
Abstract
Background:
The serendipitous discovery of heavy-chain antibodies devoid of light chains in camelids and the subsequent development of VHHs (variable region of camelid heavy chain) have provided a very important tool for research and possibly for therapeutics. In this study, we synthesized single-domain 15-kDa antibody SBT-100 (anti-STAT3 B VHH13) against human STAT3 (signal transducer and activator of transcription) that binds selectively to STAT3 and suppresses the function of phosphorylated STAT3 (p-STAT3).
Methods:
Single-chain VHH nanobodies were generated by immunizing camelid with humanized STAT3. Commercially available breast cancer cell lines including MDA-MB-231, MDA-MB-468, MDA-MB-453, MCF-7, and BT474 were used. Cell proliferation was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. The association of anti-STAT3 B VHH13 with STAT3 and p-STAT3 was determined by immunoprecipitation and Western blot analyses. The efficacy of SBT-100 on the growth of MDA-MB-231 xenografts in vivo was determined using athymic mice. Statistical significance for cell proliferation was determined using analysis of variance. If a significant difference (P < .05) was observed, then Tukey-Kramer multiple comparison test was conducted.
Results:
SBT-100 suppressed cell proliferation of triple-negative breast cancer cells (P < .01) as well as provided significant inhibition of tumor growth (P < .05) in a xenograft model without any toxicity. Results are presented to show that anti-STAT3 B VHH13 selectively binds to STAT3 suggesting that the effects were mediated by inhibiting STAT3.
Conclusions:
A very large number of human malignancies and benign diseases have constitutive STAT3 activation. Therefore, the results described here suggest that anti-STAT3 B VHH13 can be developed for therapeutic intervention for cancer cells expressing STAT3 or p-STAT3.
Keywords
Introduction
The discovery of camelids having antibodies that consist only of heavy chains and devoid of light chains 1 eventually led to the development of the variable region (VHH) of this antibody to bind targets. These are the smallest independent proteins that bind antigen similar to the traditional 4-chain antibodies. 2 VHHs, as shown in Figure 1A, also have unique properties such as its small size, approximately 15 kDa, elongated complementarity-determining region, ease of production, and durability. 3 VHHs have shown good safety in humans and efficacy in treating patients in multiple clinical trials. 4
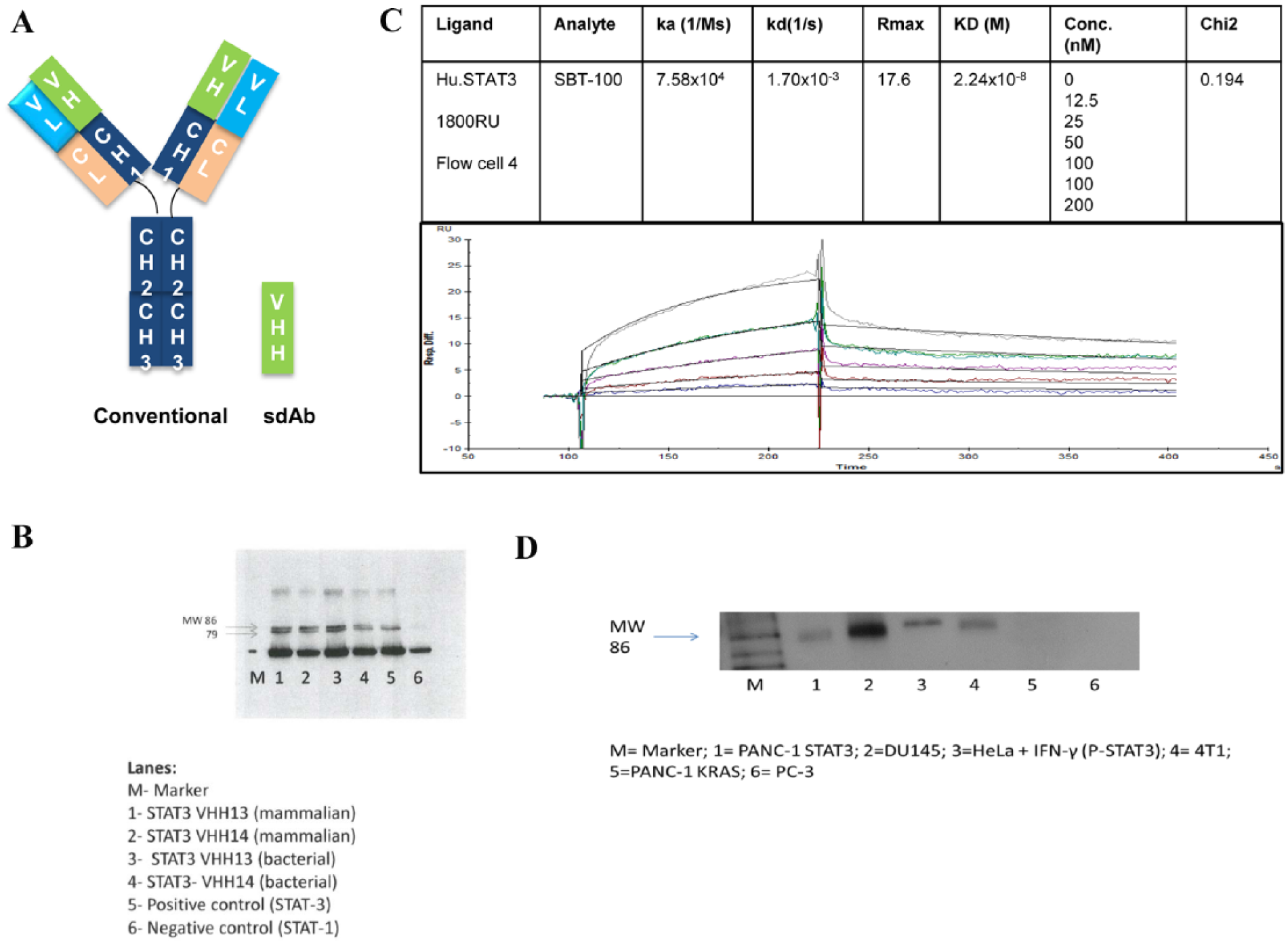
Binding of STAT3 with VHH13 generated in bacteria and VHH14 derived from mammalian source. (A) Schematic comparison of conventional antibodies and small-molecular-weight camelid antibodies devoid of light chain comprising heavy chain with variable region (VHH). (B) The kinetics profiles of bacterial VHH13-human STAT3. Binding of human STAT3 with anti-STAT3 B VHH was determined using Biacore 3000. For scouting, the sample was allowed to flow over the chip and the binding of sample to the ligand was monitored in real time. The affinity constant (KD) was determined as a ratio of dissociation and association rates. (C) Western blot showing the immunoprecipitation of various STAT3 VHHs to MDA-MB-231 cells using Dynabeads coupled with STAT3 VHHs, commercially available STAT3 was used as a positive control, and STAT1 as a negative control. The band densities were determined using UN-SCAN-IT software (Silk Scientific Inc., Orem, UT, USA). (D) Comparative expression of STAT3 in breast (MDA-MB-231, 4T1), pancreatic (Panc-1), and prostate (DU145) cancer cells and HeLa cells. Western blot showing the immunoprecipitation of bacterial STAT3 VHH13 to cell lysates from breast, pancreatic and prostate cancer cells as well as HeLa + IFN-γ. For this, cell lysate was incubated with Dynabeads coupled with STAT3 VHH13.
The use of humanized monoclonal antibodies for therapeutic purposes for many diseases has made a significant and positive impact in patient care. For example, trastuzumab and bevacizumab are used for treating patients with cancer. However, because of their large size, approximately 150 kDa, conventional heterotetrameric antibodies cannot access intracellular targets. The VHHs, however, are about one-tenth the size (15 kDa) of the conventional heterotetrameric antibodies. This much smaller size appears to allow VHH to access intracellular targets without the need for conjugation to another molecule 5 or the need for a facilitated transport into the cytoplasm across the cell membrane.
Nearly 70% to 80% of all human malignancies as diverse as breast cancer, pancreatic cancer, leukemia, lymphomas, sarcomas, and multiple myelomas exhibit constitutive expression of activated STAT3.6–17 Signal transducer and activator of transcription 3 (STAT3) is a transcription factor and one of the 7 STAT proteins. It is encoded by the STAT3 gene and is highly conserved in evolution. In response to cytokines, growth factors, cytoplasmic kinases, and cell membrane receptor tyrosine kinases, STAT3 is phosphorylated. This activation produces P-STAT3 which then can form homodimers or heterodimers and then these dimers translocate into the nucleus. There the P-STAT3 dimer can transcribe a large number of genes important for cell growth, proliferation, differentiation, and immune response. Therefore, we used the beneficial properties of VHH to target intracellular STAT3. By immunizing a camelid with human STAT3, we generated anti-STAT3 VHHs and determined its efficacy against the proliferation of human breast cancer cells including steroid receptor–positive and triple-negative breast cancer (TNBC) cells. The effects of anti-STAT3 VHH on the growth of MDA-MB-231 xenografts in athymic mice were also evaluated.
Methods
VHH generation
Recombinant full-length human STAT3 with a GST tag fused to its N-terminal (STAT3-1496H) was provided by Creative BioMart (Shirley, NY, USA). VHH discovery was achieved. Briefly, a camel (Camelus bactrianus) was used for immunization with the recombinant human STAT3. After the immunization protocol was completed, peripheral blood cells were collected for total RNA purification. Total RNA was isolated from each of the bleeds using TriZol reagent. The RNA was immediately used to prepare first strand complementary DNA (cDNA) using RNAsin (Promega) and Super RT (HT Biotechnology Ltd, Cambridge, UK). Reverse transcription was conducted using oligo-dT primers. Single-stranded cDNA was briefly centrifuged and stored at −20°C. For the first round polymerase chain reaction (PCR), after quantification of cDNA by UV spectrometry, VH-CH1-FC/VHH-FC gene was amplified; FC downstream primer and VH/VHH-FR1 upstream primer were used with 200× diluted cDNA as the template. The PCR product was collected and purified. For the second round PCR, VHH-FR4 downstream primer and VHH-FR1 upstream primer were used to amplify VHH gene. The second PCR product and pCDisplay-3M phagemid from Creative Biolabs (Shirley, NY, USA) were then digested with restriction endonucleases, BssHII and NheI, simultaneously and purified. The DNA fragments were ligated using NEB T4 DNA ligase. The ligation products were desalted before being electrotransformed into TG1 host cells. In the end, a VHH library was constructed successfully with a size of 109 diversity. In biopanning, enzyme-linked immunosorbent assay (ELISA) plate wells were coated with 5 to 30 μg/mL STAT3 protein at 4°C for 2 hours. Wells were washed 3 times and blocked at 4°C overnight. The original library phages were added in preblocked wells (noncoated) and incubated at room temperature (RT) for 30 minutes to subtract background. Then, subtracted phages were transferred into preblocked and coated wells and incubated at RT for 1 hour. Wells were washed 10 times and bound phages were eluted by trypsin digestion, followed by titration and amplification of eluate. Positive binder clones were confirmed with phage ELISA. The plasmids for positive clones from bacterial cells were isolated using a Qiagen Mini-prep kit according to the manufacturer’s instructions. DNA sequencing of the VHH binder clones was conducted using “L1” and “S6” as the primers.
Two clones that demonstrated strongest binding to the target recombinant human STAT3 were selected for further characterization. These 2 clones, VHH13 and VHH14, were produced in 2 different expression systems: one was in an Escherichia coli system and the other was in an HEK 293 cell system. Those VHHs produced in the E coli system were called anti-STAT3 B VHH13 and anti-STAT3 B VHH14. Those VHHs produced in HEK cells were called anti-STAT3 H VHH13 and anti-STAT3 H VHH14.
For bacterial expression, expression constructs were transformed into E coli BL21 (DE3) Star competent cells. Following sonification of the isopropyl β-
Cell lines
The human breast cancer cell lines (MCF-7, BT474, and MDA-MB-231, MDA-MB-468, and MDA-MB-453) were obtained from American Type Culture Collection (Manassas, VA, USA). Cells were grown in Minimum Essential Medium (MEM; Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Atlanta Biologicals, Flowery Branch, GA, USA) and penicillin-streptomycin-glutamine (Life Technologies). All cells were grown in the presence of 5% CO2 at 37°C in an incubator.
Animals
Athymic nude-Foxn1nu female mice aged 5 to 6 weeks were purchased from Envigo Laboratories (Indianapolis, IN, USA). Animals were quarantined for 1 week and housed 5 mice per cage, with a 12-hour light-dark cycle and a relative humidity of 50%. Drinking water and diet were supplied to the animals ad libitum. All animals were housed under pathogen-free conditions and experiments were performed in accordance with the IIT Research Institute Animal Use and Care Committee.
Immunoprecipitation and Western blot analysis
Cancer cells were grown to 50% to 70% confluence and then were disrupted in freshly prepared ice-cold lysis buffer (20 mM HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid], pH 7.9, 400 mM NaCl, 0.1% NP-40, 10% glycerol, 1 mM sodium vanadate, 1 mM sodium fluoride, 1 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride, 10 μg/mL aprotinin, and 10 μg/mL leupeptin for 45 minutes on ice. Lysates were then centrifuged, the supernatant was collected, and protein concentration was determined using a modified Lowry method (Bio-Rad, Hercules, CA, USA). Total protein (1 mg) was incubated with 1.5 mg of Dynabeads (Invitrogen, Carlsbad, CA, USA) containing 1 of 4 VHHs against STAT3 (Creative Biolabs), a positive control (STAT3, cat#SC-482; Santa Cruz Biotechnology, Dallas, TX, USA), or negative control (STAT1, cat# 9172; Cell Signaling, Danvers, MA, USA) for 1 hour at 4°C. Beads were then washed. Following the final wash, 60 μL of lysis buffer was added, and the resulting supernatant was subjected to Western blot analysis. Briefly, samples were separated on 10% polyacrylamide gels and transferred to a nitrocellulose membrane. The membranes were blocked and then incubated with anti-STAT3 antibody (Cell Signaling, Cat# 4904). The chemiluminescence reaction was performed using the ECL system from Santa Cruz Biotechnology.
MTT assay
Cancer cells were grown to 90% confluence. Then, they were washed, trypsinized, and counted using a Coulter Counter (Beckman, Brea, CA, USA). The proliferation studies were conducted using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. These cells were seeded in a 96-well plate at a density of 5 × 10E3 per well as indicated by the manufacturer (Roche Diagnostics Corporation, Indianapolis, IN, USA). Cells were allowed to adhere for 24 hours and then the VHHs were added at the appropriate concentrations (ie, 0, 0.5, 1.0, 10.0, or 100 µg/mL). At the time of termination (day 3), 10 μL of MTT reagent (0.5 mg/mL) was added to each well as indicated by the manufacturer. After a 4-hour incubation period, 100 μL of solubilization solution was added and the plate was placed in the incubator overnight. All the plates were read at 570 nm wavelength using the BioTek plate reader (Winooski, VT, USA). Statistical analyses of all data were analyzed using GraphPad InStat 3 (GraphPad Software, Inc., La Jolla, CA, USA). Treatment groups were compared with vehicle control group using 1-way analysis of variance. If a significant difference (P < .05) was observed, then Tukey-Kramer multiple comparison test was conducted.
Xenografts
MDA-MB-231 cells at a density of 5 × 10E6 were injected subcutaneously into the right flank at a final volume of 100 μL of MEM media. Tumor measurements were initiated as soon as the tumors were palpable. Thereafter, tumors were measured twice weekly. Animals were randomized when tumors reached a size range of 55 to 150 mm3 using the stratified random sampling algorithm. Treatment (anti-STAT3 B VHH13) or vehicle (phosphate buffred saline [PBS]) was initiated the day following randomization as described above and continued for 14 days.
Results and Discussion
To test the specificity of anti-STAT3 VHHs, we used cell lysates of the human breast cancer cell line MDA-MB-231, which constitutively expresses activated STAT3. Endogenous STAT3 immunoprecipitated with the anti-STAT3 B VHH13 and anti-STAT3 VHH14 produced in E coli and in HEK cells were tested at different concentrations depending on the stability and affinity of the conjugates. As shown in Figure 1B, the VHHs in lanes 1, 2, and 3 bound particularly well to the endogenous STAT3 protein, and this binding exhibits increased expression as compared with lane 5, which used a polyclonal anti-STAT3 traditional 4-chain antibody. Both mammalian and bacterial VHH13 and VHH14 bound with STAT3. These results also clearly suggest that the newly synthesized VHH is more efficient than the conventional antibodies. The negative control, a polyclonal anti-STAT1 antibody, showed no binding to the endogenous STAT3 from the MDA-MB-231 cell lysate. Furthermore, the kinetics of bacterial anti-STAT3 B VHH13 binding to human STAT3 were determined. The experiments were performed on Biacore 3000 at 25°C. Here, we used 1800 RU (resonance unit) was immobilized on the chip. Although it appears to be high, the amount of binding that we measured was so low; we had to use higher RUs for the captured ligand. When we used 400 RU, we could not detect binding. This is consistent with our being able to detect only 8% or so of the Rmax bound. Furthermore, we calculated the kinetic parameters with and without correction for mass transfer and found no difference. The unoccupied sites were quenched with 1 M ethanolamine. Binding of sample to the ligand was monitored in real time. The affinity constant (KD = kd/ka) was calculated from the observed on-rate (ka). As shown in Figure 1C, human STAT3 bound to anti-STAT3 B VHH13 with high affinity. The KD was calculated to be 2.2 × 10−8 M. The results suggest that kon is average, but not poor, and koff is fast in the beginning of the dissociation curve and then slows down; this may be because of nonspecific binding of impurities that come off first and then the actual analyte. The curve fitting results indicated a low χ2 value indicating that the curves obtained are significant and the duplicate values were in excellent agreements.
Next, we examined whether other cancer cells expressing constitutively activated STAT3 bind to anti-STAT3 B VHH. These results are shown in Figure 1D. Results show that endogenous STAT3 immunoprecipitated in the constitutively activated STAT3 cell lines expressing p-STAT3, namely, PANC-1, DU145, and 4T1. No bands were noted for either negative control (ie, PANC-1 probed with KRAS VHH and for PC-3 a STAT3-null cell line). From these results it can be concluded that anti-STAT3 B VHH binds with high affinity to p-STAT3 from cell lines that have constitutively activated STAT3.
Once we showed that the anti-STAT3 VHHs had good binding to endogenous STAT3 in MDA-MB-231, we determined the effects of SBT-100 on proliferation of MDA-MB-231 cells. The results from the MTT assays are shown in Figure 2A. The results showed that there was a significant inhibition of cell proliferation for both mammalian and bacterial VHHs at 10 and 100 μg/mL concentrations. The anti-STAT3 bacterial VHH13 gave the highest degree of inhibition at both 10 and 100 μg/mL concentrations. Therefore, based on the MTT experiments, bacterial VHH13 was selected for further efficacy studies with other breast cancer cell lines. We determined the efficacy of SBT-100 against 4 breast cancer cell lines including MCF-7 (ER+PR+HER2+), BT474 (HER2 amplified), MDA-MB-453 (TNBC), and MDA-MB-468 (TNBC). Results showed that all the cell lines except MDA-MB-453 exhibited 80% growth suppression, whereas MDA-MB-453 was less responsive (Figure 2A). Interestingly, MDA-MB-453 has been used as a negative control for p-STAT3 since it is a cell line without p-STAT3 and thus this cell line does not respond to small molecule inhibitors of STAT3 (6). In addition, IC50 values were determined for MCF-7, BT474, MDA-MB-231, MDA-MB-468, and MDA-MB-453 by incubating cells for 3 days with increasing concentrations of anti-STAT3 B VHH in the range of 10 to 100 μg/mL. Results are shown in Figure 2B. The IC50 for various cell lines ranged from 5.11 to 25.2 μg/mL, which was not significantly different among these cells.

Effects of anti-STAT3 B VHH on the proliferation of breast cancer cell lines. (A) Steroid receptor–positive and negative breast cancer cells were incubated with increasing concentrations of anti-STAT3 B VHH. The cell proliferation was determined by MTT assays as described in the “Methods” section. (B) IC50 determination of ST effects for all the cell lines. The IC50 was determined using the IC50 Tool Kit (http://www.ic50.tk/). MTT indicates 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Next, the activity of anti-STAT3 B VHH was evaluated using the MDA-MB-231 human breast cancer xenograft model. Briefly, animals were inoculated with MDA-MB-231 cells at a density of 5 × 10E6 and then tumors were allowed to grow until a stage size of 50 to 150 mm3. The animals were then randomized into 4 groups. The dosing schedule was as follows: group 1 received PBS intraperitoneal twice daily for 14 days; group 2 received 1 mg/kg intraperitoneal twice daily for 14 days; group 3 received 2 mg/kg intraperitoneal twice daily; and group 4 received 2 mg/kg intraperitoneal once a day for 14 days. A postdosing observation period of 7 days followed at the end of the 14 days of VHH treatment. At the time of termination (day 42), the tumors in the SBT-100–treated mice were smaller in size as compared with vehicle control (Figure 3A). The mean growth inhibition in the treatment groups was 78.3%, 75.2%, and 55.9% for groups 2, 3, and 4, respectively (Figure 3B).

Effects of anti-STAT3 B VHH in MDA-MB231 xenografts in vivo. (A) Effects of SBT-100 on the tumor size after transplantation of MDA-MB231 in athymic mice treated with either vehicle or anti-STAT3 B VHH. (B) Growth curves of the MDA-MB231 tumors in athymic mice treated with either 1 or 2 mg/kg anti-STAT3 B VHH.
The target of VHH is an intracellular transcription factor and not an extracellular receptor. Here, we have shown that a VHH can freely cross the cell membrane to target an intracellular protein such as STAT3. Anti-STAT3 B VHH can bind its target protein with high affinity and as a result can significantly inhibit cancer growth in vitro within 3 days. Cancers with constitutively activated STAT3 can also be significantly inhibited in its growth with anti-STAT3 B VHH. Tumor regression in the treatment groups persisted for 7 days after discontinuation of the treatment. Moreover, administration of anti-STAT3 B VHH more than 14 days showed no clinically observable toxicity (eg, diarrhea, hair loss, tremors, or failure to thrive) or significant weight loss in multiple xenograft experiments. These animals were further maintained for additional 28 days and body weights as well as survival were determined. Therefore, these toxicity studies were in fact conducted for 42 days. This may be the first demonstration of a VHH freely crossing the cell membrane that results in therapeutic cancer growth suppression in vitro and in vivo by targeting STAT3 with a VHH.
Many small-molecule inhibitors of STAT3 cause cancer growth inhibition by inhibiting phosphorylation of STAT3, inhibiting dimerization of p-STAT3, or inhibiting DNA binding of p-STAT3 dimers. The anti-STAT3 VHH described here binds both unphosphorylated STAT3 and phosphorylated STAT3. This may make the anti-STAT3 VHH more effective in inhibiting STAT3 function than small-molecule inhibitors because it can bind STAT3 in at least 2 of its forms. It has been described that unphosphorylated STAT3 can target many of the same genes as p-STAT3 dimers. 18 VHHs have shown excellent safety in human patients in multiple clinical trials, 4 and inhibiting STAT3 in humans has also been shown to be safe. 17 This is in contrast to many small-molecule STAT3 inhibitors which have shown significant toxicity or have not shown efficacy in vivo.
There are many groups that have used intracellular expression of VHHs as intrabodies to demonstrate enzyme inhibition or cancer growth inhibition. 19 These models have inserted the gene for the VHH into the DNA of the host cell being tested. In preliminary studies, we and others have shown that VHH can freely cross cell membranes. Moreover, we recently showed that VHHs also cross the blood-brain barrier (unpublished data). If that indeed is true, then the VHH can provide a broader application in cancer chemotherapeutics. Based on the present studies indicating that SBT-100 suppresses the functions of p-STAT3, efforts are underway to develop this antibody for the clinical trials. To the best of our knowledge, this is the first demonstration of targeting an intracellular protein (ie, STAT3) with a single-domain antibody, such as VHH, and demonstrating a therapeutic effect. Being able to target STAT3 with a VHH appears to be a way to treat malignant diseases with efficacy and safety. The overall potential for this suppression of activated STAT3 is summarized in Figure 4. Because activated STAT3 is involved in the signaling pathways of many targets, the inhibitors of activated STAT3 such as VHH may have broad clinical significance.

Schematic diagram of the cell signaling pathways that are affected by activated STAT3. It is proposed that many of these pathways interacting with activated STAT3 can be suppressed by anti-STAT3 B VHH.
Conclusions
This is the first report that provides evidence for the efficacy of activated STAT3 nanobodies in TNBC. Because many human malignancies have constitutive STAT3 activation, the results described here suggest that anti-STAT3 B VHH can be developed for therapeutic intervention for cancer cells expressing STAT3 or p-STAT3.
Footnotes
Acknowledgements
The authors thank Mandar Mali and Karen Castillo for their competent technical assistance and precision antibodies, Columbia, MD for the STAT3-VHH binding studies.
Funding:
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was supported by Singh Biotechnology LLC.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SS contributed to concept development, experimental design, interpretation of the results, manuscript writing, and funding for the project; GM to study design, conducting experiments, and manuscript writing; DC to synthesis of VHH; ASP to interpretation of results and statistical analyses; and RM to experimental design, interpretation of results, and manuscript writing.