
Abstract
Background:
Intestinal metaplasia (IM) of gastric epithelium has traditionally been regarded as an irreversible stage in the process of the Correa cascade. Exploring the potential molecular mechanism of IM is significant for effective gastric cancer prevention.
Methods:
The GSE78523 dataset, obtained from the Gene Expression Omnibus (GEO) database, was analyzed using RStudio software to identify the differently expressed genes (DEGs) between IM tissues and normal gastric epithelial tissues. Subsequently, gene ontology (GO) analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis, Gene Set Enrichment Analysis (GESA), and protein-protein interaction (PPI) analysis were used to find potential genes. Additionally, the screened genes were analyzed for clinical, immunological, and genetic correlation aspects using single gene clinical correlation analysis (UALCAN), Tumor–Immune System Interactions Database (TISIDB), and validated through western blot experiments.
Results:
Enrichment analysis showed that the lipid metabolic pathway was significantly associated with IM tissues and the apolipoprotein B (APOB) gene was identified in the subsequent analysis. Experiment results and correlation analysis showed that the expression of APOB was higher in IM tissues than in normal tissues. This elevated expression of APOB was also found to be associated with the expression levels of hepatocyte nuclear factor 4A (HNF4A) gene. HNF4A was also found to be associated with immune cell infiltration to gastric cancer and was linked to the prognosis of gastric cancer patients. Moreover, HNF4A was also highly expressed in both IM tissues and gastric cancer cells.
Conclusion:
Our findings indicate that HNF4A regulates the microenvironment of lipid metabolism in IM tissues by targeting APOB. Higher expression of HNF4A tends to lead to a worse prognosis in gastric cancer patients implying it may serve as a predictive indicator for the progression from IM to gastric cancer.
Introduction
In the classical Correa cascade process, intestinal metaplasia (IM) is considered a precancerous lesion of gastric cancer. Despite numerous studies conducted over the past decades, the underlying molecular mechanisms have remained elusive. 1 In gastritis, related markers (LGR5, SI, MME, etc.) of intestinal stem cells can be induced by the caudal-related homeobox1,2 (CDX1, 2). These genes play a regulatory role in determining intestinal phenotypes by impacting relevant signaling pathways such as NFkB and Wnt/β-catenin.2 -7 Moreover, a clinical correlation analyses also showed that the development of gastric macula with foam cells as the main pathological change was closely related to the IM. 8 This finding suggests that gastric macula with foam cells may serve as a potential risk factor for the development of gastric cancer.9,10
In recent years, numerous studies demonstrated that fatty acids (FAs) are necessary for the tumor metabolism and the maintenance of tumor stem cells, which can also activate related pathways to promote tumor invasion.11 -15 Reprogramming of lipid metabolism in tumors is characterized by enhanced lipid uptake and oxidative capacity. The similar situation may also exist in the procession of IM to gastric cancer. However, few studies have revealed the underlying mechanism between IM and lipid metabolism.
As a highly conserved member of the nuclear receptor family, 16 hepatocyte nuclear factor 4A (HNF4A) plays a vital role in metabolic homeostasis, and the progression of many malignancies.17,18 As a transcription factor, HNF4A promotes the expression of B cell receptor associated protein 31 (BAP31), which has immune-related functions. The upregulation of BAP31 not only facilitates the growth of GC cells and promotes G1/s transition, but also reduces the sensitivity of gastric cancer to chemotherapy. 19 Furthermore, HNF4A has been regarded as a potential regulator in the upstream of WNT signaling pathway, and it was found that HNF4A may regulate “metabolic switch” characteristic of a general malignant phenotype. 20 A study utilized mathematical algorithms to large-scale DNA methylation and transcriptome profiles to reconstruct transcription factor (TF) networks reveals that HNF4A can promoted the proliferation and survival of cancer cells by transcriptional activation of numerous downstream targets. 21 Our findings suggest that HNF4A could potentially serve as a bridge connecting intestinal metaplasia (IM) and gastric cancer through the regulation of lipid metabolic pathways. This proposed mechanism offers a novel perspective to explain the transition from IM to gastric cancer.
Materials and Methods
Drug intervention: BI6015 (T21867, TargetMol, USA) was used as a specific inhibitor of HNF4A, and the corresponding concentration gradient was established by the recommended guideline. The drug intervention lasted for 24 hours, and then the total protein of cells was extracted and verified by WB experiment.
Results
Identification of key genes
Firstly, we divide the samples of GES78523 dataset into two groups: the first group consisted of completely IM tissues and normal tissues, while the second group comprised of incomplete IM tissues and normal tissues. In order to find the similarities and differences between the two groups. We compared the differences of genes between the two groups. Moreover, in order to improve the representativeness of the selected genes, we raised the threshold accordingly (| logFC |> 4, P < .01). Finally, 26 differential genes were screened in the first group, and 16 differential genes were screened in the second group. After intersection analysis, 15 hub genes (FABP1, MUC2, MUC3A, MUC17, CLCA1, CPS1, MTTP, CLDN7, SI, APOB, OLFM4, ALDOB, ANPEP, REG4, DMBT1) were screened in both groups (Figure 1a). Then, the enrichment analysis of GO and KEGG on these genes were performed. The result of GO enrichment analysis showed that these genes were significantly enriched in very-low-density lipoprotein particle assembly, triglyceride metabolic process, low-density lipoprotein particle remodeling, lipoprotein transport, lipoprotein localization, acylglycerol metabolic process, and neutral lipid metabolic process. And the result of KEGG enrichment analysis demonstrated that these genes were significantly related to fat digestion and absorption pathways (Figure 1c and d). To further investigate the correlation of these genes, the protein-protein interaction (PPI) analysis was performed. Moreover, the MCODE tool of Cytoscape software was used to analyze the gene enrichment score. The analysis revealed that the gene set where APOB is located gained the highest score. These results showed that APOB occupies the most important position in PPI network, so we speculated that APOB may play a very important role in IM (Figure 1b). According to the above bioinformatics analysis results, the western blot analysis of protein samples showed that the expression of APOB in IM tissues was significantly higher than that in normal gastric epithelial tissues (Figure 1e). In order to investigate the potential regulatory mechanisms underlying the expression of APOB, the hTFtarget website was used to identify transcription factors associated with APOB. Finally, the gene of HNF4A was identified. Similarly, HNF4A was also significantly overexpressed in IM tissues in our western blot results (Figure 1e). In addition, our results showed that there is a significant positive correlation between APOB and HNF4A in most tumors, including gastric cancer (Figure 1f). Therefore, we speculate that HNF4A and APOB play a vital role in IM tissues and potentially contribute to the development of gastric cancer.
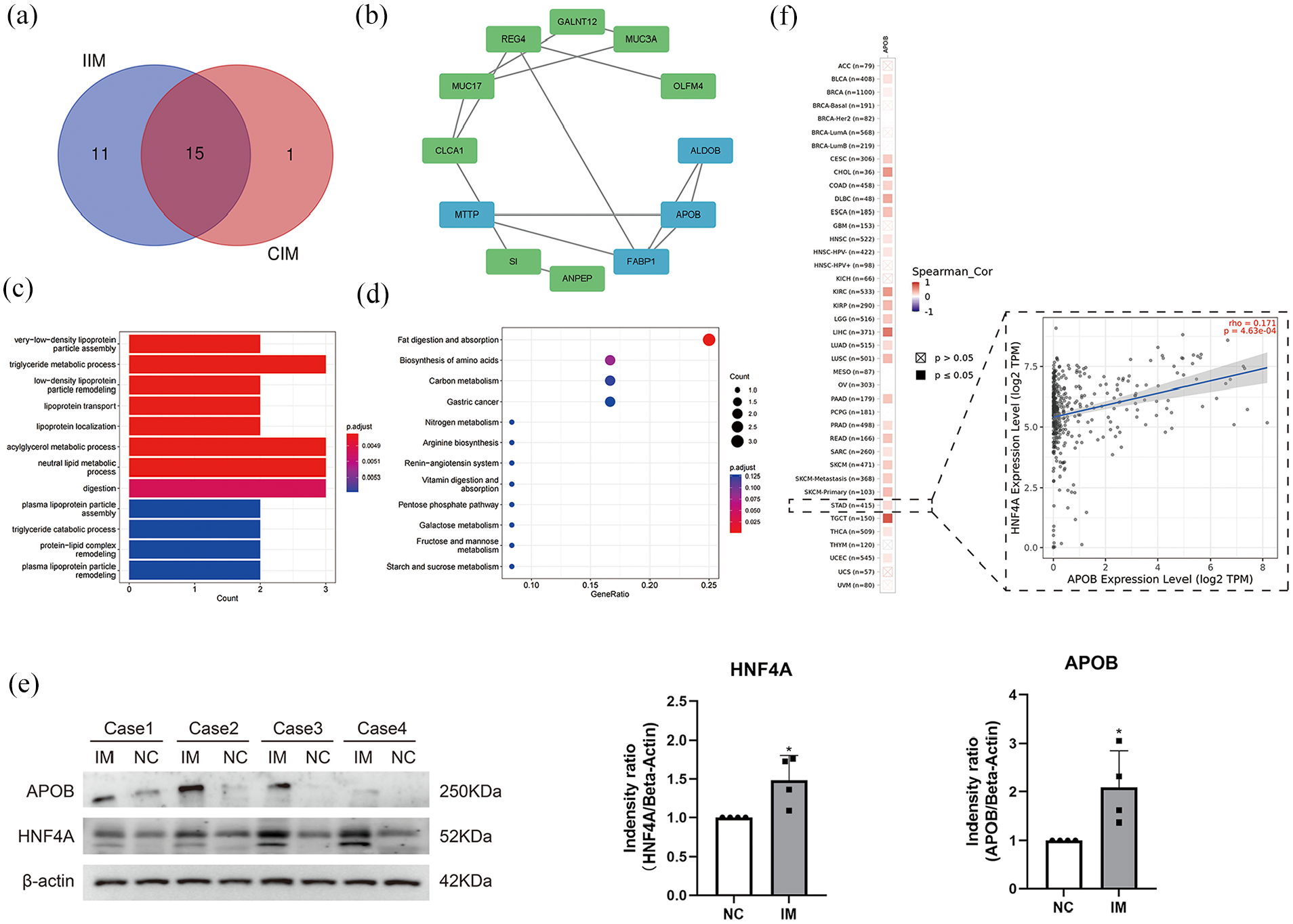
Identification of key genes: (a) Venn diagram showing the intersection of DEGs between the complete IM and incomplete IM groups (|logFC| > 4, P < .01). (b) PPI analysis: the blue part is the set of genes with the highest enrichment screened by the MCODE tool. (c) Functional enrichment analysis shows that the intersecting genes are mainly enriched in lipid metabolism-related functions. (d) KEGG enrichment analysis shows that the intersecting genes are mainly enriched in lipid digestion and uptake-related pathways. (e) The expression levels of APOB and HNF4A in IM and normal gastric epithelial tissues were detected by western blot while NC was the normal control and IM was the corresponding IM tissue. All experiments were repeated three times, and P < .05 was considered statistically significant. (f) Correlation analysis of APOB and HNF4A in various malignancies (red represents positive correlation, blue represents negative correlation).
Expression analysis of key genes
To further investigate the relationship between HNF4A and APOB and its biological function in the progression of IM to gastric cancer, the TCGA database was used for pan-cancer analysis. The results showed that HNF4A was significantly increased in gastric cancer (Figure 2a). In addition, the HNF4A expression level was higher in gastric cancer than in most other malignancies (Figure 2b). Similarly, the WB results indicated a substantial increase in the expression of HNF4A in gastric cancer cell lines (MGC 803 and HGC 27) when compared to normal gastric epithelial cells (GES-1). However, there were no significant differences in APOB expression between gastric cancer cells and normal gastric epithelial cells, although the expression level of APOB in gastric cancer exhibited an upward trend (Figure 2c-e). To further validate the relationship between APOB and HNF4A, the follow-up experiment was conducted using HGC 27 cells that exhibited relatively high APOB expression. Firstly, the drug of BI6015 was used to specifically inhibit the expression of HNF4A, and select the appropriate concentration according to the instructions. The results showed that the expression of HNF4A was significantly inhibited when we gave 2, 4, and 8 μM drug concentrations, and at the same time, APOB showed the same expression trend. The results showed that APOB was regulated by HNF4A and positively correlated with the expression of HNF4A. Combined with the above research, we conclude that the high expression of APOB in IM tissues may also related to the increase of HNF4A expression.

Expression level analysis of HNF4A: (a) Box plot of HNF4A expression in different malignant tumors and normal tissues, abscissa is tumor type, and coordinate is gene expression level. Red represents tumor tissues and blue represents normal tissues (*P < .05, **P < .01, ***P < .001). (b) Box plot of the gradient of HNF4A expression levels in various malignant tumors, abscissa is tumor type, and coordinate is gene expression level. (c-e) Comparison of HNF4A expression levels in GES-1, HGC27, and MGC803 cells by western blot (left) and quantified(right) and All samples derive from the same experiment and that gels/blots were processed in parallel (*P < .05, **P < .01, ***P < .001). (f-h) WB was used to verify the changes in the expression of HNF4A and the corresponding changes in the expression of APOB under the action of different concentrations of specific inhibitor BI6015. The selection of different action concentrations of BI6015 is based on the suggestions in the instructions. All experiments were repeated three times, and P < .05 was considered statistically significant. (*P < .05, **P < .01, ***P < .001).
HNF4A-related pathway identification
After the key gene HNF4A was identified in gastric cancer, the potential mechanism of its biological function was investigated by enrichment analysis based on the gene set enrichment analysis (GSEA). Through bioinformatics analysis, we discovered that high expression of HNF4A in gastric cancer was significantly enriched in various pathways, including the cancer pathway, calcium signaling pathway, cytokine interactions, neuroactive ligand-receptor interactions, and olfactory signaling pathway. These results show that HNF4A may play an important role in cancer development (Figure 3).

GSEA enrichment analysis of HNF4A gene in gastric cancer.
Clinical correlation analysis of HNF4A and gastric cancer
In order to investigate the impact of HNF4A on the gastric cancer, we used online database resources to evaluate the activity of HNF4A in various cancers. Statistical analysis revealed that HNF4A exhibited significantly high activity in gastric cancer tissues (Figure 4a and b). Next, the correlation between the expression of HNF4A and the clinical features of patients with gastric cancer was investigated through the online database of UALCAN. The results revealed a significant association between HNF4A expression and gender, age, stage, as well as TP53 mutation status in gastric cancer patients. (Figure 4c-f). Furthermore, overall survival analysis curves drawn by the Kaplan-Meier plotter showed that patients with gastric cancer exhibiting high HNF4A expression had a poorer prognosis (Figure 4g). We also constructed survival curves of first progression survival time (FP) and postprogression survival (PPS), which further confirmed that elevated HNF4A expression is unfavorable for patients with gastric cancer (Figure S1).

Clinical correlation analysis of HNF4A: (a) Box plot of HNF4A activity in different malignant tumors and normal tissues (*P < .05, **P < .01, ***P < .001). (b) Box plot of the gradient activity of HNF4A in various tumors. (c) Association of HNF4A expression with age in gastric cancer; 41-60 years, P = 3.3 × 10−3; 61-80 years, P = 2.6 × 10−6; 81-100 years, P = 2.7 × 10−3. (d) Association of HNF4A expression with gender in gastric cancer; male, P = 2.6 × 10−6; female, P = 2.5 × 10−3. (e) Association between HNF4A expression and stage in gastric cancer; Stage 1, P = 1.4 × 10−3; Stage 2, P = 8.2 × 10−4; Stage 3, P = 2.4 × 10−5; Stage 4, P = 7.0 × 10−3. (f) Association of HNF4A expression with TP53 mutation in gastric cancer, P < .01. (g) Overall survival analysis of gastric cancer (n = 875). Kaplan-Meier survival curves: gastric cancer with low (n = 393) and high (n = 482) HNF4A expression (Log-rank P = 9.6 × 10−7). Affymetrix ID is 208429_x_at.
HNF4A expression and tumor infiltrating immune cells
Numerous studies have indicated a strong association between the tumor microenvironment and the initiation and progression of tumors. However, it remains unclear whether the microenvironment can facilitate the transition from intestinal metaplasia (IM) to gastric cancer. Therefore, we want to explore the correlation between HNF4A and immune-related infiltrating cells, so as to preliminarily explore the possible role of highly expressed HNF4A in IM. Tumor infiltrating immune cells are crucial components of tumor microenvironment (TME), and are considered as indicators of malignant tumors. Because of the complexity of various types of infiltrating cells in the tumor microenvironment, they can play different functions in promoting/suppressing tumor development. 28 Numerous studies have found that these infiltrating cells were involved in the induction of immune tolerance in gastric cancer. Furthermore, studies found that HNF4A can mediate immune signaling molecules and immune escape.29 -31 Therefore, Timer 2.0 and TISIDB databases were used to analyze the correlation between the expression of HNF 4A and tumor infiltrating immune cells. Our analysis revealed a significant correlation between HNF4A expression and various tumor-infiltrating immune cells. Notably, HNF4A expression exhibited positive correlations with CD8+ T cells, common myeloid progenitor cells, granulocyte-monocyte progenitor cells, hematopoietic stem cells, and myeloid dendritic cells. Conversely, several tumor-associated infiltrating immune cells were negatively correlated with HNF4N expression (Figure 5a). Similar results were also observed in the TISIDB database (Figure 5b). Although these results suggest that HNF4A may promote cancer development. Its positive correlation with T cell follicular helper, neutrophil, as well as its negative correlation with cancer-associated fibroblasts, suggest a more complex underlying regulatory mechanism. Subsequently, the correlation between the expression of HNF4A and immune-related factors was analyzed. These findings suggest HNF4A may exert regulatory influence on immune responses within the tumor microenvironment, potentially impacting tumor progression and immune surveillance. However, further investigations are required to fully elucidate the underlying mechanisms and functional implications of these associations (Figure 5c and d).

Correlation analysis between HNF4A expression and tumor-associated infiltrating immune cells: (a-b) Overall correlation analysis between HNF4A expression and tumor-associated infiltrating immune cells, red represents positive correlation and blue represents negative correlation. (c) Correlation analysis between HNF4A expression and immune-activating factors. (d) Correlation analysis of HNF4A expression with immunosuppressive factors.
Alteration analysis of HNF4A in gastric cancer
Previous studies have established a strong association between HNF4A and APOB, as well as tumor infiltrating immune cells, and the high expression of HNF4A is closely related to the prognosis of gastric cancer patients. Therefore, our objective was to investigate the genetic alteration of HNF4A in gastric cancer and explore its possible role. Firstly, the cBioPortal website was used to analyze the mutation frequency of HNF4A in different malignant tumor samples. Notably, the results indicated that the mutation frequency of HNF4A in gastric cancer samples was comparatively high, accounting for approximately 0.02% (Figure 6a). Subsequently, we analyzed the mutation situation of HNF4A in gastric cancer in more detail, focusing on the specific types of alternation and the differences among different subtypes of gastric cancer. The findings revealed that while the number of HNF4A alterations in signet ring cell carcinoma of the stomach samples was relatively low, there was a notable increase in both the copy number and mutation rate. This observation strongly aligns with our current understanding that signet ring cell carcinoma of the stomach is the most aggressive subtype among all gastric cancer subtypes (Figure 6b and c). In addition, we investigated the high-frequency mutation sites, types, and their corresponding protein domains of HNF4A in gastric cancer across different conditions. The results that demonstrated the missense mutation of HNF4A were predominantly observed under normal conditions and following phosphorylation modification. Truncated and in-frame mutation was more prominent after acetylation and ubiquitination modification (Figure 6d). Finally, we conducted a preliminary investigation to determine whether HNF4A has the potential for immunotherapy by examining the correlation between tumor mutational burden (TMB), microsatellite instability (MSI), and HNF4A. Currently, TMB and MSI have been recognized as important biomarkers associated with the sensitivity of immune-checkpoint inhibitors (ICIs). Studying the correlation between these biomarkers and tumors is crucial for predicting the effectiveness of immunotherapy. Our results showed that TMB and MSI had a strong positive correlation with HNF4A in gastric cancer (Figure 6e and f). The above conclusion indicates that high expression of HNF4A is frequently correlated with a poor prognosis among patients with gastric cancer. Meanwhile, HNF4A also showed the potential for immunotherapy.
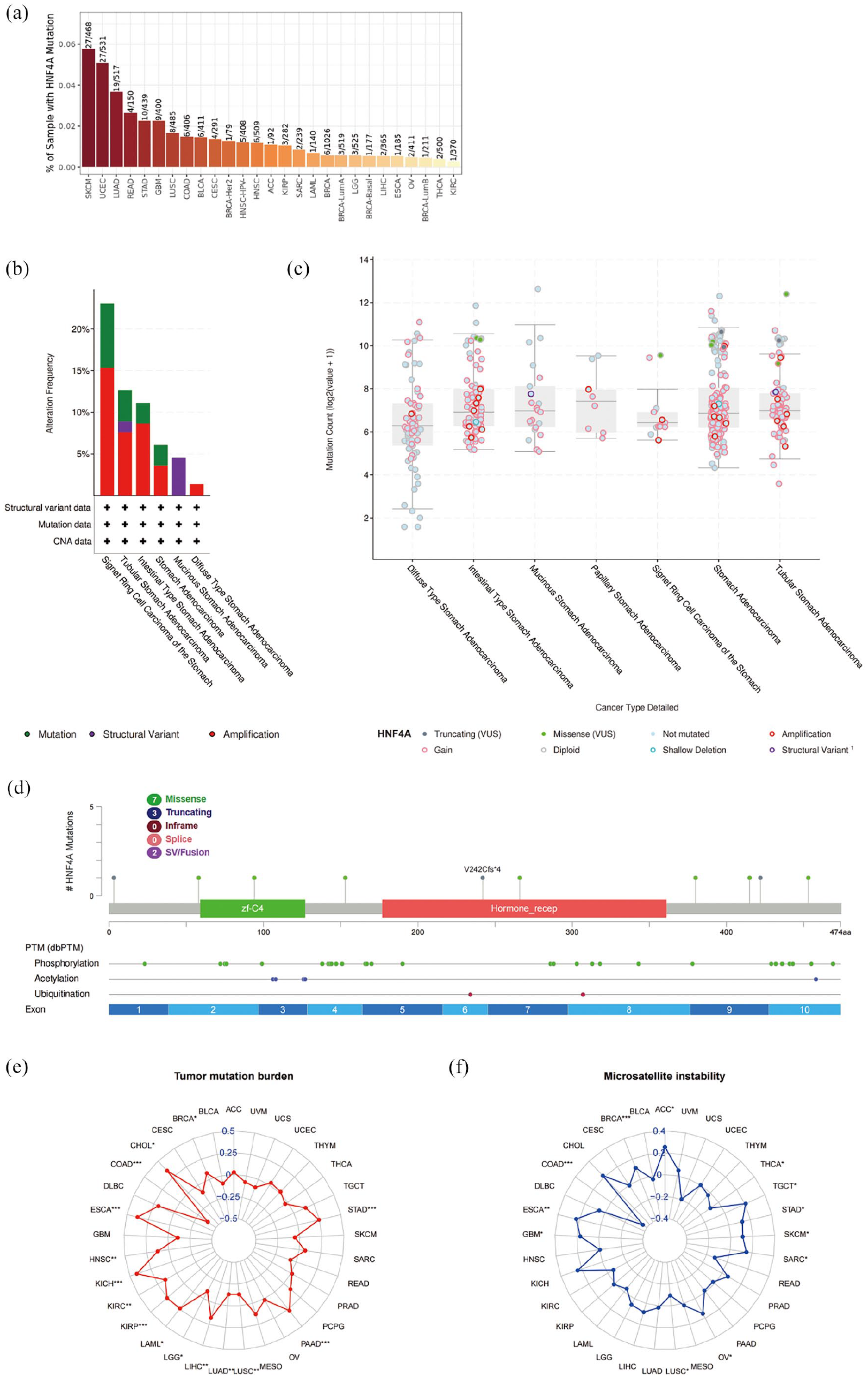
HNF4A alteration analysis in gastric cancer: (a) The overall mutation frequency of HNF4A in malignant tumors. The horizontal axis represents the type of malignant tumor, and the vertical axis represents the mutation rate of the gene. (b) The alteration frequency with mutation type of HNF4A in different subtypes of gastric cancer. The horizontal axis is the histological subtype of gastric cancer, and the vertical axis is the alteration frequency. The color of the bar chart represents the corresponding change type (green represents mutation, red represents structural variant, and purple represents amplification). (c) Mutation counts of HNF4A in different subtypes of gastric cancer. (d) Mutation sites, types, and corresponding protein domains of HNF4A in gastric cancer under different conditions. (e-f) Radar plots of HNF4A expression correlation with TMB and MSI, and the numbers in the radar map represent the correlation between the target gene and the corresponding cancer TMB and MSI (*P < .05, **P < .01, ***P < .001).
Discussion
In this study, differentially expressed genes between IM tissues and normal gastric tissues were identified from the GSE 78523 dataset. Though enrichment and PPI analysis of these genes, we observed a close association between APOB and lipid metabolism in IM. In order to explore the regulation mechanism of APOB expression changes in IM, the upstream regulator HNF4A was identified by hTFtarget. Finally, the correlation between the expression of APOB and HNF4A was verified by correlation analysis and western blot.
The gene of APOB encodes two proteins, Apob100 and Apob48. Apob100 is the primary constituent of low-density lipoprotein (LDL) and is exclusively synthesized in the liver. 32 Apob48 participates in the assembly of chylomicron (CM) in the intestine, a process catalyzed by apolipoprotein B mRNA editing enzyme, catalytic polypeptide 1 (APOBEC1).33,34 Macrophages have two primary pathways for lipid uptake. The first is scavenger receptor-mediated lipid uptake, which is a well-known pathway. The second pathway involves LDL receptor-mediated lipid uptake. Furthermore, apob48r receptor expressed in macrophages can specifically binds to apob48, which is considered to be another specialized pathway for lipid metabolic uptake. Previous research conducted over the past few decades has provided evidence demonstrating that this pathway operates independently of APOE.35 -37 This process is critically involved in the formation of foam cells and the development of related diseases, including atherosclerosis and xanthomas.38,39 A clinical cross-sectional study showed that there was a more significant correlation between IM and gastric xanthomas, which was considered as an independent factor to predict IM. 40 Meanwhile, numerous clinical studies have shown that gastric xanthomas were closely related to the development and rapid growth of gastric cancer. 10 It was also regarded as a reliable marker to predict early gastric cancer. 41 Based on preliminary data analysis, we speculated that the high expression of APOB in IM might contribute to the increase of lipid uptake of infiltrated macrophages, subsequently inducing the development of gastric cancer by promoting the formation of a lipid microenvironment.
In our results, microsomal triglyceride transfer protein (MTTP), aldolase B (ALDOB), fatty acid binding protein 1 (FABP1) also plays important role in promoting APOB to play its own functions. MTTP, which is highly expressed in adipose tissue, plays a crucial role in regulating lipid metabolism by facilitating the transport of triglycerides between membrane vesicles. 42 It is co-located with APOB in a tissue-specific way, and its existence ensures the stability of apob100. 43 ALDOB primarily participates in fructose metabolism. While limited studies have explored the association between ALDOB and APOB, a substantial body of research has indicated that elevated ALDOB expression is often associated with a poorer prognosis in cancer patients. 44 The main function of FABP1 is to uptake fatty acids, which is essential for maintaining the dynamic balance of cell membrane, signal transduction, and proliferation of tumor cells. However, besides FABP1, there are also other fatty acid uptake pathways. 45 Besides, what role it plays in gastric cancer needs further investigations.
In subsequent research, we found that HNF4A could potentially promote gastric cancer by interacting with tumor-associated immune infiltrating cells. Previous studies have also demonstrated that HNF4A plays a crucial role in regulating energy metabolism, which is essential for the proliferation and metabolism of gastric cancer cells.20,46,47 Therefore, we speculate that the high expression of HNF4A in IM may serve as a trigger for gastric cancer. Our experiments have revealed that HNF4A can regulate APOB, and there is a positive correlation between them. IM is considered a precancerous lesion, but whether it is a direct precursor for gastric cancer is still controversial. 48 Recent theories have shown that have revealed that gastric cancer is caused by increased genetic instability of gastric stem cells rather than a direct transition from IM to cancer. 49 This conclusion can explain why the expression level of lipid metabolism-related genes, such as APOB, were dramatically altered in gastric cancer compared with IM.
Based on our hypothesis, the interaction between HNF4A and APOB has the potential to impact the microenvironment, leading to increased metabolic stress in normal cells and ultimately facilitating the development of gastric cancer. Meanwhile, the lipid microenvironment could also promote the expression of HNF4A in gastric cancer cells and establish a positive feedback loop. However, further experiments are needed. The identification of tumor-associated macrophages (TAM) is closely related to cancers. 50 Previous clinical studies have revealed that gastric macula with foam cells as pathological features was closely related to gastric cancer. Nonetheless, limited research has been conducted on the underlying molecular mechanisms. Through bioinformatics analysis, we hypothesized that the gastric microenvironment can be changed by the high expression level of APOB. This alteration could influence the lipid uptake ability of macrophages, which may be related to the procession of inflammation, repair, and metabolism of surrounding normal gastric epithelial cells. This complex effect may promote the development of gastric macula, which in turn contribute to the development of gastric cancer. Further studies are necessary to identify other important factors that may be involved.
There are some limitations to the present research. First, our study relied on a small sample size and only conducted preliminary validation through western blot analysis. Second, further investigations including correlation analysis, as well as cellular and animal experiments are needed. In the subsequent experiments, we aim to increase the sample size to minimize the errors caused by individual differences. Additionally, we plan to conduct a more comprehensive analysis of lipid metabolism differences among IM tissues, normal tissues, and gastric cancer tissues, while also exploring the key intermediate metabolites involved. These proposed measures will help to address the limitations and provide a more robust understanding of the topic.
Conclusion
Through our experiments and analyses, we have made initial discoveries regarding a novel mechanism that underlies the transition from IM to gastric cancer. One crucial aspect of this process is the significant role played by HNF4A. Our findings provide compelling evidence supporting the idea that the occurrence and progression of gastric cancer are influenced by the interplay between HNF4A expression and lipid metabolism. These results shed light on the complex relationship between HNF4A and gastric cancer and contribute to our understanding of the disease at a molecular level.
Supplemental Material
sj-tif-1-evb-10.1177_11769343241249017 – Supplemental material for HNF4A-Bridging the Gap Between Intestinal Metaplasia and Gastric Cancer
Supplemental material, sj-tif-1-evb-10.1177_11769343241249017 for HNF4A-Bridging the Gap Between Intestinal Metaplasia and Gastric Cancer by Yihang Zhao, Hong Tang, Jianhua Xu, Feifei Sun, Yuanyuan Zhao and Yang Li in Evolutionary Bioinformatics
Footnotes
Acknowledgements
We thank Dr. Xu and Dr. Zhang from the Department of Gastroenterology of the First Affiliated Hospital of Anhui Medical University for offering their help. Dr. Xu provided help in verifying the interaction between SIRT3 and HIF1A, and Dr. Zhang provided suggestions for the whole research design idea.
Authors’ Contributions
Yang Li completed the conception and design and provided administrative support. Feifei Sun and Jianhua Xu were responsible for the collection of clinical samples. Feifei Sun and Yuanyuan Zhao were responsible for the collection and assembly of data, and Yihang Zhao was responsible for the analysis and interpretation of data. All the authors participated in the writing of the manuscript and approved the final version of the manuscript.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by grants from the National Natural Science Foundation of China (No. 61976007), Youth Program of Natural Science Foundation of Anhui Province (No. 2008085QH415), and Youth Fund of National Natural Science Foundation Projects (No. 82103040).
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Availability of Data and Materials
Ethics Approval and Consent to Participate
We declare all experiments in this study were performed in accordance with the Declaration of Helsinki. The study was approved by the ethics committee of Anhui Medical University (No.20190199), and informed consent was obtained from all patients.
We declare all experiments in this study were performed in accordance with the Declaration of Helsinki (as revised in 2013). The study was approved by the ethics committee of Anhui Medical University (No.20190199), and informed consent was obtained from all patients.
Consent to Publish
Not applicable.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.