conditions (ie, after having completed a digit spat memory task), tyrosine was unable to enhance performance. 132 Previous studies have demonstrated that under stressful, demanding situations, catecholamine activity is increased, depleting neurotransmitter levels. 133 In line with the inverted-U shaped hypothesis of DA function and cognitive performance, we speculated that tyrosine supplementation in our study was insufficient to replenish catecholamine levels to improve task-switching above baseline. This view is reinforced by an additional study in which tyrosine supplementation was beneficial to response inhibition performance only in those who were T/T homozygotes in the DA D2 receptor gene and hence with lower baseline striatal DA levels, 134 compared to C/C homozygotes with higher striatal DA levels. Finally, our tDCS findings in relation to anodal stimulation of the dlPFC, the APTD and cognitive flexibility (task-switching), 124 were replicated in another study where tyrosine administration improved cognitive flexibility (over placebo), but this effect could be ‘nullified’ by cathodal (inhibitory) stimulation of the dlPFC. 135
The role of tryptophan in reward processing
ATD studies
As discussed in a previous section accumulating evidence suggests that deficits in the regulation of mood (eg, depression) may be related to impairments in executive function, and reward processing. The latter will be discussed in this section with reference to ATD studies.
In a series of studies by the Cools and Robbins group using both instrumental and Pavlovian tasks that allowed to separate reward processing from punishment, the authors reported that the ATD facilitated prediction for punishment but not for reward.136-140 Neuroanatomically, neural responses to negative outcomes (eg, losses) by the ATD have been reported in the dorsomedial prefrontal cortex and amygdala. 141 The findings by the Cools and Robbins group have been confirmed by a number of investigations, whereby the ATD worsened performance for goal-directed behaviour under reinforcement but improved it for goal-directed punishment (ie, the avoidance of loss),142,143 and by studies on Pavlovian-instrumental transfer (PIT) looking at appetitive and aversive outcomes.144,145 However, an investigation by Rogers and colleagues only reported an impairment for reward but no improvement for punishment prediction, 146 similar to reports in either probability discounting, 147 counterfactual learning, 148 and a risk-seeking task. 149 Additionally, in a decision making task, the ATD reduced vmPFC activity in relation to reward outcome but not to punishment avoidance. 150
In support of the idea that tryptophan is not only important for punishment prediction but also reward representation, a series of pivotal studies by the Doya group demonstrated that the ATD affects reward discounting. More specifically, Doya and colleagues found that tryptophan depletion led to a preference for small, immediate rewards over larger delayed rewards.151-153 Moreover, in a separate ATD/loading study, Doya and colleagues demonstrated that whilst low tryptophan levels (induced by the ATD) correlated with encoding of reward prediction errors at short timescales, higher levels of tryptophan (induced by tryptophan loading) were correlated with prediction errors at longer timescales. 153 These results complement the extensive literature on the role of DA in reward prediction error (ie, the difference between expected and obtained reward) and add the dimension of timing to the way in which rewards are encoded. Finally, Worbe and colleagues showed that the ATD produced a shift from goal-directed behaviour to habitual like responses, 154 a finding that resembles manipulations of the dopaminergic system in the striatum. 155
Tryptophan loading studies
As for the scarcity of studies reported using tryptophan loading in the context of executive function in healthy populations (see section 5b), a similar pattern emerged for reward processing. Again, to the best of the author’s knowledge, only two publications could be found both by the Doya group. In the first study, tryptophan loading was correlated with prediction errors at longer timescales. 153 In the second, tryptophan loading neither affected learning from delayed punishments or delayed rewards. 156
TPH1 and TPH2 studies
There were no studies that isolated the effects of either TPH1 or TPH2 polymorphisms in healthy volunteers and tested these in experimental cognitive tasks that measure reward processing. In two cases, a composite measure of the 5-HTTLPR (S allele( and TPH2-703 (G allele) genes was related to diminished altruistic punishment (ie, ‘impulsive response to perceived provocation resulting from uncooperative behaviour’), 157 whereas in a second study, the combination of polymorphisms from the candidate genes TPH2, HRTR2A and HRTR1A was related to differences in reward sensitivity. 158
The role of tyrosine in reward processing
Tyrosine depletion studies
In line with extensive evidence from animal studies of the role of DA in reward processing, several APTD investigations in healthy volunteers have replicated these findings. For example, in a study by Hebart and Glasher, 145 the APTD selectively impaired PIT for appetitive cues, consistent with the established role of dopamine in reward prediction. The APTD has also been reported to affect incentive salience and motivation for reinforcement which again mirrors the animal literature. Leyton and colleagues found that the APTD reduced the ability to respond to stimuli that predicted reward, and that co-administration of L-Dopa (a DA agonist) could prevent this impairment. 159 Moreover, Bjork et al using fMRI were able to pinpoint the neuronal signal that was responsible for reduced processing of appetitive cues. NAc activity was reduced following APTD administration when participants were viewing cues that anticipated high rewards. 160 A similar finding was reported by Nagano-Saito et al at a neuronal network, although the APTD did not affect behavioural performance. 161 These neuroimaging results were consistent in a third study by Frank and colleagues, 162 who furthermore tested whether body mass index (BMI) may mediate this response, given that reward sensitivity and DA alterations are associated with obesity. The authors did not find a mediation effect of BMI on reward processing, suggesting that other factors in obesity may be at play in determining reward-related responses. Kelm and Bottinger, on the other hand, reported a genetic × APTD interaction in reward processing. More specifically, the authors found 163 that those homozygous for the val/val allele of the COMT genotype (encoding an enzyme that degrades extracellular DA particularly in the prefrontal cortex), and that were administered the APTD displayed increased impulsive choice for immediate rewards. Val/val individuals are known to have the lowest level of DA activity 164 (compared to val/met and met/met), and these findings therefore demonstrate that the combination of reduced DA synthesis by the APTD coupled with lower baseline levels of DA activity in the val/val COMT genotype negatively impacted performance as previously suggested by the theory of an inverted-U shaped function with respect to DA and cognition.
Finally, de Wit and colleagues replicated the findings of Worbe et al for the ATD, in that the APTD also produced a similar shift from goal-directed to habitual like responses in a reinforcement task. 165
Tyrosine loading studies
No studies assessing the impact of tyrosine loading on reward processing tasks in healthy volunteers could be found.
Conclusion and future research
Accumulating evidence over the past three decades has identified the serotonergic precursor tryptophan and the dopaminergic precursor tyrosine as involved in the regulation of executive function and reward processing.
The key take home point of this review is that the functional interactions that occur at the cellular and neuronal network level between the 5-HT and DA system contribute to a shared role of tryptophan and tyrosine in various subcomponents of both executive and reward processing, despite some clear differentiation. For both tryptophan and tyrosine related studies, there remains a substantial research gap which would need to address how the ATD/tryptophan loading and the APTD/tyrosine loading procedures affect 5-HT and DA release in the human brain. Nevertheless, neuroimaging (mostly in the form of fMRI), genetic, and non-invasive brain stimulation investigations have started to pave the way for a more comprehensive understanding of the mechanisms by which tryptophan and tyrosine affect executive function and reward processing.
Footnotes
Declaration of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.