
Abstract
The acute tryptophan or tyrosine plus phenylalanine depletion and loading tests are powerful tools for studying the roles of serotonin, dopamine and noradrenaline in normal subjects and those with behavioural disorders. The current amino acid formulations for these tests, however, are associated with undesirable decreases in ratios of tryptophan or tyrosine plus phenylalanine to competing amino acids resulting in loss of specificity. This could confound biochemical and behavioural findings. Compositions of current formulations are reviewed, the biochemical principles underpinning the tests are revisited and examples of unintended changes in the above ratios and their impact on monoamine function and behaviour will be demonstrated from data in the literature. The presence of excessive amounts of the 3 branched-chain amino acids Leu, Ile and Val is responsible for these unintended decreases and the consequent loss of specificity. Strategies for enhancing the specificity of the different formulations are proposed.
Keywords
Introduction
Cerebral monoamines control many physiological functions, not only in the periphery, but also in the central nervous system. Many behavioural processes in the brain are associated with these monoamines, particularly 5-hydroxytryptamine (5-HT or serotonin) and the catecholamines dopamine (DA) and noradrenaline (NA). These processes include affect, aggression, anxiety, appetite, arousal, cognition, drive, emotions, impulsivity, mood, movement, reward and self-control. The acute tryptophan (Trp) depletion (ATD) and loading (ATL) tests 1 are powerful research and diagnostic tools for studying the role of serotonin in some of the above behaviours in healthy volunteers and in those with psychiatric and other behavioural disorders. Thus, hundreds of such studies have administered these tests to examine behaviour in healthy volunteers1–4 and many disease conditions, including aggressive behaviour, 5 alcoholism, 6 Alzheimer's disease, 7 anorexia nervosa, 8 anxiety disorders, 9 autistic disorder, 10 bipolar affective disorder, 11 bulimia nervosa, 12 depression, 13 insomnia, 14 irritable bowel syndrome, 15 obsessive-compulsive disorder, 16 panic disorder, 17 premenstrual syndrome, 18 schizophrenia 19 and seasonal affective disorders. 20
The acute tyrosine (Tyr) and phenylalanine (Phe) depletion (ATPD) test 21 is similarly used to assess the role of the catecholamines dopamine (DA) and noradrenaline (NA) in normal subjects and those with psychiatric and other behavioural disorders. Because of the preferential role of serotonin in many of the conditions described above and as the ATPD test 21 was developed 11 years after the ATD test, 1 fewer studies of the ATPD test have been performed, e.g. in healthy volunteers,22,23 cigarette smokers, 24 depression 25 and manic illness. 26 As far as we could ascertain, no attempts have been made to study the effects of acute Tyr and Phe loading (ATPL) by giving amino acid mixtures rich in Phe and Tyr. Previous studies on Tyr loading were performed mainly in animals, examined aspects such as acute stress, the acute respiratory syndrome, endurance, muscle strength and catecholamine excretion, and involved administration of a single dose of Tyr, usually alone, but sometimes in combination with another amino acid (see, e.g. ref 27 ).
More recently, some investigators28–31 have attempted to investigate behavioural measures under conditions leading to simultaneous depletion of serotonin, dopamine and noradrenaline by performing combined ATD and ATPD depletion tests by giving an amino acid mixture lacking in all three relevant amino acids, namely Trp, Phe and Tyr.
In this review, the nature and biochemical basis of the above tests will be discussed, drawing on important biochemical mechanisms derived from animal studies. The poor specificity in humans of the current amino acid test and control formulations will be demonstrated, its cause will be identified and strategies for enhancing it will be proposed. In seeking to illustrate the above aspects, this review will not be exhaustive and will be limited as far as possible to human studies, as species differences in amino acid metabolism render direct comparison difficult.
Nature of the tests and their current amino acid formulations
Both the ATD and ATPD tests involve administration to the subjects under study of a mixture of up to 15 amino acids (AA) (both essential and non-essential) which lack tryptophan (Trp) (for the ATD test) or lack phenylalanine (Phe) and tyrosine (Tyr) (for the ATPD test). Both Phe and Tyr, rather than Tyr alone, must be omitted from the formulation, because Phe can be converted to Tyr by Phe or Tyr hydroxylase.32,33 By the same token, when loading, as opposed to depletion, is required, the amino acid mixture will then contain an excess of Trp (for acute Trp loading or ATL) or an excess of Phe and Tyr (for acute Phe plus Tyr loading or ATPL) and moderate amounts of the corresponding amino acids (Phe and Tyr for the ATL and Trp for the ATPL tests). Additionally, in either the depletion or loading studies, a “balanced” amino acid mixture is administered as a control treatment. This control formulation will contain the same AA used in the depletion or loading tests, but with moderate amounts of Trp, Phe and Tyr.
Table 1 gives the amino acid (AA) composition of the ATD, ATL and control balanced formulation originally published by Young et al 1 in amounts per a 100 g dose. Because women weigh on average 16.7% less than men, the original 103 g AA mixture used by this Canadian research group was subsequently lowered to 85.5 g for studies in women. 2 Other investigators have introduced minor variations to the above original formulation, 1 mainly to increase the Trp content in the control formulation to varying extents. These variations will be discussed below.
Amino acid composition of the original acute tryptophan depletion, loading and control formulations of Young et al. 1

Amounts of amino acids are in g per a 100 g formulation. The * denotes that, these two amino acids are given in capsules due to their offensive odours and tastes.
For the ATPD test, two AA formulations have been used. In the first, designed by the group in Oxford (Sheehan et al), 21 only 7 amino acids were used, which were supplemented with Phe and Tyr in the control formulation. The AA composition of the ATPD test and its control as reported 21 is shown in Table 2. Because the original amounts of AA added up to only 45 g in the depletion- and to 57.5 g in the control, formulation, the AA amounts shown in Table 2 here have been adjusted to a total dose of 100 g.
Amino acid composition of the control and test formulations for acute tyrosine plus phenylalanine depletion (ATPD) by Sheehan et al. 21

In the original formulation, 21 the total amounts of amino acids were 57.5 g (control) and 45 g (ATPD). Amounts have been adjusted here to a total of 100 g for comparative purposes with other formulations.
In the other formulation for the ATPD test, reported by the Canadian group (Ellenbogen et al), 2 authors of the original ATD formulation, 1 the composition resembled more closely that used in the ATD and ATL tests, 1 namely containing the same 15 AA originally used. Thus, the compositions of the ATPD and its control formulation used by Leyton et al 28 based on those adapted for women by Ellenbogen et al 2 is shown in Table 3 below. Because the total amount of AA in the control formulation was only 85.5 g, the contents in Table 3 have been adjusted to a 100 g total for both the control and ATPD formulations, though the latter is short of the contents of Phe and Tyr. As was the case with ATD and ATL, other investigators have used the above two formulations for ATPD21,28 with minimal variations.

From the data in Tables 1–3, it is clear that there are 6 amino acids which must all be present in the control formulation, but that some of which must be absent in the depletion, or present in excess in the loading, formulation (namely Trp, Phe and Tyr). The remaining 3 amino acids always present in all formulations are the branched-chain amino acids (BCAA) valine (Val), leucine (Leu) and isoleucine (Ile). Their concentrations, sums and proportions in the various formulations are shown in Table 4 and their significance will be discussed below. As shown in Table 4, the content of BCAA as a proportion of the total amino acid content of the different formulations ranges between 29% and 33% in the formulations of the Canadian group,1,28 as opposed to the much larger content (48%–61%) in those of the Oxford group. 21
Contents of branched-chain amino acids (BCAA) in the original control and test formulations for acute tryptophan and tyrosine plus phenylalanine depletion.

Amounts of the 3 branched-chain amino acids and their sum (BCAA) are in g per the total content of amino acids in the formulations, from which the proportion of BCAA (as a %) is calculated.
Physiological and Behavioural Effects of the Formulations
The effects of the above formulations can be classified into those: 1) caused by the amino acid mixtures themselves; 2) resulting from modulation of levels of the three monoamines 5-HT, DA and NA. The amino acid formulations are generally unpalatable and induce changes varying between slight nausea and drowsiness to severe nausea and emesis,1,21,34 resulting in drop-out of study subjects. The latter authors 34 addressed the question of side effects during ATD and ATL at the 50 g and 100 g dose levels in detail, using a bodily symptom scale that assesses a variety of symptoms ranging from sweating, fast heart, shaking, dizziness, irritability, nausea, anxiety, depression, tension, headache and loss of appetite, to loss of concentration, tiredness and stomach ache. They found that, during ATD, there was no dose difference in the increases in the above side effects, which were highest in the last three symptoms above. During ATL, a clear dose difference was observed, with subjects receiving the 100 g dose showing significantly greater side effects. As a result of these side effects, these authors 34 reported attrition, which was confined to female subjects receiving the 100 g ATD or ATL dose, and, to avoid side effects and attrition and their effects on sample size and interpretation of behavioural data, these authors recommended the use of a 50 g dose in both ATD and ATL studies. Side effects after ATPD have not been studied in detail.
The effects of the above formulations resulting from modulation of central monoamines involve changes in mood, behaviour and cognition. The effects of ATD have recently been reviewed. 35 Briefly, in healthy subjects, ATD lowers mood in females, in those with a baseline depression score at the upper end of normal, or in subjects vulnerable to disturbances in central serotonin. By contrast, transient lowering of mood is a more consistent observation after ATD in patients with remitted depression. ATD also affects other behaviours, such as social behaviour, aggression and impulsivity, all of which may be closely connected to the serotonin system. ATD also affects a variety of cognitive processes in both healthy subjects and those with a serotonergic vulnerability. In particular, ATD impairs declarative episodic memory processes of delayed recall and memory consolidation, learning on visual discrimination and memory retrieval, episodic memory, stimulus reward learning and cognitive flexibility (for references, see 35 ). Much less work has been done on the effects of ATPD on mood, behaviour and cognition, most of which in healthy subjects. Thus, a study 36 of 12 healthy subjects suggested that ATPD does not alter mood, measures of memory, attention or behavioural inhibition. However, other studies in healthy subjects showed that ATPD increases vulnerability to lowered mood following exposure to aversive psychological events 37 and impairs affective processing 38 and spacial working memory. 29 In manic patients, Tyr depletion by administration of BCAA ameliorates manic symptoms. 39
Principles of the Tests
These tests are based on a number of biochemical, metabolic and physiological principles, which are set out below first for the role of Trp in serotonin synthesis, then that of Phe and Tyr in catecholamine synthesis.
Role of tryptophan in serotonin synthesis
As the rate-limiting enzyme of serotonin synthesis, Trp hydroxylase, is unsaturated with its Trp substrate,
40
brain [Trp] is the most important single metabolic determinant of the rate of serotonin synthesis
41
under acute short-term conditions. Although other important determinants include the rates of synthesis and degradation of Trp hydroxylase, its state of phosphorylation, the rate of firing of serotonergic neurons, and the influence of other neurotransmitters on this firing, it is unlikely that, under acute conditions, these determinants play a primary role in the rapid changes in serotonin synthesis observed during the ATD test. This is because of Trp hydroxylase having a relatively long half-life of 2–3 days, a slow axonal transport (~7 mm per day)
42
and its phosphorylation under optimal conditions enhances its activity by only 10%–30%.
43
Moreover, none of these factors can influence the rate of Trp hydroxylation
Role of tyrosine and phenylalanine in catecholamine synthesis
The role of Phe and Tyr in catecholamine synthesis is based on broadly similar principles. Thus, the rate-limiting enzyme of catecholamine synthesis, Tyr hydroxylase, is also partially saturated with its Tyr substrate, 40 though less so (~75% versus 50% for Trp hydroxylase). Tyr hydroxylase activity and hence the rate of catecholamine synthesis can therefore be influenced by changes in Tyr availability to the brain.58–63 This availability is also best expressed by the corresponding ratio, namely that of [Phe + Tyr]/[BCAA + Trp].
The potential effects of Tyr loading on catecholamine synthesis have received lesser attention, despite existing evidence. Thus, excess Tyr in the brain is not always reflected in enhanced catecholamine synthesis or turnover.40,57,64 This may be due to either feedback40,64 or substrate
59
inhibition of Tyr hydroxylase activity. The latter authors
59
showed that Tyr hydroxylase is activated
Poor Specificity of Current Amino Acid Formulations
Definitions of specificity
Specificity of the ATD or ATL test formulation implies that only the rate of serotonin synthesis will be decreased or increased respectively, with no change to the rate of dopamine or noradrenaline synthesis. Therefore one would expect that, whereas the [Free Trp]/[CAA] and [Total Trp]/[CAA] ratios will be either decreased (after ATD) or increased (after ATL), that of [Phe + Tyr]/[BCAA + Trp] should remain unaltered from the baseline value before intake of the AA formulations. By the same token, specificity of the ATPD or ATPL tests implies that, whereas the [Phe + Tyr]/[BCAA + Trp] ratio is either decreased (after ATPD) or increased (after ATPL), those of [Free Trp]/[CAA] and [Total Trp]/[CAA] should remain unaltered from baseline. As regards the control formulation(s) for the ATD, ATL, ATPD or ATPL tests, all three ratios should remain unaltered from baseline. In practice, existing AA formulations do not show this specificity and the following sections will demonstrate this defect and provide an explanation for it and a means of its rectification.
Previous demonstration of lack of specificity
Reilly et al 3 noted that many investigators using the ATD or its control formulation did not determine the free or total [Trp]/[CAA] ratio and only a few measured peripheral levels of Tyr or its ratio to other competing amino acids. While it must be assumed that the [Trp]/[CAA] ratio is decreased after ATD and increased after ATL, a decrease in this ratio has also been observed with a 100 g “balanced” control formulation for ATD containing the usual 2.3 g of Trp. 65 This ratio was also decreased in the control formulation if the Trp content was increased to 3.0 g, 66 but further increases to 4.1 g 67 or 4.6 g 68 of Trp caused elevations of this ratio. From data from our previous detailed pharmacokinetic and behavioural study comparing a 50 g with the traditional 100 g dose of the amino acid formulations for the ATD and ATL tests, 34 we reported 69 that intake of 50 g of the control formulation containing 1.15 g of Trp decreased the [Free Trp]/[CAA] ratio maximally by 61%.
We also found that the [Phe + Tyr]/[BCAA + Trp] ratio was decreased in the ATD, ATL and also in the control formulation by about 50%. Broadly similar decreases (40%–60%) in this latter ratio have been reported previously36,70,71 in control formulations used in the ATPD test, which are essentially similar to the control formulation for the ATD or ATL test. Furthermore, in the ATPD test, both the control and the Phe plus Tyr-deficient formulations are associated with decreases in the [Trp]/[CAA] ratio, of 30%–62% with the control formulation, and of 34%–96% with the depleting formulation.21,36,37,71–73
Reasons for lack of specificity
The main, if not only, reason for the above undesirable decreases in the [Phe + Tyr]/[BCAA + Trp] and [Trp]/[CAA] ratios in the control or the corresponding relevant depletion (or loading) formulations is the relatively larger contents of the three BCAA (i.e. Leu, Val and Ile), compared with those of Phe, Tyr and/or Trp, in the original Trp 1 or Tyr + Phe 21 formulation. The differences in contents of these 6 competitors is clearly reflected in the disproportionate increases in their plasma concentrations across the entire 7h time-course of administration of the 50 g control formulation in our previous study 34 illustrated here in Fig. 1, where the order of the absolute increases was as follows: Val > Leu > Ile > Trp > Phe > Tyr, whereas that of the increases relative to baseline was: Ile > Leu > Trp > Val > Tyr > Phe. In either case, Phe and Tyr were at the greatest disadvantage, followed by Trp. The decrease in the [Phe + Tyr]/[BCAA + Trp] ratio was even greater after ATL, because of the extra large increase in [Trp] that is combined with the high [BCAA]. 69
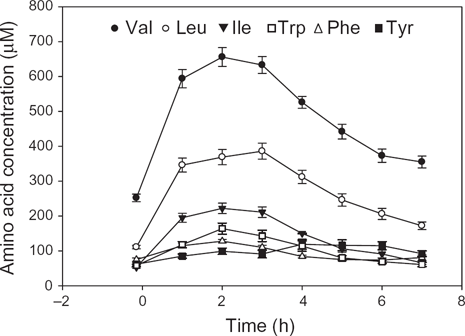
Time-course of changes in plasma competing amino acid conentrations following oral intake of a 50 g dose of the control formulation for the acute tryptophan depletion and loading tests.
Basis of the amino acid composition of the original formulations
In the original ATD, ATL and their control formulations, 1 the choice of an amino acid mixture with a composition based on human milk was a fortunate one in that it ensured a high level of the 3 BCAA Val, Leu and Ile to achieve a strong competition with Trp for entry into the brain. The presence of high concentrations of these 3 BCAA in human milk is of particular importance in human physiology, as BCAA, particularly Leu, are vital and essential for protein synthesis and hence the growth of the infant. 74 However, the use of a composition based on human milk as a control formulation is not necessarily the best choice for a human adult undergoing ATD or ATL studies, as its large content of BCAA is the cause of the decreases in the [Trp]/[CAA] and the [Phe + Tyr]/[BCAA + Trp] ratios in the control or the relevant depletion formulation. By the same token, in the original formulation for ATPD, 21 the content of the 3BCAA was even greater (48%–61% of the total AA content) than in the ATD-related formulations 1 (29%–33% of the total AA content), as illustrated in detail in Table 4 above.
Implications of the Lack of Specificity in Interpretation of Biochemical and Behavioural Data
Biochemical considerations
The above decreases in the [Trp]/[CAA] and [Phe + Tyr]/[BCAA+ Trp] ratios therefore suggest that 5-HT, DA and/or possibly NA synthesis could be inhibited by the control formulation and the corresponding ones for ATD, ATPD and their loading counterparts, an effect that could confound interpretation of behavioural changes (or lack of them). Whereas the role of brain Trp in serotonin synthesis is now well-established (see above), that of brain Tyr in catecholamine synthesis has received less attention in human studies, despite existing evidence. For example, the relationship between Tyr depletion and DA synthesis in various brain structures has been studied in detail in rats. Carlsson and Lindqvist
40
found that the rate of Tyr hydroxylation
The question then arises as to whether decreases in the above ratios of the order of 50%–60% leading to significant decreases in synthesis of 5-HT, DA and/or NA can alter monoamine function and thus influence behaviour. As regards DA function, the above decrease in DA synthesis 75 was accompanied by an even-stronger decrease in amphetamine-induced DA release. In a dose-finding study 71 in healthy adults, plasma levels of prolactin, a surrogate marker of DA function that is elevated when DA levels are decreased, were moderately elevated by a 10 g dose of BCAA, which decreased the [Tyr + Phe]/[BCAA] ratio by 70%. The [Trp]/[CAA] ratio was also decreased by this small dose of BCAA by 58% (the resulting decrease in 5-HT synthesis would be expected to cause a decrease in prolactin) and had this ratio decrease been avoided, the elevation of prolactin could have been greater. Also, it was found 76 that serotonin depletion induced by a Trp-deficient diet enhances amphetamine-induced DA release and causes a greater increase in motor activity than in controls. In a positron-emission tomographic study, 77 it was estimated that a modest decrease in brain [DA] of 10%–20% after ATPD in normal subjects can explain the 6% increase in [11C] raclopride binding, which results from DA receptors no longer being occupied by DA, and suggested that the decrease in the [Phe + Tyr]/[BCAA] ratio observed with the control formulation could have also increased this binding, relative to a control which did not decrease this ratio. It thus appears that DA function can be undermined by the changes in brain DA and the [Phe + Tyr]/[BCAA + Trp] ratio observed with the control formulations.
Behavioural considerations
In general, behaviour does not seem to be influenced in normal volunteers of either gender by the ATPD or its control formulation. 36 Behaviour is also not influenced by the control formulation for ATD or ATL in normal males or females, nor in males undergoing ATD: only ATD in normal females may influence behaviour.2,35 The situation in patient populations is, however, different, not only regarding ATD, which can precipitate a depressive episode in recovered depressed subjects, 13 but also the control formulation itself. Thus, e.g. in depressed patients, the free and/or total [Trp]/[CAA] ratio is already known to be decreased by 16%–36% relative to controls.78–84 Total [Trp] is also known to be decreased in depressed patients by 20%–29%.78,79,82,83,85 In 4 of the studies listed above which reported both parameters simultaneously,78,79,82,83 the decreases in the total [Trp] and total [Trp]/[CAA] ratio were generally comparable (respectively 26% and 23%; 26% and 23%; 20% and 18%; 29% and 32%). Thus, in depressed individuals, a further decrease of 50%–60% induced by the control formulation might lead to an even greater depletion of brain serotonin. Even if monoamine-dependent behaviour does not appear to be influenced when measured by existing instruments, investigators using a control formulation in the knowledge that it will decrease the Trp and Tyr ratios by ~50% in control subjects could not rule out a greater decrease in patient populations, which may precipitate significant behavioural changes. In fact, it has been suggested 86 that mood can be significantly altered when the decline in plasma [Trp] is >60%, a value that could be reached easily in patients receiving the control formulations in their present compositions.
The Need to Enhance the Specificity of the Control and other Amino Acid Formulations
We believe that: 1) it is inappropriate to argue that the use of a control formulation which alters the above ratios so significantly is permissible as long as it does not affect behaviour measured by existing instruments; 2) it would be both impractical and confusing to use two control formulations: one for normal subjects (whose behaviour may or may not be impaired) and the other for patient groups, who, by virtue of having a low or borderline [Trp]/[CAA] ratio are likely to experience greater ratio decreases and consequently behavioural changes; 3) it is more prudent to err on the side of accuracy and establish a truly balanced control formulation which does not alter these ratios.
Previous attempts to enhance the specificity of the control formulation for tryptophan depletion
Three previous attempts have been made to overcome the above ratio changes, but with only partial success. In the first, Weltzin et al 87 succeeded in maintaining the baseline [Trp]/[CAA] ratio by increasing the Trp content to 4.6 g/100 g of the traditional amino acid mixture. However, they did not measure the [Phe + Tyr]/[BCAA + Trp] ratio and it is almost certain that, with this level of Trp loading (which is 45% of that of the ATL dose), or even without it, this latter ratio will have been decreased. Booij et al 88 used a low dose ATD (25% of the normal one) as a control [based on a previous design by Krahn et al 89 ]. However, although this low-dose mixture did not alter the [Tyr]/[CAA] ratio, it still decreased the [Trp]/[CAA] ratio by 42%, against a 92% decrease by the full dose. However, interpretation of some, or all, of these biochemical changes is difficult because the subjects consumed a lunch during the test procedure. Still, while the use of a low-dose mixture may be useful in studies examining the effects of sub-optimal depletion of Trp and 5-HT, it cannot be considered an appropriate control for the ATD test dose. As regards the ATPD and its control formulation, no previous attempts have been made to address the issue of their specificity.
Biological and behavioural reasons for improving the specificity of the control formulations
The need for a truly balanced control or test formulation has already been emphasized. 3 Most investigators using the ATD or ATL (or the ATPD or ATPL) tests would agree that a “balanced” formulation should ensure that the control treatment maintains baseline values without altering the biochemical or behavioural parameters being studied, which would further enhance accurate interpretation of results. As BCAA play a pivotal role in the ATD test, the use of a balanced control formulation, rather than an amino-acid-free “neutral” placebo, is even-more important, particularly in relation to behavioural studies, because, apart from inducing a central 5-HT deficiency through Trp depletion and a central catecholamine deficiency through Phe and Tyr depletion, the BCAA have the potential to exert other equally important metabolic changes which could also further impact behaviour (for commentary, see Ref 74 ). Thus, in the human brain, BCAA are transaminated by branched-chain amino acid aminotransferase (BCAT) to branched-chain keto acids, converting in the process 2-oxoglutarate to glutamate.74,90 BCAA are thus nitrogen donors for the synthesis of the excitatory amino acid glutamate and the inhibitory neurotransmitter γ-aminobutyric acid (GABA), with Leu playing a particularly prominent role90–92 and it is noteworthy that the brain cytosolic isoenzyme of BCAT is located in GABAergic and glutamatergic neurons. 93 Since in humans, the brain normally accounts for 10%–20% of their total body metabolism, 94 a significant increase could be expected after loading with BCAA, as during the depletion or loading tests, resulting in enhanced synthesis of glutamate and GABA. Changes in glutamatergic and/or GABAergic neurotransmission are therefore expected under these conditions, which could impact on 5-HT and dopamine functions, with the potential to modulate behaviours associated with these cerebral monoamines.
Strategies for Enhancing the Specificity of the Control and Test Formulations for Acute Tryptophan and Tyrosine plus Phenylalanine Depletion and Loading
In the light of this and the preceding discussion, and based on theoretical graphs designed to maintain normal [Trp]/[CAA] and [Phe + Tyr]/[BCAA + Trp] ratios under the depletion, loading, or balanced (control) condition, if the contents of the [BCAA] or of [Phe + Tyr] were to be altered independently, we proposed 69 two strategies for normalizing these ratios: (1) decreasing the contents of the three BCAA by ~30%; or (2) increasing those of Phe and Tyr by ~50%. Of these, the first is the preferred strategy, because it avoids the metabolic consequences of Phe and Tyr loading associated with the latter strategy, because, if adopted, it could contribute to a further lowering of both the free and total [Trp]/[CAA] ratios in the control formulation. A third strategy, applicable only to the control formulation, is to increase the Trp content, as performed previously. 87 However, while this may improve the [Trp]/[CAA] ratio, it can only further decrease the [Phe + Tyr]/[BCAA + Trp] ratio and thus lead to a greater depletion of brain catecholamines. As BCAA appear to be the components of amino acid mixtures responsible for the undesirable decreases in the above ratios, we believe that modulating their content in the various formulations is the most appropriate strategy for enhancing the specificity of these important research and diagnostic tests.
Conclusions and Comments
We hope that this review has addressed the important biochemical principles underpinning the acute Trp and Tyr plus Phe depletion and loading tests, illustrated clearly the poor specificity of the various test and control amino acid formulations and made constructive proposals for improving their specificity. In the accompanying paper, we successfully demonstrate the normalisation of the free and total [Trp]/[CAA] and [Phe + Tyr]/[BCAA + Trp] ratios in a new balanced control formulation for all the above tests by decreasing the content of BCAA, and propose adjustments of the contents of the active formulations to ensure their specificities.
Disclosures
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers of this paper report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material.
Footnotes
Acknowledgements
Work reviewed from our respective laboratories was supported by grants from the Wellcome Trust (069301) and the NIH (R01-AA012046, RO1-AA014988 and T32-AA07565).