
Abstract
Dietary supplements containing plant materials of Annonaceae species (Annona muricata L., A. squamosa L., A. mucosa JACQ., A. squamosa × cherimola M
Introduction
Graviola (Annona muricata L.), ata (A. squamosa L.), atemoya (A. cherimola × squamosa M
Epidemiological studies, however, linked the consumption of Annonaceae to a high prevalence of atypical parkinsonism, in Guadeloupe, 6 –8 in parts of the Afro-Caribbean and Indian population in London 9 and New Caledonia. 10 In several patients who desisted in their consumption of Annonaceae fruits, the progression of atypical parkinsonism ceased, while in 1 patient the symptoms even disappeared after a change in diet. 6 Previous studies found that graviola (A. muricata) and the North American pawpaw (Asimina triloba) contain annonacin. 11 –13 A more recent study showed that one sample of lyophilized pawpaw fruits contained not just 7.72 mg/g annonacin but also 0.162 mg/g squamocin. 14 Moreover, all of the dietary supplements and fruits analyzed in the current study were found to contain annonacin and/or squamocin when analyzed by LC-MS/MS. 15 NMR analysis also found that they all contained the α,β-unsaturated γ-lactone toxicophore that is present in neurotoxic acetogenins. 16
Annonacin, extracted from graviola, was shown to be toxic for embryonic rat primary mesencephalic 17 and striatal neurons. 18,19 North American pawpaw extracts were toxic in primary rat cortical neurons. 13 Moreover, annonacin induces neurodegeneration in rats 20 and increases tau pathology in tau transgenic mice. 21
Annonacin belongs to the group of Annonaceous acetogenins. These are highly lipophilic compounds and potent inhibitors of the mitochondrial nicotinamide adenine dinucleotide : ubiquinone oxidoreductase, the complex I of the respiratory chain. 19,22 Similar to annonacin, other acetogenins cause tau pathology and cell death in primary striatal cultures of embryonic rats in correlation with their potential to inhibit complex I. 22
Besides the strong evidence that complex I inhibition is responsible for the neurotoxic effect of Annonaceous acetogenins in vitro, also other mechanisms for cytotoxicity are discussed, including modulating histone H3 phosphorylation 23 or complexation with calcium ions. 24 Moreover, besides aceto-genins, Annonaceous plants contain other toxic substances, such as specific alkaloids. However, these have been shown to be 1000 times less toxic to neuronal cells than acetogenins. 11 It is unknown whether freely available dietary supplements claiming to contain graviola or pawpaw exert neurotoxicity.
The aim of this study was to determine the biological effects of Annonaceae containing dietary supplements on Lund human mesencephalic neurons (LUHMES) in terms of cell viability and neurotoxicity and to compare these data to results of fruit pulp and seed extracts of different Annonaceous species.
Materials and Methods
Test Materials and Chemicals
Four dietary supplements were obtained over the Internet. Two of them (S1 and S2) were labeled as containing dried graviola leaves and stems. Another was labeled as containing pawpaw twigs (S3). The fourth (S4) was labeled as containing dried graviola leaves. Graviola fruit pulps (A. muricata) were collected in São Paulo (Brazil). Atemoya (A. squamosa × cherimola) fruit pulp and seeds were collected in Belém (Pará, Brazil) and Pilar (São Paulo, Brazil). Ata (A. squamosa) and biribá (A. mucosa) fruit pulps were from Belém (Pará, Brazil). The pulp and seeds were removed from the fruits, lyophilized, and stored frozen until ready for extraction. High-purity ethyl acetate was obtained from Honeywell Burdick & Jackson (Muskegon, Michigan).
Extraction of Samples
About 10 g of each sample was mixed with HydroMatrix (Sigma Aldrich, St Louis, Missouri) to reach the 100 mL mark in the sample cell used in an accelerated solvent extractor (Dionex ASE 300; ThermoFisher Scientific, Sunnyvale, California). Then, 40 mL ethyl acetate was added. The temperature and pressure were increased to 100°C and 10.3 MPa (100 atm) over 3 minutes (static time). Next, the solvent was flushed out into a collection vessel. This was repeated 3 more times to obtain a total volume of 160 mL. The solvent was evaporated off, and the remaining residue was weighed. Stock solutions of portions of the residues obtained from the various extracts were prepared in dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, Missouri). The drug–extract ratio (DER) for each extract was calculated. The DER states how many parts of starting material are needed to produce 1 part of extract. It helps to compare different extracts and can be used to draw conclusions on the enrichment of extract ingredients.
Cell Culture
The LUHMES cells were used for the study of cell viability and toxicity and were derived from human mesencephalic tissue. They were immortalized by cloning in a v-myc vector under tetracycline control and can be differentiated to postmitotic dopaminergic neurons. 25 –27 The cells were proliferated on flasks (Nunclon Δ surface; NUNC A/S, Roskilde, Denmark) coated with poly-L-lysine (PLL; Sigma-Aldrich) 0.1 mg/mL at +4°C overnight in growth medium (Dulbecco modified Eagle medium [DMEM]/F12 [Sigma-Aldrich] with 1% N-2 supplement [Life Technologies, Carlsbad, California], and 0.04 µg/mL basic fibroblast growth factor [R&D Systems, Minneapolis, Minnesota]). The cells were always kept at 37°C. For experiments, cells were seeded in growth medium on multiwell plates (Nunclon DELTA surface; NUNC A/S) coated with 0.1 mg/mL PLL at 4°C overnight and washed 3 times with phosphate-buffered saline (PBS; Life Technologies) followed by coating with 5 µg/mL fibronectin (Sigma-Aldrich) for at least 6 hours in the incubator (37°C, 5% CO2). The wells were then washed once with PBS and air dried before plating at a density of 55 000/cm2. The cells were kept at 37°C, 5% CO2, and 100% humidity. Twenty-four hours after plating, medium was changed to differentiation medium (DMEM/F12 with 1% N-2 supplement, 1 µg/mL tetracycline, 0.49 mg/mL dibutyryl cyclic AMP [Sigma-Aldrich] and 2 ng/mL GDNF [R&D Systems]). The cells were differentiated for 8 days with changes of 50% of the differentiation medium every second day.
Treatment of Cells
After 8 days of differentiation, the medium was changed to low glucose differentiation medium composed of DMEM with 250 µmol/L of glucose with the same supplements as described above. This was done in accordance with previous studies investigating the toxic effects of acetogenins or other complex I inhibitors in primary cell cultures 18,19 and LUHMES cells. 28
After medium change, the cells were kept at 37°C, 5% CO2, for at least 1 hour. Thereafter, the cells were treated with ethyl acetate extracts in different concentrations dissolved in DMSO (AppliChem, Darmstadt, Germany) and diluted into low glucose differentiation medium. The concentration of DMSO never exceeded 0.1%. Negative control cells were treated with the same amount of low glucose differentiation medium containing the same amount of DMSO. Experiments were repeated 3 times with 4 different wells treated per condition in each experiment.
Measurement of Toxicity
The 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was performed 48 hours after incubation with different samples as previously described. 28,29 MTT of 5 mg/mL (Sigma-Aldrich) was dissolved in PBS. This solution was added 1:10 to the medium of each well of the cell culture plate to a concentration of 0.5 mg/mL. After 1 hour of incubation at 37°C, the MTT solution and medium were removed and the plate was frozen at −80°C for 24 hours. Thereafter, the plate was thawed and 300 µL of DMSO were added to each well. After shaking for 5 minutes, 100 µL from each well was transferred to a transparent U-shaped-bottom 96-well plate, and the absorption was measured at a wavelength of λ = 590 nm using a FLUOstar Omega plate reader (BMG Labtech, Ortenberg, Germany) in absorbance mode. Values were normalized to negative control cells.
As a second assay, the lactate dehydrogenase (LDH) released from dying cells into the culture medium was measured 48 hours after treatment of the cells with the extracts using the CytoTox-ONE Homogeneous Membrane Integrity Assay (Promega, Fitchburg, Wisconsin) according to the manufacturer’s instructions with a FLUOstar Omega plate reader (BMG Labtech) in fluorescence mode with an excitation of λ = 544 nm and an emission of λ = 590 nm. Data were normalized to data from cells lysed by treating them with the lysis buffer of the CytoTox-ONE Homogeneous Membrane Integrity Assay for 30 minutes as positive controls (maximum LDH-release).
Statistics
One-way analysis of variance (ANOVA) followed by Tukey post hoc test was used to determine significance using Prism 6 for Windows (GraphPad Sofware Inc, La Lolla, California).
Results
Pressurized Liquid Extraction
Using an accelerated solvent extractor, we carried out a pressurized liquid extraction with ethyl acetate and were able to receive extraction yields between 1.9% and 23% (Table 1). Ethyl acetate preferably solubilizes lipophilic substances in the plant material and is known to extract neurotoxic ingredients of Annonaceae as annonacin. 13 Expectedly, the DERs that we determined for the fruit pulp ethyl acetate extracts are very high (Table 1), indicating that the fruit pulps only contain a low amount of lipophilic substances. By contrast, the extracts of the leaves, stems, twigs, and seed material had a relatively low DER, showing that a larger amount of the contained substances are more lipophilic.
Overview of Produced Ethyl Acetate Extracts.
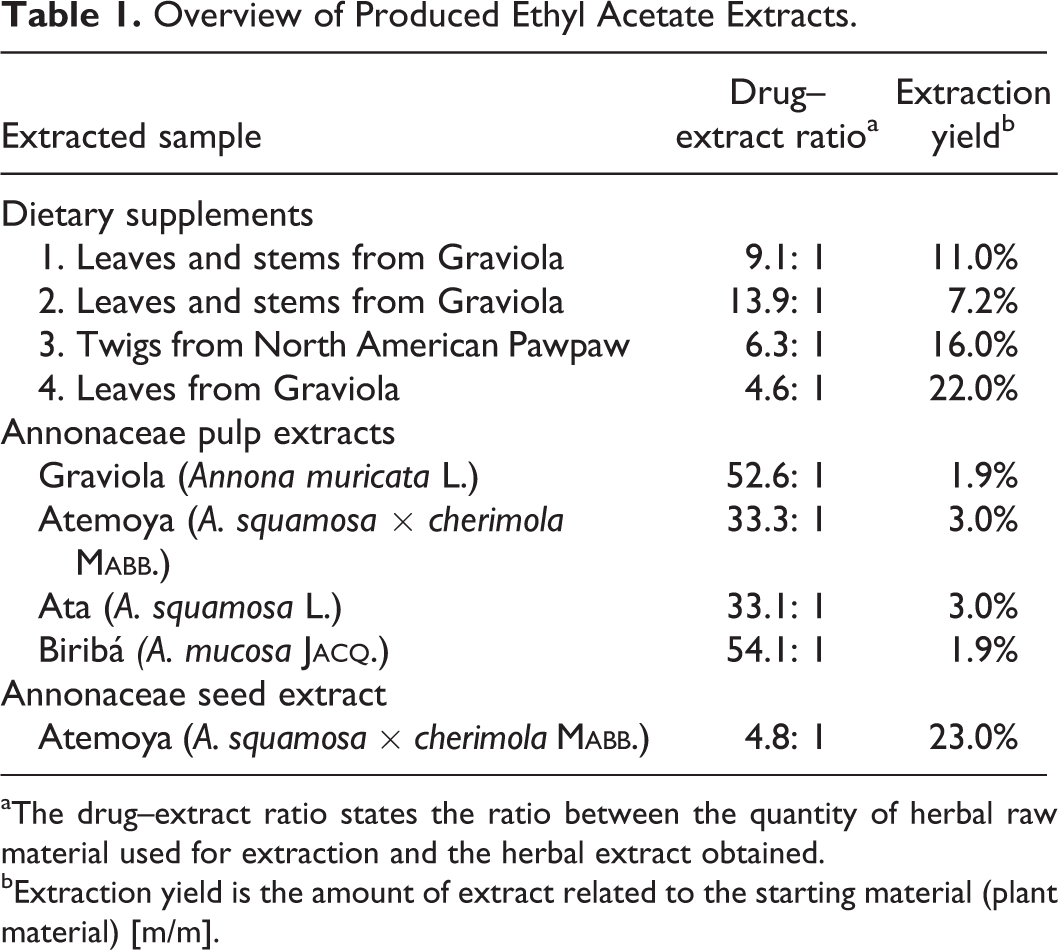
aThe drug–extract ratio states the ratio between the quantity of herbal raw material used for extraction and the herbal extract obtained.
bExtraction yield is the amount of extract related to the starting material (plant material) [m/m].
Neurotoxicity of Dietary Supplements
In order to test the toxicity of extracts of 4 different dietary supplements claiming to contain different parts of Annonaceous plants, graviola (A. muricata L.) and the North American pawpaw (Asimina triloba (L.) D
Concentrations of Annonacin and Squamocin Found in Samples Evaluated in This Study. 15
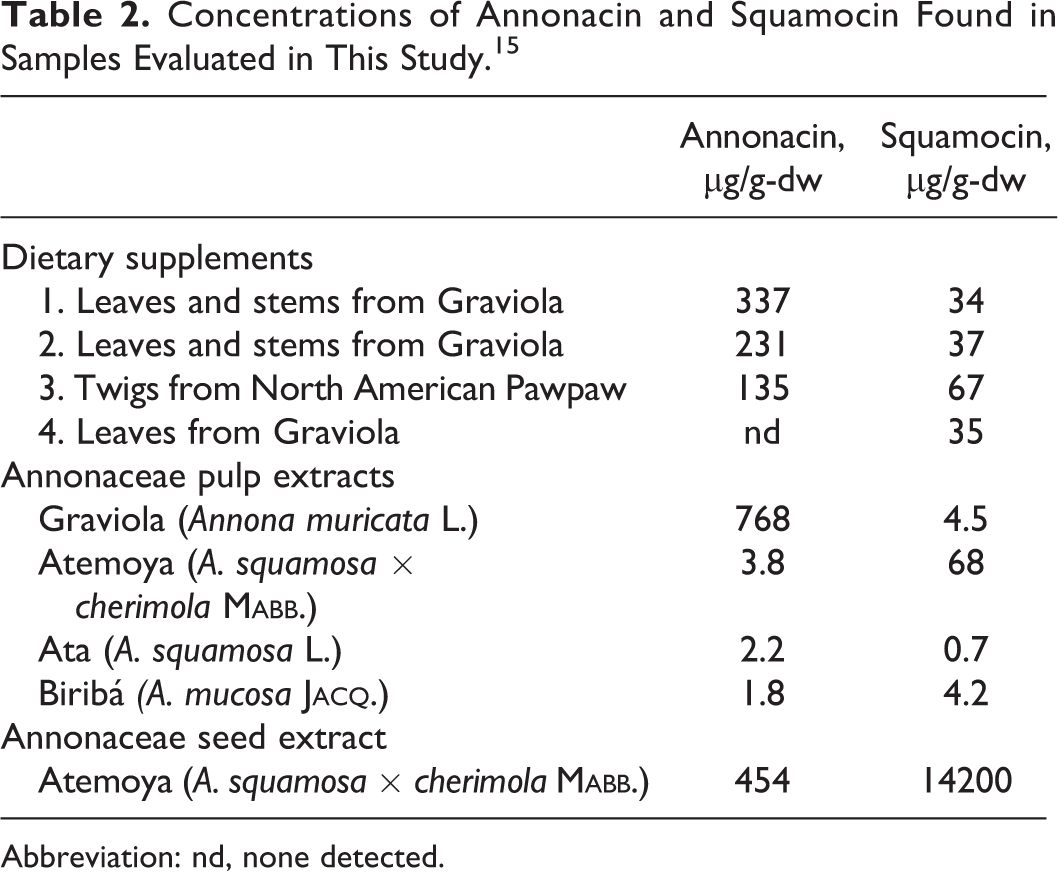
Abbreviation: nd, none detected.
The treatment of LUHMES cells with extracts from 3 of the 4 dietary supplements led to a reduced cell viability. Already at a concentration of 0.1 µg/mL, the extract of supplement S1 reduced the MTT level to 57.5% ± 19.8% of that of negative control. At concentrations of 1.0 and 10 µg/mL, the MTT level was further reduced to 12.3% ± 2.1% and 9.0% ± 0.7% (P < .0001 vs negative control, Figure 1B [S1]). With 1 µg/mL of the extract from supplement S2, the MTT level was reduced to 48.4% ± 4.6% (P < .0001 vs negative control), while a concentration of 10 µg/mL further reduced the MTT level to 13.4% ± 2.4% (P < .0001 vs negative control; Figure 1B [S2]). While 1 µg/mL of the extract from supplement S3 did not decrease the MTT signal, a concentration of 10 µg/mL reduced the MTT level to 37.1% ± 10.9% (P < .0001 vs negative control; Figure 1B [S3]). Concentrations of up to 10 µg/mL of supplement S4 did not decrease the cell viability (Figure 1B [S4]), indicating that this supplement was not toxic in the tested range. The 3 supplements (S1, S2, and S3) that reduced cell viability all led to cell death as measured by LDH release into the cell culture medium. In correspondence to the data from the MTT assay, the extract from supplement S1 was the most toxic, leading to a significant LDH increase in 37.5% ± 2.5% at a concentration of 1 µg/mL, compared to the LDH level in negative controls of 21.1% ± 0.3% (P = .003). Ten µg/mL of this extract led to an increase in the LDH level to 78.9% ± 4.3% (P < .0001 vs negative control), indicating almost maximal cell death (Figure 1B [S1]). While treatment with 1 µg/mL of the extract from supplement S2 was not toxic, we observed a significant increase in the LDH level with 10 µg/mL to 87.0% ± 2.6% compared to 23.5% ± 1.0% in negative controls (P < .0001; Figure 1B [S2]). We also did not observe significant cell death with 1 µg/mL of the extract from supplement S3, while 10 µg/mL of this extract led to an increase in the LDH level to 62.7% ± 12.3% compared to 27.1% ± 2.5% in negative controls (P = .001, Figure 1B [S3]). In correspondence to the data from the MTT assay, the extract from supplement S4 did not induce cell death in the tested concentration range (Figure 1B [S4]).
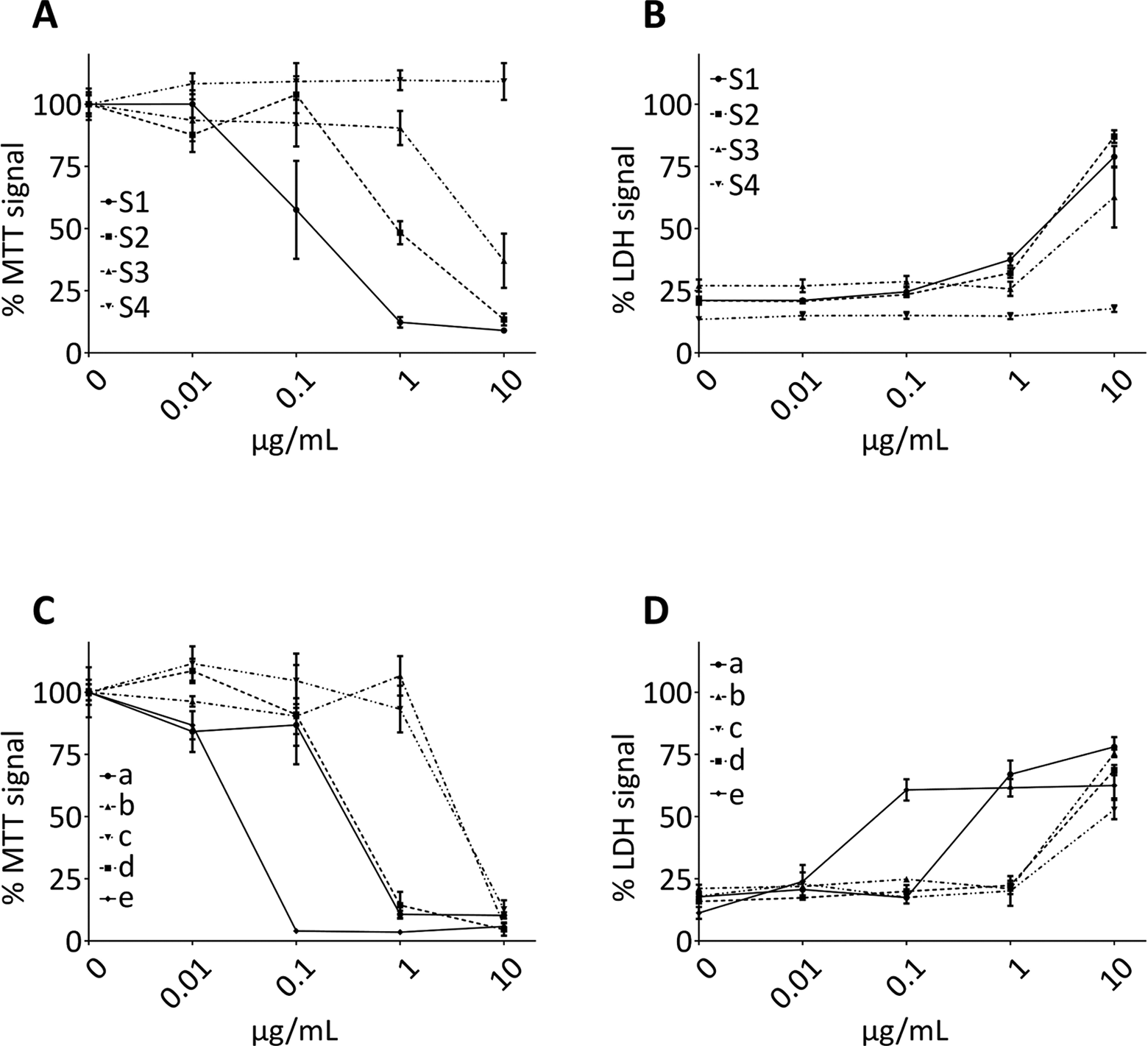
Toxicity of Annonaceous ethyl acetate extract. Human mesencephalic (LUHMES) cells were treated for 48 hours with different concentrations of ethyl acetate extracts from 4 Annonaceae-containing dietary supplements (S1–S4) and from Annonaceous fruit pulp (a: Annona muricata, b: Annona squamosa, c: Annona mucosa, d: Annona cherimola × squamosa) and seeds (e: Annona cherimola × squamosa). Cell viability was measured by relative MTT levels (A) and cell death measured by LDH release (B) after treatment with 4 dietary supplements and corresponding data after treatment with Annonaceous pulp and seeds extracts (MTT: C, LDH: D). Data are normalized to the MTT signal measured in DMSO-treated control cells. Data from the LDH release assay are normalized to lysed cells (maximal cell death) as positive controls. For details and statistics, see text. LDH indicates lactate dehydrogenase; DMSO, dimethyl sulfoxide; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Neurotoxicity of Annonaceous Fruit Pulps or Seeds
We measured the MTT levels in LUHMES cells and the release of LDH into the cell culture medium after treatment with extracts of fruit pulps in the same way that we tested the extracts from the 4 dietary supplements. We tested fruit pulp extracts of A. muricata L., A. squamosa L., A. mucosa J
Discussion
The consumption of herbal tea and fruits from the Annonaceae family (A. muricata and A. squamosa) was linked to atypical parkinsonism on Guadeloupe.
6
–8
Previous studies showed that acetogenins, which are long-chained lipophilic inhibitors of the mitochondrial complex I and found in Annonaceous species, are highly neurotoxic in vitro
18,19
and in vivo.
21
Nevertheless, different parts of Annonaceae are sold as dietary supplements in the United States and are promoted to have anticancer effects and to increase general health. In the present study, we demonstrated that extracts from these freely available dietary supplements are toxic to neuronal cells by an extent comparable to the neurotoxicity of extracts from fruit pulp and seeds of different Annonaceous species (A. muricata L., A. squamosa L., A. mucosa J
We used ethyl acetate for the extraction procedure to solubilize lipophilic ingredients of Annonaceous plant material including neurotoxic acetogenins. 13 The amount of raw material used to obtain a certain amount of extract was lower in the dietary supplements (DERs 4.6:1 to 13.9:1) than in the pulp of 4 different Annonaceous species (DER 33.1:1 to 52.6:1). The reason for that was likely that fruit pulp contains more hydrophilic material than the dietary supplements that contained leaves, stems, and twigs of the plants. However, our data suggest that the content of lipophilic ingredients from Annonaceous species, including potentially toxic lipophilic acetogenins, is higher in the plant material used for the dietary supplements than for pulp fruit extracts. Dietary supplements S1, S3, and S4 had an extraction yield similar to Atemoya seeds. This implies that they contain similar amounts of lipophilic components.
The toxicities of the extracts from dietary supplements and Annonaceous fruit extracts and seeds were measured with 2 different assays, MTT and LDH release. The MTT assay is used as a cell viability assay that is affected by changes in mitochondrial oxidative phosphorylation as well as other cellular pathways, including glycolysis. 32 However, it does not necessarily indicate cell death. On the other hand, LDH is only released into the cell medium by dying cells, when cell membranes become leaky. Therefore, it is a good tool to measure cell death.
Of the 4 dietary supplements, 2 (S1 and S2) led to a gross loss of cell viability, as measured by MTT assay, at a concentration of 1 µg/mL. Supplement S1 led to a reduction in the signal by 87.7% ± 2.1% and S2 by 51.7% ± 4.6% (Figure 1A). Moreover, supplement S1 already showed significant cell death at this concentration (Figure 1B). With the higher concentration of 10 µg/mL, 3 supplements showed marked reduction in cell viability and cell death. This observation is in line with previous observations made with several acetogenins that concentrations needed to impair the cellular energy metabolism were slightly lower than concentrations that lead to cell death. 19 This suggests that impairment of energy metabolism can be compensated for a while before ultimately resulting in cell death. The concentrations of the extracts from the dietary supplements which led to reduced cell viability and cell death were in the same range as the concentrations of the extracts from pulp of A. squamosa and A. cherimola × squamosa, which also induced cell death at 10 µg/mL. On the other hand, the extract from pulp of A. muricata was slightly more toxic, leading to marked cell death at 1 µg/mL. Interestingly, the extract from seeds of A. cherimola × squamosa seeds already led to marked decrease in cell viability and induced cell death at 0.1 µg/mL. This suggests that the seeds contain even more toxic ingredients than the fruit pulp. Also, dietary supplements S1 and S2 (containing leaves plus stem from A. muricata) were toxic at the tested concentrations, while dietary supplement S4 that contains leaves but not stems from A. muricata was not toxic at the toxic concentrations. This suggests that the stems contained more toxic compounds than the leaves.
In advertising texts, the ingredients of the Annonaceous dietary supplements are described as generally healthy, reasoned due to their long tradition in herbal medicine. Some advertisements even concede that the acetogenins, which are present in Annonaceous fruits, impair the ATP production in mitochondria. It is suggested that this might be the case for specific cell types, especially cancer cells. This is supported by several reports showing that extracts from Annonaceae are toxic to cancer cells. 33 –36 In line with that, it was recently reported that A. muricata extracts inhibited the growth of a promyelocytic leukemia cell line in vitro with an IC50 between 6 and 49 µg/mL. 37 The data may not directly be comparable to the data we obtained here. A different solvent was used to extract the samples, and different cell culture media were used to grow tumor cell lines and neuronal cells. Still, it is worth noting that the concentrations of A. muricata extract that led to growth inhibition in the promyelocytic leukemia cell line were much higher than the concentrations of A. muricata extract that led to reduced cell viability and cell death in LUHMES neurons here.
The effects of acetogenins on the cell viability of tumor cells, which are essentially dependent on high levels of energy, are undisputed. However, there is currently no evidence that the cytotoxic effect of acetogenins is selective or specific for cancer cells. In contrast, the data presented here and by previous reports show that extracts from Annonaceae or purified acetogenins are especially toxic to neuronal cells. The toxic effects of annonacin on rat primary mesencephalic 17 and striatal cells 18,19 as well as on human neuronal cells 28 have been shown previously. Moreover, in vitro experiments revealed that treatment with Annonaceae acetogenins increased tau phosphorylation 18,19 and led to an isoform shift from 3R-tau to 4R-tau, 28 both contributing to the pathophysiology of neurodegenerative tauopathies.
We treated the cells with ethyl acetate extracts from dietary supplements, fruit pulp, or seeds of Annonaceae. These extracts contain a complex mixture of lipophilic substances, and the toxic effect that we observed can therefore not exclusively be attributed to acetogenins. A previous report showed that crude ethyl acetate extracts of A. muricata pulp contain up to 10% annonacin, 13 but it is very likely that the other 90% contain other potentially toxic compounds, for example, other acetogenins or specific alkaloids that contributed to the observed effect. By the intake of the dietary supplements, one ingests all toxic ingredients, regardless of the specific composition. Because of this, we investigated full extracts and not solely isolated compounds in the present study.
In summary, our data clearly demonstrate that dietary supplements containing twigs or leaves from Annonaceous species are toxic to neuronal cells in the same manner as fruit pulp extracts from Annonaceae that are known to contain huge amounts of neurotoxic acetogenins. This finding should be taken serious because these dietary supplements are advertised as being health enhancing and helpful against malignant diseases. Moreover, they are designated for chronical intake. The consumption of non-evidence–based dietary supplements of Annonaceae could contribute to the occurrence of neurodegenerative disorders in the same way as the consumption of Annonaceous fruits in Guadeloupe.
Footnotes
Acknowledgments
We acknowledge support by the German Research Foundation (DFG, HO2402/6-2), the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico do Brasil), FAPERJ (Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro), and FAPESPA (Fundação de Amparo à Pesquisa do Estado do Pará).
Authors’ Note
This work should not be taken as reflecting FDA policy or regulations. Matthias Höllerhage, and Thomas W. Rösler equally contributed to this work.
Author Contribution
Höllerhage, Matthias, Höglinger, Guenter, and Rösler, Thomas contributed to conception and design, contributed to acquisition, analysis, and interpretation. Berjas, Magda contributed to conception and design and analysis. Luo, Rensheng contributed to conception, acquisition, and interpretation. Tran, Kevin contributed to conception, acquisition, analysis, and interpretation. Richards, Kristy contributed to conception and analysis; Sabaa-Srur, Armando, Maia, JGM, De Moraes, MR, and Godoy, HT contributed to conception and design and acquisition. Smith, RE contributed to conception and design, acquisition, analysis, and interpretation. All authors drafted manuscript, critically revised manuscript, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.