
Abstract
An emerging alternative to conventional animal models in toxicology research is the zebrafish. Their accelerated development, regenerative capacity, transparent physical appearance, ability to be genetically manipulated, and ease of housing and care make them feasible and efficient experimental models. Nonetheless, their most esteemed asset is their 70% (+) genetic similarity with the human genome, which allows the model to be used in a variety of clinically relevant studies. With these attributes, we propose the zebrafish is an excellent model for analyzing cognitive and neuromuscular responses when exposed to toxicants. Neurocognition can be readily analyzed using visual discrimination, memory and learning, and social behavior testing. Neuromuscular function can be analyzed using techniques such as the startle response, assessment of activity level, and evaluation of critical swimming speed. Furthermore, selectively mutated zebrafish is another novel application of this species in behavioral and pharmacological studies, which can be exploited in toxicological studies. There is a critical need in biomedical research to discover ethical and cost-effective methods to develop new products, including drugs. Through mutagenesis, zebrafish models have become key in meeting this need by advancing the field in numerous areas of biomedical research.
Introduction
It is well established that zebrafish have numerous qualities that make them excellent models for scientific research.1-3 The goal of this review is to establish the feasibility of the zebrafish as a convenient and functionally appropriate model organism for use in behavioral studies. Examples will be given demonstrating their robust cognitive capabilities, methods to test their cognition, and observable stereotyped motor behaviors which can be manipulated upon application of toxicants to their environments. To add validity, functional similarities between the zebrafish and humans will be discussed to portray how zebrafish as mutant models are evolving and becoming key to toxicology investigations. To conclude, the ethical benefits of zebrafish will be addressed.
Materials and Methods
Articles referenced were accessed through PubMed’s database of biomedical literature, the Zebrafish International Resource Center, the Humane Society International, and Stanford Medicine. Keywords searched included zebrafish, zebrafish neuromuscular function, zebrafish motor function, zebrafish toxicology, zebrafish mutants, animal models toxicology, object recognition memory zebrafish, recognition memory zebrafish, zebrafish social behavior, zebrafish social behavior review, shoaling zebrafish. Exclusion criteria prioritized the most recent experimental studies except for a few seminal publications. Figures were developed using Microsoft Office and Adobe PDF software systems.
Results and Discussion
Convenience and Feasibility
Zebrafish practicality is evidenced by their low maintenance housing and care,4-8 accelerated life span,9-11 capability to regenerate,12-17 transparent physical appearance,18,19 and their ability to be genetically manipulated.20-25 The natural habitat of zebrafish are freshwater creeks and lakes in tropical regions of India and Nepal. 26 Although native to South Asia, zebrafish can survive in habitats of varying pH, temperature, depth, and speed of waterways.4-7 Their adaptability makes creating an optimal experimental environment simple without disrupting their living habits.4-8 For example, stocking density can vary between 16-200 embryos per petri dish in the early larval stage without adversely affecting survival rates. 5 Additionally, experimentation itself can be made quite simple; even tests analyzing higher order function can be completed using basic lab equipment.27-32
During the adult stage, creating a proper environment is simple, requiring minimal maintenance and basic equipment.33,34 The process is made even easier to manage with guidelines that are readily available for investigators to follow. These guidelines include information on supply and housing conditions as well as how to perform experiments with specified chemical variables, water quality levels, pH, and temperature.4,5,7 Keeping a favorable supply of zebrafish without breeding is also quite feasible with their cost being considerably lower than rodents 8 and primates. 35
Zebrafish development is rapid in comparison to humans. As young as 3 days post-fertilization (dpf), the embryonic development stage is completed in addition to a large degree of their biological shaping and structure. 9 Metamorphic changes rapidly progress from the juvenile and adult stages and are completed by 22 dpf, finalizing their social and physical maturation.9-11 Concerning appearance, during the first two weeks of life, zebrafish are translucent, which allows an investigator to easily measure alterations in morphological changes in the developing brain, muscle,18,19 and other anatomical structures.36-41 Zebrafish can also be readily manipulated genetically to serve as mutant models.20-25 This is of particular interest since over 70% of their genome is homologous to the human genome. 42 Because of these reasons, the zebrafish model is becoming a relevant and valuable alternative in biomedical research, especially toxicology.33,34,36-39,43
Zebrafish as a Model of Cognitive Function
Cognitive Function.
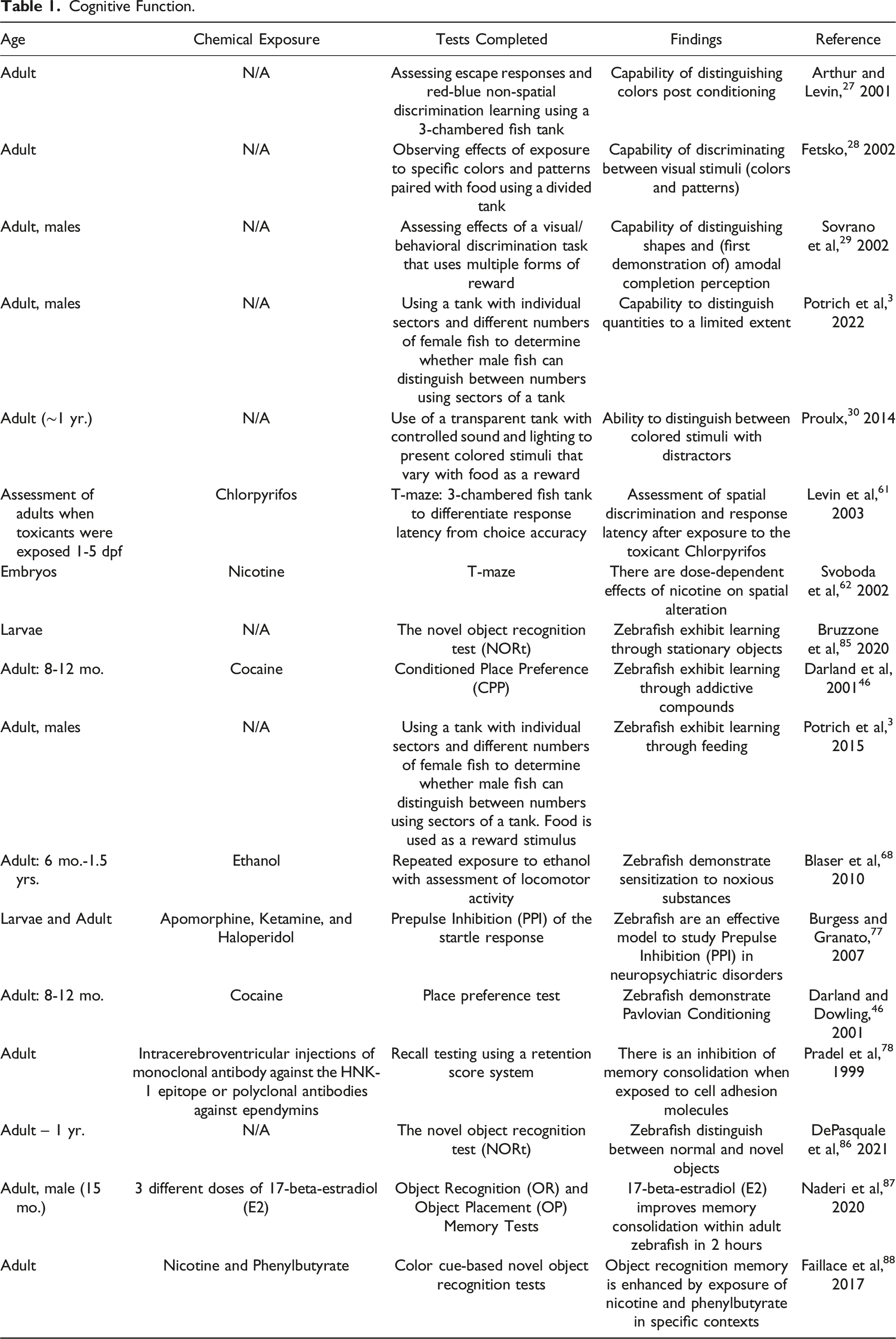
Visual Discrimination
At the dawn of the 21st century, two key findings were published regarding zebrafish’s visual discrimination abilities. In 2001, Arthur and Levin 27 determined that zebrafish can distinguish colors after being conditioned. A year later, it was demonstrated zebrafish possess the ability to discriminate between visual stimuli in more advanced capacities. 28 Using a divided tank, zebrafish distinguished between different colors (e.g., blue and yellow, blue and orange, blue and green, yellow and orange) and different patterns (e.g., vertical lines and horizontal lines) when the stimuli were paired with food. 28 These two experiments formed the foundations for further exploration. Interestingly, zebrafish may be naturally attracted to specific colors because of their short cones’ responses to their surroundings. 57 Siregar et al 58 established evidence-based standardized methods for testing natural color preference, allowing visual discrimination experiments to be implemented more effectively. Subsequent experiments showed that zebrafish are capable of distinguishing between shapes. 29 Potrich et al later discovered that zebrafish exhibit amodal completion using simple shapes. 29 With this ability, they can identify certain objects as being whole, even when the objects are partially blocked from their line of sight. Furthermore, zebrafish are able to distinguish quantities to a limited extent. 3
Another cognitive trait of zebrafish is their ability to implement visual parallel search mechanisms; for example, they can distinguish between different colored stimuli amidst other distractors. 30 In fact, when the distractions increased up to 10 times, the accuracy of the response was not diminished. 30 Since zebrafish can discriminate between objects and colors simultaneously, this also means they can distinguish between objects or colors independently. For example, they would be able to recognize the difference between two cubes of the same color while also recognizing the difference between a cube and a sphere of the same color. These findings reveal yet another layer of complexity to the zebrafish’s cognitive abilities.
Zebrafish’s visual abilities, like humans, can be significantly affected by environmental toxicants. 59 For example, N-methyl-N-nitrosourea (MNU) causes retinal degeneration in zebrafish. 59 In humans, cannabis can hinder contrast sensitivity and motion discrimination. 60 Studying the effects of neurotoxicants on zebrafish could help us understand this relatively unexplored condition.
Memory and Learning
Visual Discrimination and Learning
Zebrafish are now a choice model to test memory and learning along with visual discrimination. 51 Memory and learning are often analyzed alongside visual discrimination using a T-maze apparatus. Colwill et al 45 provided the basis for this model using three visual discrimination experiments. In their first two experiments, they established that color selection could be conditioned and then extinguished. The same result was demonstrated in a third experiment with pattern selection. In this context, zebrafish were rewarded with food for choosing the correct arm of the maze. The fish soon learned to choose the correct arm, but this skill was quickly extinguished when the stimulus was repeatedly presented without the food reward. The same T-maze design was used by Levin et al 61 to test the effect of chlorpyrifos on acquiring spatial discrimination learning in zebrafish. In a subsequent experiment, Levin et al determined that nicotine significantly improved zebrafish’s learning of delayed spatial alteration in low doses but diminished it in higher doses. 62
Zebrafish can also demonstrate learning without using colors as cues. Since Colwill et al foundational experiments, researchers have used cues ranging from stationary objects63,64 to addictive compounds 46 to feeding. 3 In each context, the zebrafish forms a cognitive link between a stimulus and a sensation. In this context, learning takes place by association, and a memory is formed.
Non-Associative Learning
Multiple studies have shown that the strength of an animal’s response to a stimulus can decrease over time without sensory adaptation, motor fatigue, or injury.65-67 In other words, learning can occur without true memory, which has been demonstrated in zebrafish.
55
The current theory used to explain this phenomenon is the dual-process theory, which describes two connected forms of non-associative learning: sensitization and habituation (followed by dishabituation).
52
Figure 1 displays graphical examples of the effect of habituation and sensitization in zebrafish. Sensitization occurs when an organism is exposed to an arousing stimulus that causes a response
65
; this stimulus is often noxious. Zebrafish have been shown to demonstrate sensitization to many substances, including nicotine
55
and alcohol.
68
Klee et al
55
suggested that zebrafish are an effective preclinical model for understanding the mechanisms behind the effects of nicotine exposure in humans, in large part because of their high reproduction rate and transparent embryos. These aspects allow zebrafish to be studied in large numbers and with unique precision; researchers can directly observe the impact of a developmental neurotoxin on the maturation stages of nervous system development.62,69 Graphical representation of habituation and sensitization curves in zebrafish. (A) Habituation—as rapid, repeated stimuli increase, responsiveness decreases over time and (B) sensitization—when stimuli are presented with larger time intervals, responsiveness increases to the point of complete sensitization.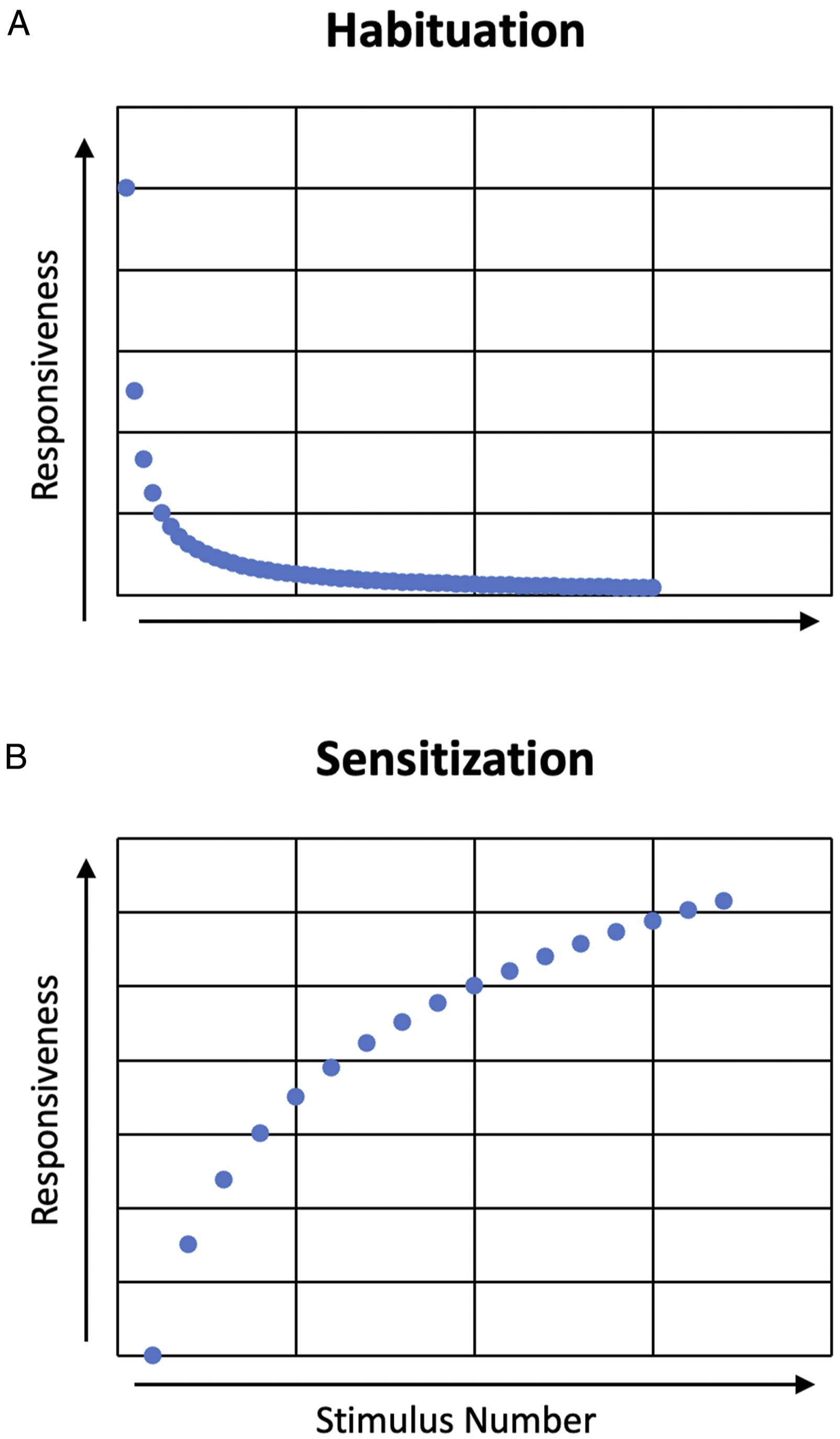
In most cases, the same substance employed to sensitize an organism can be used to elicit a habituation response. 65 As shown in Figure 1(B), habituation describes the process of repeated exposure to a stimulus, which leads to a decreased response. 52 This has direct relevance to human toxicology studies: nicotine, for example, is known to generate habituation/addiction in humans. 70
Although poorly understood mechanistically, habituation is a prerequisite for more complex types of learning.
66
One example of habituation in zebrafish is that they respond less to startling stimuli over time.
65
Zebrafish exhibit a stereotyped C-start response to startling sensory stimuli
65
; their bodies form a C-shaped curve, as depicted in Figure 2(A). Similarly, zebrafish have been shown to perform an O-bend at the extinction of light.
65
This is essentially the same reaction with a much larger amplitude, as seen in Figure 2(B). As Roberts et al
65
explain, three types of startle response habituation exist in zebrafish larvae. Rapid habituation develops after 50-120 rapid auditory pulses but lasts only 3 to 15 minutes.
71
Short-term habituation is created by increased time intervals in auditory training (about 5 minutes in length), and its effects last up to 1 hour.
71
Long-term habituation requires six-spaced blocks of auditory training, but its effects can last up to 18 hours.
72
Dishabituation occurs when the fish’s reduced response is recovered by presenting a novel stimulus.
66
A brief pulse of light
73
and a tactile stimulus with a handheld poker to the head
74
have been used to generate this effect. Most research focuses on the habituation of zebrafish larvae, but adult zebrafish have also been reported to exhibit the same pattern.
75
Essentially, all organisms, including humans, use non-associative learning to adapt their behavioral response to repeated recent stimuli.
76
Because of this, studies such as Burgess and Granato’s
77
experiment make zebrafish an appealing model to study neuropsychiatric disorders in humans. Their experimentation revealed both larvae and adult zebrafish use sensorimotor gating by looking at prepulse inhibition (PPI) of the startle response. Thus, zebrafish are an effective model to better study PPI, which is present in numerous human neuropsychiatric disorders. Startle response behavior in zebrafish. (A) C-start response is characteristic of a C-bend of the body and (B) O-bend response; amplification of the C-start response.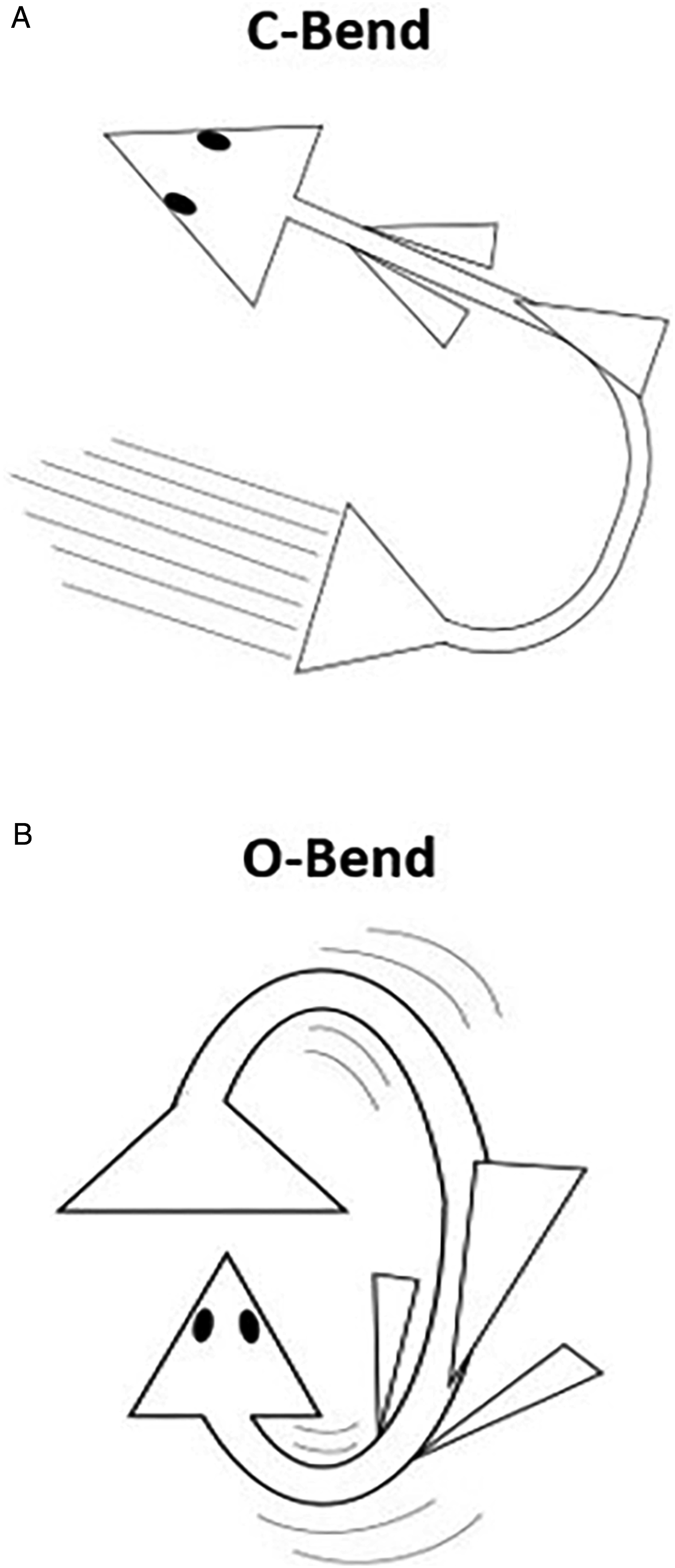
Basic Associative Learning—Pavlovian Conditioning
Darling and Dowlin first demonstrated Pavlovian Conditioning in zebrafish in its most basic appetitive form using a “place preference” test consisting of a 2-liter tank divided by a perforated wall. 46 When given cocaine, the zebrafish tended to gravitate toward the cocaine-associated side of the container. Interestingly, cocaine also increased their visual threshold, making them less sensitive to light. To explain this phenomenon, Darling and Dowlin proposed a direct effect on the zebrafish brain; an electroretinogram confirmed that these changes were physiological (the brain itself was affected) and not behavioral. 46 Cocaine sensitivity also has a genetic component; in their experiments, two fish with low cocaine sensitivity produced an F2 generation with significantly lower cocaine sensitivity than controls. 46 Similarly, Williams et al trained fish to alternate feeding sites in a simple rectangular tank. 3 There is also evidence of avoidance learning in zebrafish. This is essentially the negative form of Pavlovian Conditioning, in which zebrafish learn to avoid a stimulus vs gravitating toward it. Research has revealed that avoidance learning is influenced by group size, 47 cell adhesion molecules,78,79 and NMDA. 80
Pavlovian fear responses have also been demonstrated in zebrafish.80,81 This response has been linked to second-order conditioning, a phenomenon that has yet to be extensively researched in humans.82,83 Many different aspects of Pavlovian conditioning have been characterized in zebrafish, making them a helpful model to investigate similar behaviors in humans.
Object Recognition Memory
Extensive research has been conducted regarding the zebrafish’s ability to distinguish between novel and familiar objects in addition to their preference for one or the other. This ability is most often tested using the novel object recognition test (NORt). 64 Using this test, researchers have explored object recognition memory in zebrafish. Contrary to results in rodents, however, zebrafish spend more time exploring familiar objects vs novel objects. 84 It has been predicted that this is the result of “neophobia” caused by natural anti-predatory behavior. Such neophobia has been speculated to be the case in larvae as well. Bruzzone et al 85 discovered that zebrafish explore novel objects more often at 14 days post-fertilization (dpf) than at 21 dpf, when their anti-predatory instincts are more developed.
Multiple factors affect object recognition memory in zebrafish. For one, environmental enrichment (i.e., gravel substrate and plastic plants) positively influences object recognition memory in the NORt. 86 17-beta-Estradiol (E2), a substance known to modulate memory and learning in zebrafish, has been shown to increase object recognition memory in high doses by activating estrogen receptors. 87 E2 improves memory consolidation in male adult zebrafish within 2 hours. Naderi et al 87 showed that exposure to E2 also increases the number of synaptic protein-encoding genes, which suggests that E2 affects the specific proteins that encode neural pathways. In addition, nicotine and phenylbutyrate increase object recognition memory, although within different timeframes. 88
Social Behavior
Social Behavior.
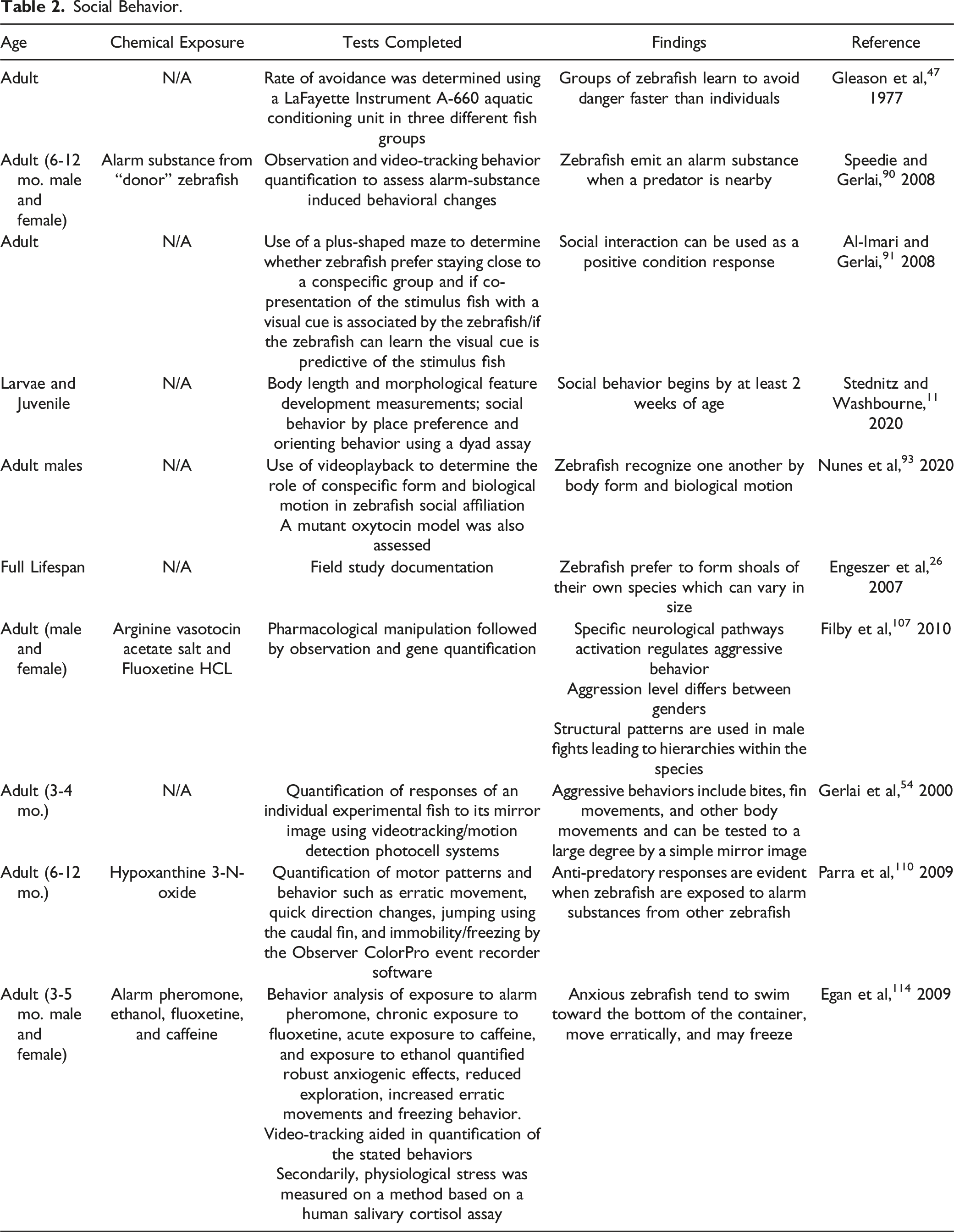
Conspecific Social Interaction
Social behavior in zebrafish begins at a very young age.11,53,56 Hinz et al 53 showed that visualizing conspecifics has “rewarding properties” in 6- to 8-day-old larval zebrafish, suggesting that it can act as an unconditioned stimulus when paired with light as a conditioned stimulus. Contrary to this finding, Dreosti et al 56 showed when using a compartmental behavioral assay, zebrafish show no social preference at one week. This suggests that social behavior begins to develop at two weeks and becomes more advanced by three weeks post-fertilization. Most recently, Stednitz and Washbourne 11 demonstrated that zebrafish develop a variety of complex social behaviors between 10 and 16 dpf. Although conflicting data exist on the precise time that social development begins, it is clear that zebrafish larvae prefer to swim near conspecifics by three weeks. This aligns with the rapid development of human social behavior which is exhibited within the first year of life.
The development of social behavior in humans and how it may be impacted by neurotoxicants can be explored using zebrafish. Nunes et al 93 provide a comprehensive description of how zebrafish recognize conspecifics visually. In summary, zebrafish recognize one another through two visual features: body form and biological motion. In their experiment, zebrafish exhibited social behavior when seeing an image of the conspecific alone, but the effect was more significant when biological motion was implemented. Biological motion also inherently generates social behavior. Similarly, humans also can distinguish conspecifics using mere form and motion; for example, Kozlowski and Cutting 94 demonstrated that humans can recognize the gender of someone walking using only a point-light display. Nunes et al 93 have suggested that a common underlying mechanism may exist in all vertebrates that deduce information from mere form and motion.
Shoaling Behavior (“Shoaling Zebrafish”)
Zebrafish prefer to form groups with phenotypically similar individuals of their own species; these groups are referred to as shoals.26,95-97 Shoaling behavior of wild zebrafish has been fairly well-documented and provides information on zebrafish interactions with one another.26,92,98,99 Shelton et al conducted a comprehensive study of zebrafish shoals at four sites in India, one of many South Asian regions where the fish naturally thrive. 26 In this study, it was discovered that zebrafish shoals vary significantly in size (from 2000 fish per group to only 11 fish per group) and average nearest-neighbor distances (NNDs). These differences largely depended on water flow, as faster-flowing water led to increased group size and decreased NND. Vegetation surprisingly had no effect, which contrasts with laboratory experiments performed on wild zebrafish, suggesting that vegetation increases aggression.98,99 Additionally, it has been shown that not all members of this species act in the same manner. It was shown in an experiment comparing six zebrafish strains, each strain displayed its own distinct level of aggressiveness, interest in conspecifics, and shoaling habits. 100 Because of this, a remarkable number of zebrafish strains have been employed as behavioral models,87,100,101 and additional strains and mutant models might be useful for measuring uninvestigated cognitive functions translatable to human neuroscience research.
Experimentation on shoaling behavior in zebrafish might seem laborious to quantify, as it involves keeping track of up to 2000 individual zebrafish in a single group. Fortunately, Miller and Gerlai 102 have developed a simple and efficacious computer software system to track zebrafish shoaling. In cases of extremely large shoals, smaller groups are sampled via video recording and used as a representative sample or scaled by calculations to estimate values for the group as a whole. This central cognitive function can be measured effectively in zebrafish.
Aggression
Zebrafish aggression is most often measured by observing dyadic fights involving two zebrafish competing for dominance. 103 These interactions include counting the number of chases and bites in each fight.104,105 Aggressive interactions between male zebrafish follow a structured pattern. 106 These battles begin with an extension of the fins, circling, and biting attempts. Battles gradually increase in intensity until both fish implement frequent biting attempts, chasing, and attempted strikes at the opposing fish. 106 The losing fish eventually becomes submissive and exhibits passive, subordinate behavior like freezing. The dominant-submissive hierarchies that arise from these fights can last five days or longer,105,106 and these dominant fish remain more aggressive than their submissive counterparts. 92 Generally, males are more aggressive than females.107,108 This model could be a helpful method to understand various vertebrates’ biological aggression mechanisms between sexes, and it could even shed light on human aggression.
Zebrafish aggression can be tested using mirror image simulation, 109 which involves placing a mirror at a 22.5° angle outside a zebrafish’s tank. 54 The zebrafish’s aggressive reaction (bites, fin movements, etc.) is measured in response to its mirror image. This test is simple and does not harm the fish, but it does not perfectly capture aggression; a zebrafish’s reaction to its mirror image is a poor representation of a wild interaction between zebrafish. 103 Therefore, other methods have been explored which have more than compensated for these shortcomings. Most notably, investigations using a direct dyadic interaction between two zebrafish are highly effective in providing a realistic model of aggression. 106 Additionally, Gutierrez et al have established an automatic mirror image aggression model using one-month-old zebrafish, allowing higher throughput experiments. 26 The primary use of this model focuses on juvenile fish because of their small size. Using larval models, researchers could explore how potentially toxic chemicals mitigate or even increase aggressive behavior. Although models are still being developed, zebrafish aggression has been well-documented and has tremendous potential to enlighten the scientific community about human aggression.
Predator Avoidance
Zebrafish exhibit anti-predatory responses when exposed to alarm substances from other zebrafish, such as synthetic hypoxanthine 3-N-oxide. 110 Interestingly, researchers have demonstrated that humans may respond similarly to certain substances, which is a phenomenon called drug-induced panic attacks. 111 After this discovery, researchers were able to produce anxiety responses on command in zebrafish, which has allowed their predator avoidance capabilities to be studied in greater depth.
The nature of anxiety responses in zebrafish has been extensively studied.112,113 Anxious zebrafish tend to swim toward the bottom half of a container, move erratically throughout the container, and exhibit freezing behavior. 114 Speedie and Gerlai 90 have hypothesized that the intention behind this behavior is to generate turbulence and stir up debris, thus clouding the zebrafish from nearby hunters. This hypothesis explains all three of zebrafish’s most stereotypical anxiolytic behaviors. It is a reasonable hypothesis that, in the wild, zebrafish might swim toward the bottom of a creek to locate themselves near sediment, generate a cloud of debris by swimming erratically, and then quickly freeze to prevent detection from nearby predators.
It has been estimated that human anxiety disorder has a worldwide prevalence of 12.9%, 115 and up to 33.7% of the worldwide population are burdened by some type of anxiety disorder during their lifetime. 116 In addition, only about 60%–85% of individuals experience positive effects from anti-anxiolytic medicine, and only 50% of these responders reach complete recovery. 117 Anxiety disorder is on the rise in the United States, although it is debated whether this increase is caused by a relaxation in the Diagnostic and Statistical Manual of Mental Disorders criteria. Nevertheless, human anxiety has extensive and detrimental effects on human life. 118 Any research findings that could alter this mental disorder's prognosis could literally change the world.
Zebrafish as a Model of Neuromuscular Structure and Function
Structural Organization
As previously shown with respect to cognitive function, zebrafish have also shown great promise in elucidating the mechanisms involved in behavioral changes and neuromuscular function. The zebrafish is structurally homologous to humans with respect to genetics, protein structure, muscle anatomy, and neuroanatomy.8,36,42,119-121 About 80% of a fish’s body mass is skeletal muscle, 122 and in the case of zebrafish, the skeletal muscle structure is easily observed because of its transparent nature.18,19,122,123 Commonalities between the motor units of zebrafish and humans are easily identifiable and include the structure of spinal motor neurons, skeletal muscle fibers, the brainstem, and the motor cortex. 124 Babin et al 124 showed evidence of similarity between motor units using in vivo applications. A key similarity between the spinal motor neurons of humans and zebrafish is their shared alpha-motor neurons and various subtypes of motor neurons. Furthermore, zebrafish and humans share aspects of skeletal muscle fiber structure, such as the preservation of the dystrophin-associated glycoprotein complex, excitation-contraction coupling machinery, and the contractile apparatus; dysfunction of these structures results in human muscle diseases. 125 The similarities in protein structure, muscle anatomy, and neuroanatomy between humans and zebrafish allow for a wide range of investigations for studying the effects, causes, and potential treatments of human muscular diseases.
Assessment of Motor Function
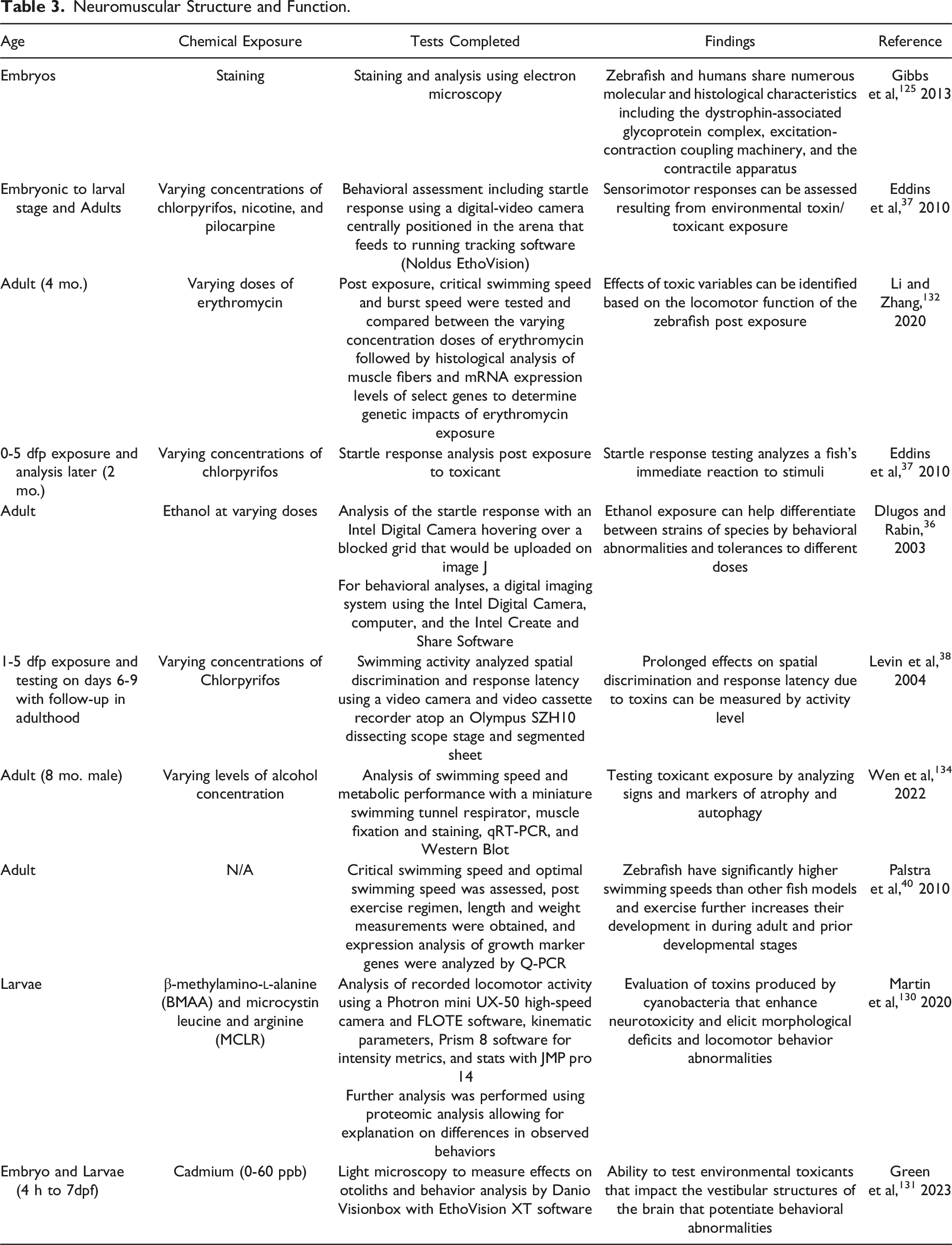
Structure determines function.126,127 Therefore, it is reasonable to conclude that with the vast number of structural similarities between zebrafish and humans, there would be numerous functional similarities as well. 128 To test functionality, there are several neuromuscular tests that hold promise for investigating toxic variables using zebrafish. As outlined in Tierney’s three tier Behavioral Hierarchy Model, 128 the lowest tier and most fundamental is motor function analysis through measurement of swimming patterns, swimming speeds, and/or distance covered. The second tier is sensorimotor responses, which can be analyzed through assessments of food attraction, odor attraction, and alarm response (i.e., startle response).36,37,128 The highest behavioral assessment level, placed at the top tier, is a cognitive response; this is where the cognitive endpoints are determined by investigating learning and memory. These behavioral responses can be predicted through the testing of a subject's locomotor function in response to toxic variables. 129 Tests involving neuromuscular function in zebrafish can provide critical insight into neuromuscular dysfunction.
There are three motor function tests that portray the functional similarities of zebrafish to humans. Startle response testing examines the fish’s immediate response to stimuli. 128 The effects observed can be further analyzed to determine the lasting neurobehavioral consequences of various toxicants. In a study to measure the acute and chronic effects of ethanol exposure on different strains of zebrafish, Dlugos and Rabin identified distinctions among three strains by analyzing behavioral abnormalities or tolerances in response to ethanol exposure. Genotypic differences among three strains of zebrafish were identified based on varying responses of the central nervous system (CNS). 36 Another study conducted by Martin et al 130 analyzed behavioral changes in zebrafish embryos exposed to cyanotoxins B-methylamino-L alanine (BMAA) and microcystin leucine arginine (MCLR) and saw a dose-dependent increase in acoustic startle response activity. Both experiments observing the effects of ethanol and cyanotoxin exposure highlight the zebrafish model’s use to predict expected endpoints of toxicant exposure using the startle response.
Activity level testing is another method used to investigate toxic effects on the developmental behavior of zebrafish.38,128 These tests include but are not limited to, prolonged negative effects on spatial discrimination and response latency. 38 Results from Lopez-Patino et al showed immediate effects of hyperactivity from an initial 24 to 72-hour period of drug withdrawal and behavioral endpoints, including lasting anxiety into adulthood. 39 Additionally, Green and Planchart showed that exposure to cadmium directly influences activity levels by inducing hyperactivity and circling behavior in larvae. Further experimentation revealed this was due to inappropriate otolith and inner ear formation; hence, improper development had occurred as a result of the heavy metal. 131
The Critical Swimming Speed test measures the effects of exercise in addition to a chosen variable’s influence on the subject ranging from chemical exposure40,132 to environmental differences.41,133 The optimal swimming speeds of zebrafish have been measured by subjecting the fish to varying velocities of water flow
40
and toxicant exposure.132,134 In an effort to show the zebrafish is an efficient exercise model, oxygen consumption and routine metabolic rate were measured at increments of 25%, 50%, 75%, and 100% of critical swimming speed. Results of the study indicated that zebrafish have strikingly higher swimming speeds than twenty-two other measured fish species and that exercise-enhanced growth was not only evident in development but also during adult stages.
40
Another study measured the swimming capability of zebrafish using four tests. These distinct assessments all measured diverse variables, such as gender, water temperature, caudal fin length, and genetic strain, that may affect the swimming capability of the zebrafish model.
41
The examples of neuromuscular assessments provide evidence that the zebrafish is an excellent model for evaluating neuromuscular function. Numerous published experiments further exemplify the functionality of the zebrafish as an excellent alternative model for toxicology and other biological studies (Figure 3). A summary table of research supporting the beneficial use of zebrafish to investigate neuromuscular structure and function can be found in Table 3. Depiction of behavioral responses in zebrafish. Each form of testing is ordered according to the complexity of the assessment. Neuromuscular responses are the most basic behavioral response and lead to more complex sensory responses, which coalesce with cognitive responses. Sub-categories illustrate different methods of testing previously used to investigate behavioral endpoints. Neuromuscular Structure and Function.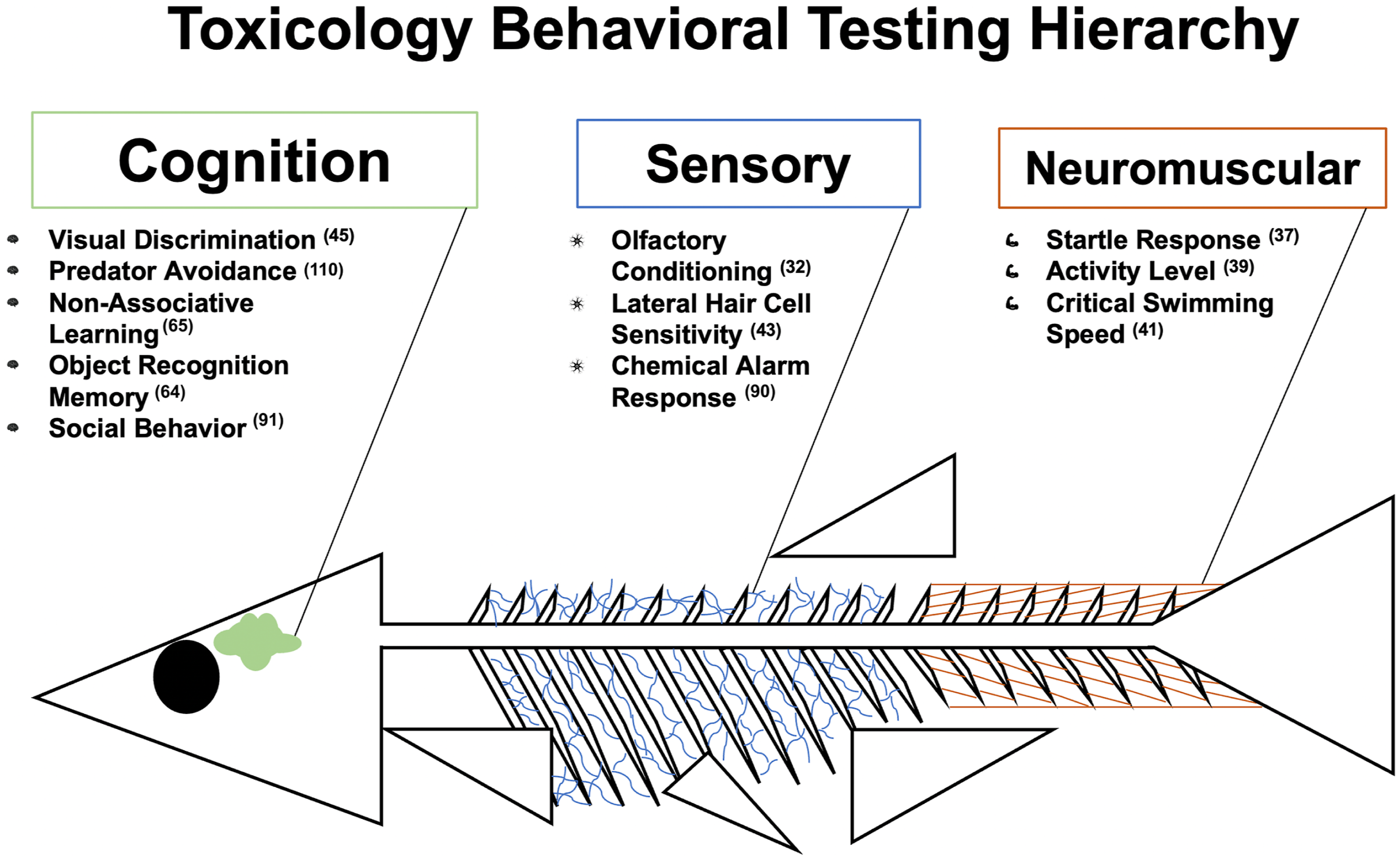
Zebrafish Mutants
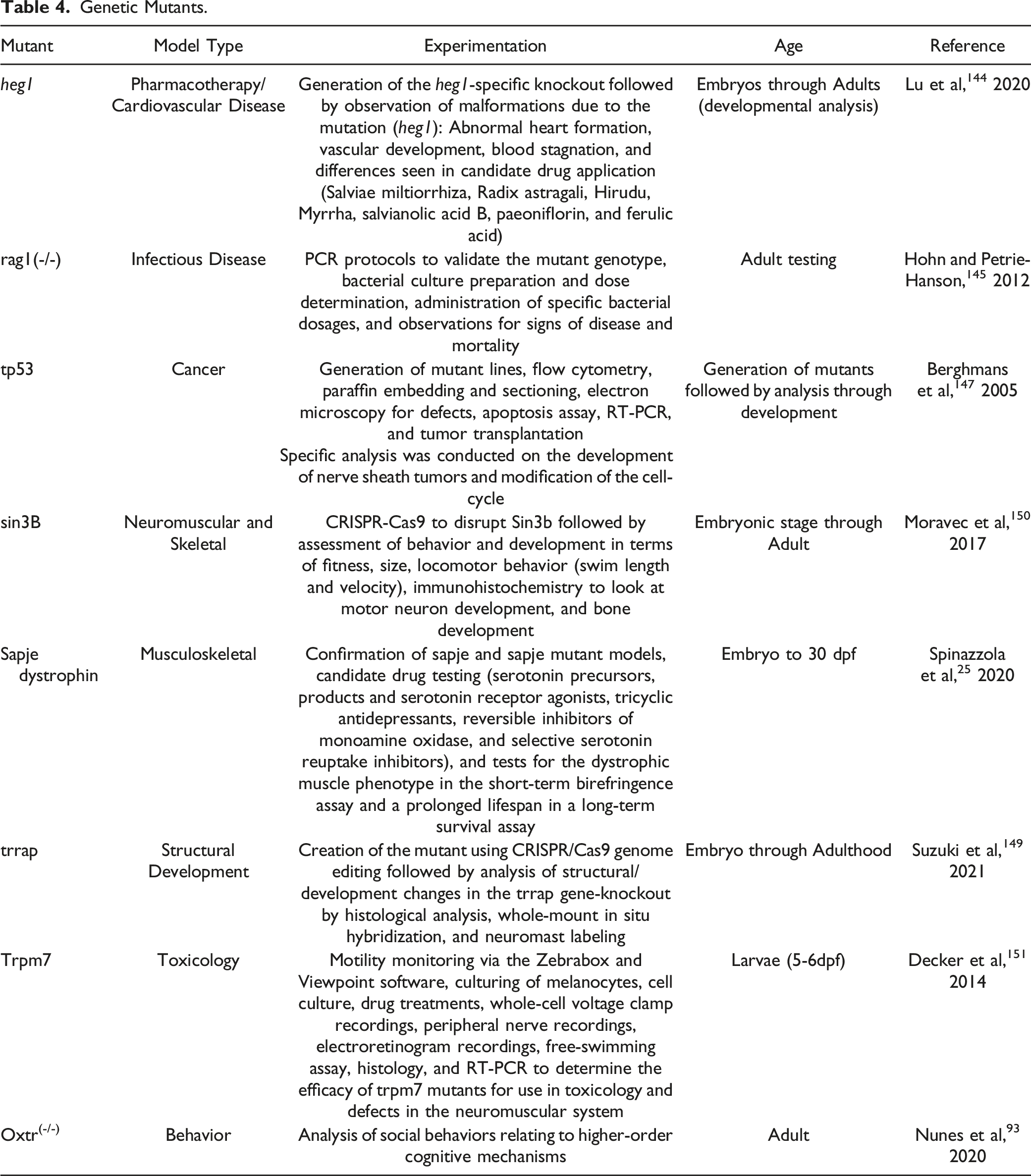
The similarities between zebrafish and humans in protein structure and function, in addition to the ease of testing their motor function, make mutant models of the zebrafish desirable for toxicology investigations. As was mentioned previously, zebrafish contain more than 70% of the human genome and 47% of human genes have a one-to-one relationship with a zebrafish ortholog. 135 Furthermore, a remarkable >82% of human genes are linked with comorbidities containing at least one orthologous zebrafish gene.124,129,136 An example of a gene homolog affecting cognitive function is the dopamine receptor (drd5) gene which is associated with working memory. When treated with a D1/D5 receptor agonist, zebrafish showed age-related declines in working memory, which is similar to what is seen in adult humans. 137 Additionally, zebrafish share three isoforms of glutamate decarboxylase and all but two of the GABAA receptor orthologs with humans. 138 The GABAA receptors serve a wide range of functions in both humans and fish, including motor control, 139 vision, 140 and learning and memory. 141 Neuromuscular gene homologs are another characteristic that makes zebrafish an attractive model for toxicological studies. For example, the tardbp gene encodes for the RNA binding TDP-43 protein, which has been investigated recently to help study human amyotrophic lateral sclerosis. Overexpression of the cytoplasmic variety of tardbp leads to neuromuscular degeneration. 130 Another neuromuscular degenerative disease is myofibrillar myopathy, which is caused by misfolded myosins. Specifically, zebrafish contain gene orthologs for two proteins associated in the formation of this disease: unc-45b and 503unc where 503unc expression is essential for the expression of other genes that promote proper muscle formation. 142 A final example of homology is seen where 23 of the 25 human genes associated with muscular dystrophy have orthologs in zebrafish. 143
Since their development, zebrafish mutant use has expanded to an array of research interests, including cardiovascular disease, 144 immunology, 145 cancer,146,147 and toxicology.2,120 The sapje dystrophin mutant is a model for exploring candidate molecules to remediate muscle degeneration in cases of human muscular dystrophy.20,25,42,148 The trrap mutant is specifically used to investigate the physiological function of the trrap gene in vertebrates. By studying this mutant, it has been observed that genetic alterations in the trrap gene have resulted in facial dysmorphisms, microcephaly, and the development of the eyes and pharyngeal arches. 149 Another mutated gene, sin3b, demonstrated a decrease in fitness, size, changes in locomotor behavior, and delayed bone development. 150
In zebrafish, mutants have the potential to be used in drug discovery as disease models. Specifically, investigations have looked at the development of trpm7 mutants. They are abnormally sensitive to 1-methyl-4-phenylpyridinium, which is a neurotoxicant that interferes with the mitochondrial electron transport chain. 151 These mutants are missing the gene which programs the trpm7 ion channel kinase.
Genetic Mutants.
Ethics
The benefit of using zebrafish in toxicology is not limited to being a comparable model to humans. They also offer an ethical solution to overcome some of the controversial aspects of scientific research. Steven Cassar et al 2 provide an application of “The Three Rs” (3Rs) concept in the use of zebrafish by detailing the benefits of connecting the model with humans in in vitro assays and mammalian in vivo studies for drug discovery. According to the European Commission Directive from 2010, experimentation in the early life stages of zebrafish can be conducted without qualifying as an “animal” research study. Independent feeding begins at 5dpf; thus, before this period, zebrafish do not qualify as an “animal” study. Because of their alignment with the 3Rs, zebrafish provide an optimal model for neuromuscular testing and are a quality alternative to animal studies.2,120
Conclusion
The use of the zebrafish model has significantly increased across several scientific fields of study. This upsurge is not merely attributed to the convenient and feasible nature of the model but primarily due to the 70% (+) genetic similarity to humans in terms of cognition, anatomical structure, and neuromuscular function. Zebrafish have been established as pivotal models to investigate cognitive behaviors such as visual discrimination, memory and learning, and social behavior. These studies have illuminated their value in neuroscience investigations with the ever-increasing rates of depression, anxiety, and stress disorders across the world. Because of these applications, toxicology/pharmacology investigations have found zebrafish promising in investigating drug-related cognitive and neuromuscular behavioral responses. The latest avenue of zebrafish research has involved the development of mutant models, which has facilitated the ability to further investigate cardiovascular abnormalities, immunological disorders, and muscular degenerative diseases. As has been evidenced, the zebrafish offer an effective alternative model in a multitude of scientific fields, and as a result, it is to be expected their use to grow exponentially.
Footnotes
Acknowledgements
The authors would like to thank the library personnel at Florida College and the institution for providing access to journal articles and guidance in attaining information for this manuscript.
Author Contributions
Clevenger, T. contributed to conception and design, contributed to acquisition, analysis, and interpretation, and drafted manuscript; Paz, J. contributed to conception and design, contributed to acquisition, analysis, and interpretation, and drafted manuscript; Hayes, W. contributed to conception, contributed to interpretation, and critically revised manuscript; Stafford, A. contributed to conception, contributed to analysis and interpretation, and critically revised manuscript; Amos, D. contributed to conception and design, contributed to acquisition, analysis, and interpretation, and critically revised manuscript. All authors gave final approval and agree to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The authors declare no potential conflicts of interest with respect to the research, authorship, or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.