
Abstract
Aspartame is one of the most widely used artificial sweeteners globally. Data concerning acute neurotoxicity of aspartame is controversial, and knowledge on its chronic effect is limited. In the current study, we investigated the chronic effects of aspartame on ionic homeostasis and regional monoamine neurotransmitter concentrations in the brain. Our results showed that aspartame at high dose caused a disturbance in ionic homeostasis and induced apoptosis in the brain. We also investigated the effects of aspartame on brain regional monoamine synthesis, and the results revealed that there was a significant decrease of dopamine in corpus striatum and cerebral cortex and of serotonin in corpus striatum. Moreover, aspartame treatment significantly alters the tyrosine hydroxylase activity and amino acids levels in the brain. Our data suggest that chronic use of aspartame may affect electrolyte homeostasis and monoamine neurotransmitter synthesis dose dependently, and this might have a possible effect on cognitive functions.
Introduction
Currently, obesity and diabetes are emerging as the most important health care issues for both developed and developing countries. Excessive calorie intake is considered as one of the main culprits in the development of these conditions. In recent times, artificial sweeteners are paid a special attention among food additives, as their use enables a sharp reduction in sugar consumption and a significant decrease in caloric intake while maintaining the desirable palatability of foods and soft drinks. 1
Aspartame (
Previous studies have shown that chronic consumption of aspartame produces impairment in learning and memory processes. 9 –11 Ionic involvement has been suggested to play a role in spatial learning and memory. 12 In an earlier in vitro study, Schulpis et al 13 have reported that aspartame significantly decreased the Na+-K+-ATPase activity in the erythrocyte membrane. Na+-K+-ATPase is crucial for maintaining ionic homeostasis and cell volume in neurons. 14 Moreover, evidence suggests that impaired Na+-K+-ATPase function can be linked to apoptosis. 15 Brain function is critically dependent on the ionic homeostasis in both the extracellular and the intracellular compartments. Modulation of Na+-K+-ATPase activity directly affects neurotransmitter signaling, neural activity as well as the whole cognitive functions. However, the information on the effect of long-term exposure of aspartame on ionic homeostasis is lacking.
On the other hand, it was also reported that administration of aspartame can alter phenylalanine levels in the brain. 16 Tyrosine, which is derived from phenylalanine and tryptophan, serves as the precursor for the synthesis of monoamines in the brain. Monoamines, including the catecholamines (dopamine, epinephrine, and norepinephrine) and serotonin, are a group of important neurotransmitters that regulate a wide array of cognitive and behavioral functions in brain. An alteration in the precursor amino acids has the potential to impact monoamine neurotransmitter synthesis in the brain. In addition, the effect of aspartame on regional brain monoamines has yielded inconsistent results with increased norepinephrine and dopamine in hypothalamus, 17 increased serotonin in striatum, 18 or increased norepinephrine in cerebral cortex and amygdala. 19 Nevertheless, the reports related to the effect of aspartame on monoamine concentrations were limited to either single-dose or short-term exposure. The effects of the food additives are visible only with time, thus single or short-term studies are not sufficient to evaluate the adverse effects.
The present study used 2 different doses of aspartame. The doses in this study were selected in order to address the safety of aspartame in the acceptable daily intake (ADI) level and to explore the feasibility of aspartame to produce adverse effects during overconsumption. According to Food and Drug Administration, the ADI of aspartame is 50 mg/kg body weight. Acceptable daily intake is derived from the no observed adverse effect level (NOAEL) dose, which is the highest dose level causing no adverse effects in animal toxicity studies. The NOAEL dose of aspartame is taken as 5000 mg/kg body wt. But recent reports demonstrate that even the ADI dose could produce adverse effects. 7,8,20 Additionally, instead of following the ADI recommendation, there is also a portion of the population, such as diabetic patients and children, who are likely to consume a level in excess of the ADI. 21 The consumption of aspartame is increasing heavily. It is likely that the actual intake would be higher than the ADI. In this study, the higher dose (1000 mg/kg body wt) was selected according to the previous reports, which used a similar dose level. 4,22 Dosages in the range of 500 to 1000 mg/kg have routinely been used in animal studies involving aspartame. 23 Due to the widespread inclusion of artificial sweeteners in many foodstuffs, the actual dosage levels experienced by people are extremely difficult to estimate accurately. In fact the higher dose used in this study is much lower than the NOAEL dose. It has been suggested that rats metabolize aspartame at a much greater rate than humans. Thus, in comparative studies in humans and rats, rodents should be given high doses of aspartame to simulate the metabolic disposition of smaller amounts in humans. 24
Since data on the aspartame acute studies are controversial and knowledge on its long-term effect is much more limited, we aimed in the present study to further explore aspartame neurotoxicity after chronic administration in experimental rats. The present study focused on the chronic effects of aspartame on ionic homeostasis, apoptosis, amino acids, and brain regional monoamine synthesis.
Materials and Methods
Chemicals
Aspartame (98%) was purchased from HiMedia Chemicals, India. Oubain, glucose-6-phosphate dehydrogenase, catalase, dithiothreitol (DTT), NADH, 2-amino-6,7-dimethyl-4-hydroxy-5,6,7,8-tetrahydropteridine (DMPH4), and dihydropteridine reductase were of analytical grade purchased from Sigma Chemicals (St Louis, Missouri). All other chemicals used in this experiment are of analytical grade purchased from Sisco Research Laboratories (SRL), India.
Animals
All experiments were performed in healthy male Wistar rats with an initial weight of 175 to 200 g purchased from Govt. Veterinary College, Kerala, India. The animals were housed in groups in standard laboratory cages (5 rats per cage) in a temperature controlled room (22 ± 2°C) with a 12-hour light–dark cycle. Rats received standard laboratory chow (Hindustan Lever Ltd, India) and tap water ad libitum. The standard chow contained, as a percentage of calories, 65% carbohydrate, 12% fat, and 23% protein. Prior to initiation of dosing, all rats were acclimated for 7 days and evaluated for weight gain and any gross signs of disease or injury. Animal protocols were approved by the Institutional Animal Ethical Committee of the School of Biosciences, Mahatma Gandhi University, Kerala, India (Approval No: B2442009/10).
Experimental Design
Animals were randomly divided into 3 groups (n = 10/group). Group 1, receiving equivalent amount of vehicle alone, served as the control group. Animals of groups 2 and 3 received oral intubation of aspartame dissolved in normal drinking water at a low dose of 50 mg/kg body wt and a high dose of 1000 mg/kg body wt, respectively, for 180 consecutive days. Body weight was measured regularly to adjust the dose and achieve a constant intake of aspartame. Food consumption was measured daily from 1 week prior to administration until the end of the study.
Analytical Procedure
After 180 days, rats were killed by decapitation, the brain was quickly removed, cleaned, weighed, and chilled on crushed ice, there after these were rinsed in ice-cold saline. For determination of enzymatic activity, the frozen brains were immediately homogenized in 50 mmol/L cold Tris–HCl, pH 7.4. The homogenates were centrifuged at 4000g for 10 minutes to yield the low-speed supernatant fractions for biochemical assays. Protein determination was carried out by the method of Lowry et al. 25 All spectrophotometric determinations were carried out by a Ultraviolet–visible spectrophotometer (Hitachi U 2800, Japan).
Determination of Membrane-Bound Adenosine Triphosphatases
Ion-dependent adenosine triphosphatases (ATPases) are enzymes that play an important role in intracellular functions and are considered to be a sensitive indicator of toxicity. The activity of Na+-K+-ATPase was assayed by following the method of Carfagna et al. 26 The reaction mixture contained 50 μL of sample, 3 mmol/L MgCl2, 125 mmol/L NaCl, 20 mmol/L KCl, and 50 mmol/L Tris–HCl, pH 7.4, in a final volume of 500 μL. The reaction was initiated by the addition of ATP (3 mmol/L). Control samples were carried out under the same conditions with the addition of 0.1 mmol/L ouabain. The samples were incubated at 37°C for 30 minutes; the incubation was stopped by adding trichloroacetic acid solution (10% TCA). The activity of Na+-K+-ATPase was calculated by the difference found between 2 assays (with and without ouabain). Released inorganic phosphate (Pi) was measured by the method of Fiske and Subbarow. 27 The enzyme activity was expressed in µmol/L of Pi formed/min/mg protein.
The activity of Ca2+-ATPase in the brain tissue was assayed as described previously. 28 In brief, 0.1 mL of tissue homogenate was added to a mixture containing 0.1 mL of 125 mmol/L Tris–HCl buffer (pH 8), 0.1 mL of 50 mmol/L CaCl2, and 0.1 mL of 10 mmol/L ATP. The contents were incubated at 37° C for 15 minutes. The reaction was then arrested by the addition of 0.5 mL of ice-cold 10% TCA and centrifuged. Released inorganic phosphate (Pi) was measured by the method of Fiske and Subbarow. 27 The activity of Ca2+-ATPase was expressed in µmol/L of Pi formed/min/mg protein.
Determination of Electrolyte Contents
Concentration of Na, K, and Ca in the brain samples was determined by inductively coupled plasma optical emission spectrometry (Perkin Elmer, Massachusetts). About 0.3 g of brain tissue was weighed into a glass vessel and 3 to 8 mL of HNO3 (ultrapure grade) was added for digestion. Samples were digested simultaneously with optimized microwave digestion system at 130°C for 3 hours. Then, 2 mL of H2O2 was added, and samples were heated again for 1 hour. After digestion, the solution was transferred into a volumetric flask and made up to 25 mL with ultra-pure water and analyzed. Electrolyte concentrations were expressed as µg/g tissue.
Apoptosis Detection
Apoptosis is a highly regulated process that involves a series of molecular event that lead to cell death. To detect apoptosis in brain, terminal deoxynucleotidyl transferase-mediated 2′-deoxyuridine, 5′-triphosphate (dUTP) nick-end labeling (TUNEL) assay was performed. The TUNEL assay is an established method for detecting apoptosis. The paraffinized tissue sections were dewaxed with xylene and rehydrated through a series of decreasing concentrations of ethanol 20 g/mL. The slides were partially digested with proteinase K (10 μg/mL; Boehringer Mannheim, Mannheim, Germany) for 10 minutes, with 0.3% H2O2 in methanol for 10 minutes, and 0.1% Triton X-100 in 0.1% sodium citrate for 2 minutes on ice. Then, the sections were treated with the DeadEnd Fluorometric TUNEL System assay kit (Promega, Wisconsin). Slides were examined under Nikon E-600 fluorescence microscope (Nikon Instruments Inc, California).
Brain Amino Acid Levels
Amino acids were determined according to the procedure described by Joseph and Marsden 29 with slight modifications. Tissue samples were weighed and then homogenized by sonication in a solution of 0.1 mol/L perchloric acid. After that, samples were centrifuged at 16 000g (4°C) for 15 minutes. Supernatant was filtered through 0.22 mm nylon filters and conserved at 4°C until analysis. The amino acid samples were subjected to high-performance liquid chromatography (HPLC) analysis after derivatization with orthophtaldialdehyde-thiol reagent (orthophtaldialdehyde-thiol reagent was made up by dissolving 27 mg o-phthaldialdehyde in 500 μL absolute alcohol. Then 5 mL of 0.1 mol/L sodium tetraborate (pH 9.5) was added, followed by 50 μL mercaptoethanol. The standard stock solutions of phenyalanine, tyrosine, tryptophan, aspartic acid, and glutamic acid were prepared in perchloric acid and stored at −20°C. A series of working solutions with different concentrations were prepared by diluting each standard stock solution with the perchloric acid before use. High-performance liquid chromatography was performed using a Waters Alliance 2690XE equipped with an autosampler and a Waters 474 programmable fluorescence detector (Waters, Milford, Massachusetts). Data were acquired and processed using the Empower Pro software package from Waters. Separation was carried out on a Symmetry C18 column (4.6 × 150 mm, 3.5 µm) coupled with a guard column. The sample and column compartments were maintained at 4°C and 30°C, respectively, for all analyses, and the flow rate was 0.5 mL/min in a gradient of the mobile phase (buffer A: methanol [80%] and phosphate buffer; buffer B: methanol [20%]), and the duration of each analysis was 45 minutes. Mobile phases were filtered through a 0.2-µm filter prior to use. The eluted amino acids were monitored using fluorescence excitation at 228 nm and fluorescence emission at 306 nm from 0 to 4.5 minutes for tyrosine, at 210 and 300 nm from 4.5 to 8 minutes for phenylalanine, at 285 and 353 nm for tryptophan, and at 330 and 365 nm and 440 and 530 nm for aspartic acid and glutamic acid excitation and emission wavelength, respectively, by the programmed wavelength detection setting. The amino acid concentrations were quantitatively determined by relating their chromatographic peak areas to those obtained for a known standard mixture. Concentrations were expressed as nmol/L/g tissue.
Tyrosine Hydroxylase
Tyrosine hydroxylase activity was measured by the method of Shiman et al.
30
The incubation mixture contained 0.25 mL
Tryptophan Hydroxylase
Tryptophan hydroxylase activity was determined as described previously.
31
In brief, the complete reaction mixture volume of 0.3 mL contains the following components: 50 mmol/L of Tris–acetate, pH 7.6, 2 mmol/L DTT, 50 µg catalase, 250 µmol/L
Monoamine Neurotransmitter Levels
Monoamines (dopamine, epinephrine, norepinephrine, and serotonin) were estimated by HPLC with electrochemical detector by the method of Beyer et al. 32 Brains were rapidly removed and dissected into 2 regions: cerebral cortex and corpus striatum. The tissue samples were weighed and stored at −20°C until analysis. Brain samples were homogenized in 0.1 mol/L perchloric acid. After that, the samples were centrifuged at 15 000g for 15 minutes. The supernatant was further filtered through 0.25-μm nylon filters before injecting in the HPLC injection pump. Waters standard system consisting of a high-pressure isocratic pump, a 20 μL sample injector valve, C18 reverse phase column,, and electrochemical detector was used. Mobile phase consisted of 0.15 mol/L NaH2PO4, 0.25 mmol/L EDTA, 1.75 mmol/L 1-octane sulfonic acid, 2% isopropanol, and 4% methanol (pH 4.8). Samples (20 μL) were injected manually. Data were recorded and analyzed with the help of Empower software Waters Milford, MA, USA. Electrochemical conditions for the experiment were +0.800 V, sensitivity ranges from 1 to 100 nA. Separation was carried out at a flow rate of 1 mL/min. Concentrations were expressed as ng/g tissue.
Statistical Analysis
The results were expressed as means ± standard deviations. Statistical differences between control and treated brains were done using 1-way analysis of variance followed by least significant difference post hoc multiple comparison test to compare means between the different treatment groups. Values with P < 0.05 were considered significant.
Results
Membrane-Bound ATPases
Table 1 shows the effect of aspartame administration on membrane-bound ATPases activity in the brain. A decrease in the activity of Na+-K+-ATPase was observed in the 1000 mg/kg aspartame-administered group (P < 0.001) compared to the control group, whereas no statistically significant change was observed in low-dose treated group. The activity of Ca2+-ATPase remained unaffected in either of the aspartame-administered groups.
Effect of Aspartame on Membrane Bound ATPases.a
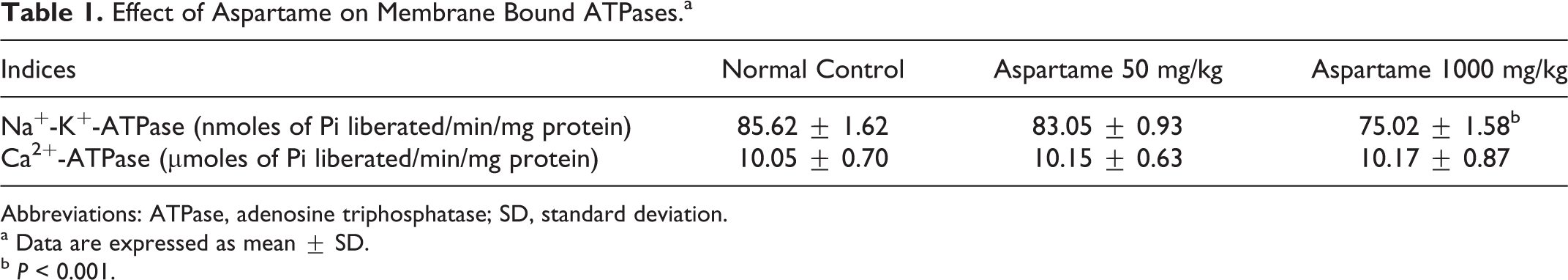
Abbreviations: ATPase, adenosine triphosphatase; SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.001.
Electrolyte Contents
The contents of Ca, Na, and K were determined in the brain tissue. These elements are important building blocks of cells and play important roles in neuronal excitability and neural transmission. The contents of Ca, Na, and K in the rat brain are listed in Table 2. It can be seen that exposure to aspartame has obviously changed the electrolyte contents in the rat brain. Administration of aspartame at high dose has significantly increased the Na (P < 0.001) content and decreased the K (P < 0.001) content in rat brain. However, low dose of aspartame had no significant effect on ionic concentration when compared to the control group. No significant difference in the level of Ca was observed in either of the aspartame-treated rats when compared to control.
Effect of Aspartame on Electrolyte Contents (µg/g Tissue).a
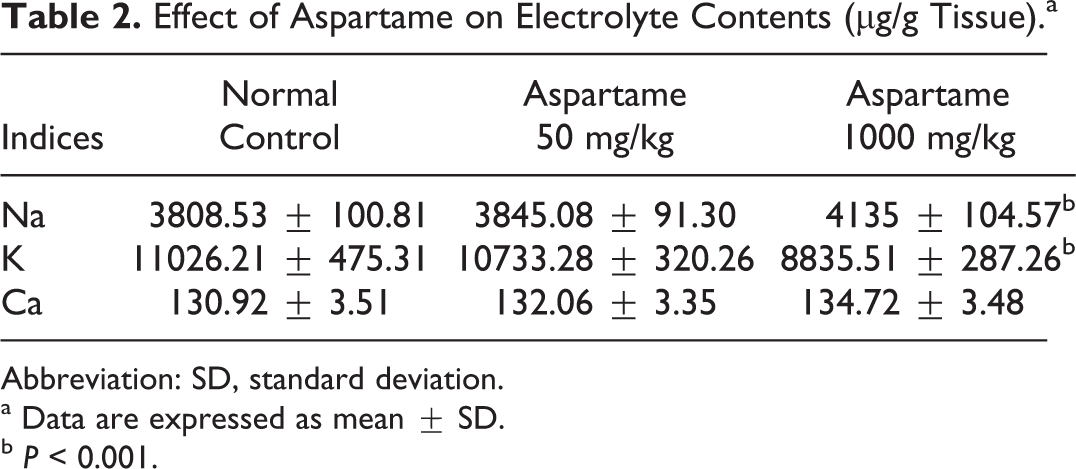
Abbreviation: SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.001.
Apoptosis Detection
The TUNEL assay was performed to detect apoptosis in the brain tissue sections. The TUNEL assay detects the genomic DNA fragments generated during apoptosis by labeling the free 3′-OH termini using terminal deoxynucleotidyl transferase enzyme. DNA fragmentation, as evidenced by fluorescein-12-dUTP incorporation, is an indicator of the occurrence of apoptosis. A significant (P < 0.001) percentage of TUNEL-positive (green fluorescence) cells was found throughout the brain section of rats treated with 1000 mg/kg aspartame compared to the control rats (Figure 1). However, only a few TUNEL positive cells were visible in the brain sections of low-dose group, suggesting that the apoptosis induced by aspartame may be dose dependent.

Aspartame-induced apoptosis in the brain cells of rats as detected by the terminal deoxynucleotidyl transferase mediated 2′-deoxyuridine, 5′-triphosphate nick-end labeling (TUNEL) assay. The representative sections are from corpus striatum. A, normal control (B) rats treated with aspartame 50 mg/kg showing no TUNEL positive cells (C) rats treated with aspartame 1000 mg/kg showing TUNEL positive cells (10×).
Amino Acid Levels
The concentrations of the following 5 neuroactive amino acids phenylalanine, tyrosine, tryptophan, aspartic acid, and glutamic acid were determined in the rat brain using HPLC. The effect of aspartame on these amino acids was shown in Table 3. Significant increase in phenylalanine levels was observed at both doses of aspartame. Administration of 1000 mg/kg of aspartame produces a significant increase in tyrosine (P < 0.05) and decrease in tryptophan level (P < 0.05) compared to control. But the low dosage of aspartame (50 mg/kg) produced no change in tyrosine and tryptophan concentrations. Table 3 further represents that no significant effect was observed in brain aspartic acid and glutamic acid in either of the aspartame doses.
Effect of Aspartame on Amino Acids (nmol/L/g tissue).a
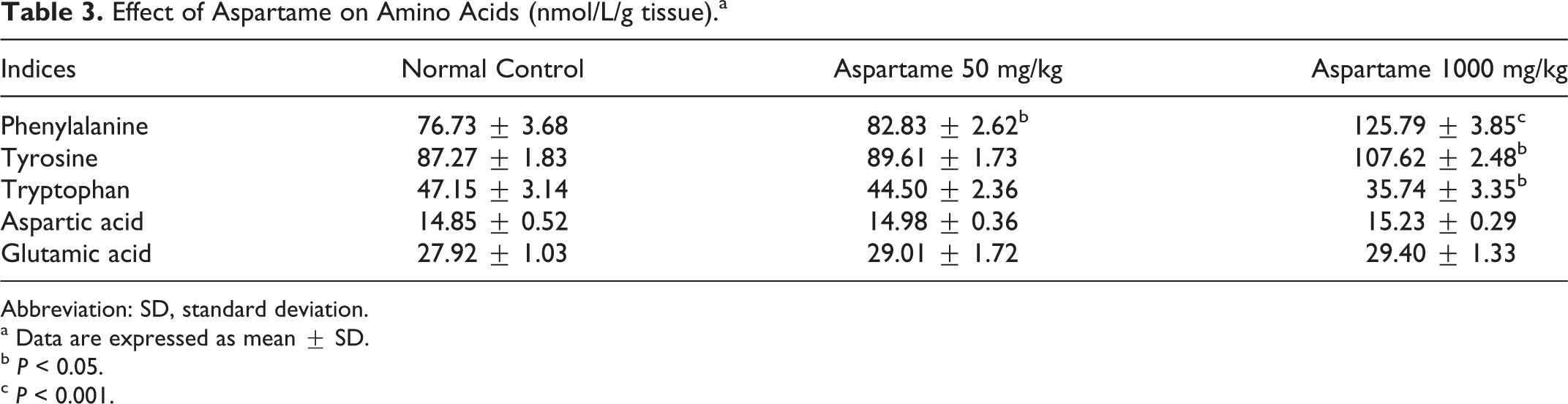
Abbreviation: SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.05.
c P < 0.001.
Tyrosine Hydroxylase and Tryptophan Hydroxylase
Tyrosine hydroxylase is the rate-limiting enzyme in catecholamine synthesis. It is the enzyme responsible for the conversion of the
Effect of Aspartame on Tyrosine Hydroxylase and Tryptophan Hydroxylase.a
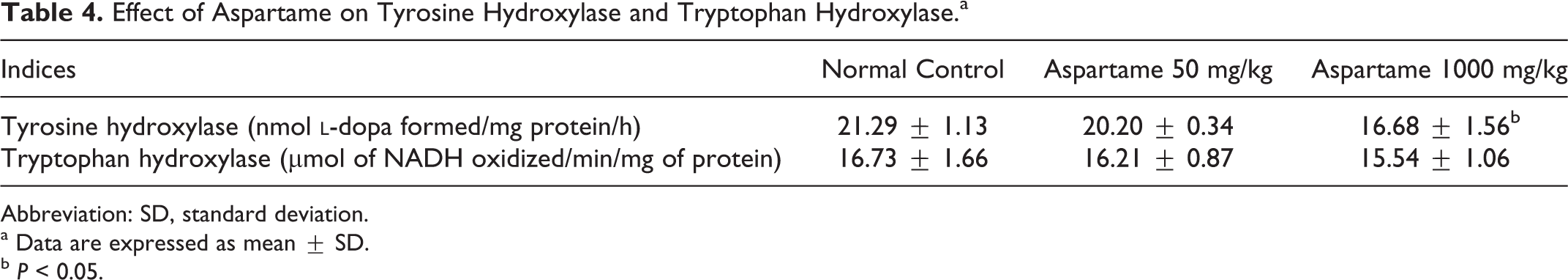
Abbreviation: SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.05.
Monoamine Neurotransmitter Levels
We analyzed the impact of aspartame administration on catecholamines and serotonin in tissue samples from corpus striatum and cerebral cortex. Chronic treatment of aspartame produced significant reduction in dopamine in the corpus striatum (Table 5) in both the experimental groups, whereas the cerebral cortex (Table 6) showed a significant reduction only at high dose of aspartame. Serotonin showed a significant decrease (P < 0.05) in the striatum (Table 5) at high dose of aspartame, while the low dose demonstrates no change. Norepinephrine and epinephrine were not affected by aspartame treatment.
Effect of Aspartame on Neurotransmitters Concentration in Corpus Striatum (ng/g tissue).a
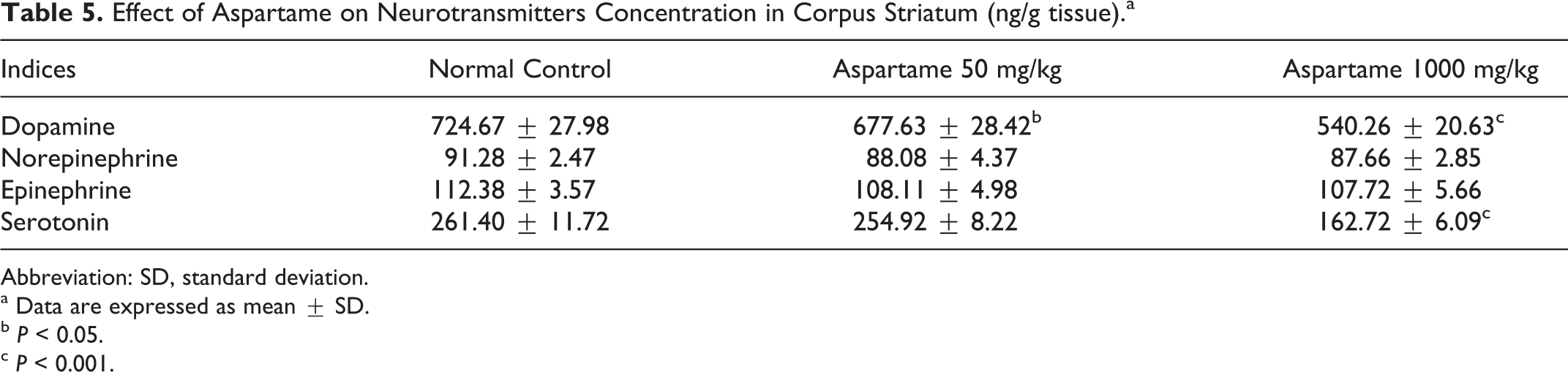
Abbreviation: SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.05.
c P < 0.001.
Effect of Aspartame on Neurotransmitters Concentration in Cerebral Cortex (ng/g tissue).a
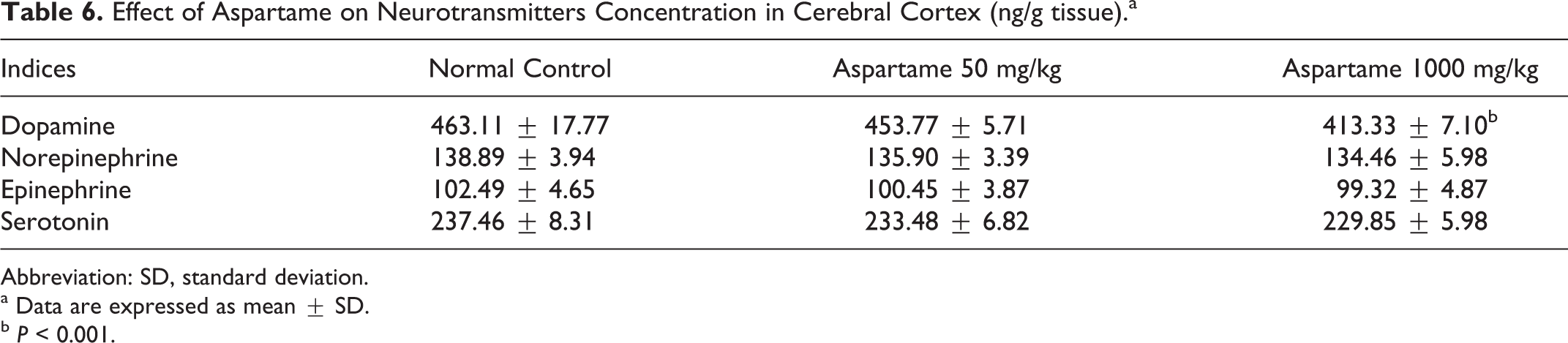
Abbreviation: SD, standard deviation.
a Data are expressed as mean ± SD.
b P < 0.001.
Discussion
A large number of the diet beverages and food products currently in the market use aspartame as a sweetener. There is an ongoing controversy over the decades surrounding the adverse effects of aspartame on the central nervous system. In the present study, we highlight the effect of long-term consumption of aspartame on ionic homeostasis and monoamine neurotransmitter synthesis in the rat brain. Our results showed that the activity of Na+-K+-ATPase was significantly reduced at the high dose of aspartame followed by a disturbance in electrolyte homeostasis and induction of apoptosis. There has been a debate over the potential of aspartame in producing alterations in regional monoamine synthesis. This study demonstrated that aspartame administration caused a dose-dependent variation in monoamine precursor amino acids and regional monoamine neurotransmitter concentrations. It is noteworthy that a decrease in the activity of tyrosine hydroxylase, the rate-limiting enzyme in catecholamine synthesis, has also been observed.
Na+-K+-ATPase is a membrane-bound enzyme responsible for the active transport of sodium and potassium ions in the nervous system, thus maintaining the ionic gradient necessary for neuronal excitability and regulation of neuronal cell volume. In the present study, we observed a significant reduction in Na+-K+-ATPase activity in rats administered with a dose of 1000 mg/kg body wt of aspartame. Our observation is in contrast with a previous study, which reported an increase in brain Na+-K+-ATPase activity after aspartame administration. 33 Growing body of evidence suggests that the Na+-K+-ATPase is susceptible to attack by reactive oxygen species (ROS) 34 and the SH groups of the enzyme are highly susceptible to oxidative stress. 35 The function of membrane proteins is modulated by the composition and fluidity of the lipid bilayer. Since Na+-K+-ATPase is a membrane-embedded protein, the nature of constituents comprising the membrane components should be an important determinant of enzyme function. It is known that the plasma membrane is a critical site for free radical attack. Recent studies suggest that aspartame produces oxidative stress and lipid peroxidation in brain. 36,37 Oxidative stress and lipid peroxidation change physiological functions of cell membranes by modifying the membrane bilayer, which is susceptible to free radical damage due to the presence of unsaturated fatty acids and the transmembrane proteins containing oxidizable amino acids. Consecutively, the properties of membrane-bound enzymes could be altered. It is suggested that phenylalanine, an important metabolite of aspartame, could induce reactive species production 13 and oxidative stress in experimental models. 38,39 Further, it has been found that high phenylalanine concentrations reduced Na+-K+-ATPase activity in rat brain. 40 Supporting this observation, we also observed a significant increase in phenylalanine in this study. Therefore, it is conceivable that reduction in Na+-K+-ATPase activity observed in this study may possibly result from free radicals elicited by aspartame metabolites, which attack the lipid bilayer followed by membrane disorganization and suppression of enzyme activity.
Establishing and maintaining high K+ and low Na+ in the cytoplasm are required for the maintenance of normal membrane potential and for various cellular activities. Our results showed that potassium content is significantly decreased and sodium content increased at high dose compared to control rats. The Na+-K+-ATPase uses about 40% to 50% of the ATP generated in the brain to pump Na+ out and K+ into the cell. 41 Since Na+-K+-ATPase is vital for maintaining ionic gradients in brains, it is accepted that a reduction in the activity of this enzyme may affect memory. Christian et al 10 have shown that chronic consumption of the aspartame may affect the T-maze cognitive performance of male rats, promoting impairment in the retention of learning behavior when compared to the performance of controls. It has previously been suggested that Na+-K+-ATPase inhibition can lead to memory impairment in the inhibitory avoidance and in the Water Maze tasks. 42 Recent studies 11,37 observed an impairment of spatial memory in the aspartame diet group when performed the Morris Water Maze test and suggest that exposure to aspartame may affect rodent spatial memory. Compelling evidence has indicated that modulation of Na+-K+-ATPase activity can affect learning and memory. 43 In this context it can be assumed that the decrease in Na+-K+-ATPase activity altered the Na+and K+ electrochemical gradient and disturbed the ionic homeostasis that subsequently cause impairment in spatial recognition memory observed in these studies.
It has been reported that failure of Na+-K+-ATPase, as a result of combination of energy deficiency and the production of ROS, is a common event in the apoptotic cascade. 44 Our results demonstrate that aspartame administration at high dose produces apoptosis as observed from TUNEL assay, which is an indicator of DNA fragmentation; a step in the apoptotic cascade. In recent years, considerable evidence has suggested that aspartame intake alters the neuronal cell integrity. 45,46 A large number of stimuli and signal pathways may contribute to the occurrence of apoptosis including oxidative imbalance. It is known that the brain may be particularly vulnerable to oxidative injury because of its high level of polyunsaturated fatty acids, high rate of oxygen consumption, and low or moderate levels of the antioxidant enzymes. 47 We reported a significant reduction in glutathione (GSH) and glutathione reductase activities in the brain after aspartame administration. 48 Recently, it has been demonstrated that aspartame administration produces oxidative stress in the brain. 36 Glutathione functions as a major antioxidant in tissue defense against oxidative stress, including the brain. It is suggested that glutathione loss is an early hallmark in the progression of cell death in response to different apoptotic stimuli. 49 Indeed, reports demonstrated that GSH depletion resulted in the impairment of Na+-K+-ATPase activity during apoptosis. 50 As Na+-K+-ATPase is a critical player in maintaining ionic homeostasis, impairment in Na+-K+-ATPase activity concomitantly reduces intracellular K+ and increases Na+. 51 Further, it is reported that apoptotic cells were characterized by a reduction in potassium content and an increase in sodium content. 52 Several studies have suggested that changes in ionic content, primarily K+, play a pivotal role in the progression of apoptosis. 53,54 It has been observed that apoptotic cells usually lose large amounts of intracellular K+. 55 Moreover, Franco et al 56 demonstrated that GSH loss regulates the progression of apoptosis by the modulation of K+ loss, which further regulates the execution phase of apoptosis. In addition to the changes in potassium content, the present study showed a higher concentration of sodium content. It was found that apoptotic cells are also characterized by a higher concentration of sodium content. 57 It is likely that impairment in Na+-K+-ATPase activity and related disturbances in electrolyte contents coupled with reduction in GSH may be responsible for the apoptosis observed in this study.
We observed a marked increase in phenylalanine and tyrosine and a decrease in tryptophan concentrations at high dose of aspartame in the brain, while the low-dose group showed significant increase in phenylalanine only. A similar increase in phenylalanine level was observed in previous studies also. 16,24 It is well known that the transport of phenylalanine from blood to brain is a dynamic process facilitated by the large neutral amino acids (LNAA) transporter type I (LAT 1) along with tyrosine, tryptophan, and other neutral amino acids. 58 The LAT 1 transporter has different affinities for each LNAA, and the binding in turn depends on their own plasma concentrations (LNAA). 59 Previous studies reported that aspartame significantly increased plasma phenylalanine levels. 60,61 Thus, elevated blood phenylalanine concentrations, which results from aspartame metabolism, are believed to increase the uptake of phenylalanine from blood to brain and reduce the uptake of other amino acids including tyrosine and tryptophan. Since phenylalanine is converted into tyrosine in liver catalyzed by phenylalanine hydroxylase, tyrosine level also increases. The increased concentrations of tyrosine observed in this study may be caused by this mechanism. Torii et al 62 also found a similar increase in tyrosine in the brain. It has previously been demonstrated that the elevated plasma phenylalanine concentrations will severely impair the transport of tryptophan into the brain. 63 It is feasible that the higher plasma phenylalanine resulting from aspartame consumption competitively decreases the tryptophan levels in the brain.
Phenylalanine plays a significant role in the synthesis and regulation of monoamine neurotransmitters. In this study, we observed regional variation in monoamine neurotransmitter levels, suggesting a specific aspartame-induced alteration in the monoamine biosynthesis. We found a significant decrease in the dopamine levels in the corpus striatum and cerebral cortex in both experimental groups after chronic aspartame administration. Our results are in consensus with a previous study, which reported a similar decrease in dopamine in striatum. The rate of synthesis and release of monoamine neurotransmitters are directly modified by the brain concentrations of their amino acid precursors, that is, phenylalanine, tyrosine, and tryptophan, which in turn are influenced by their availability from the blood. 64 Dopamine is synthesized from tyrosine, which is derived from phenylalanine. The initial step involves hydroxylation to dihydroxyphenylalanine, catalyzed by the enzyme tyrosine hydroxylase. Kaufman 65 has suggested that the rate of tyrosine hydroxylation is mainly determined by the tyrosine hydroxylase activity. In this study, we observed a marked reduction in tyrosine hydroxylase activity at high dose of aspartame. It is known that the major metabolite of aspartame, phenylalanine, at high concentration is a competitive inhibitor of tyrosine hydroxylase. 66 Reduced synthesis of dopamine may be caused by competition between brain phenylalanine and tyrosine for hydroxylation by tyrosine hydroxylase. 67 It is likely that the reduction in dopamine observed in this study may be caused by a reduction in the tyrosine hydroxylase activity induced by high concentration of phenylalanine. Dopamine plays a role in a wide range of cognitive tasks that involve manipulation and monitoring of information. Interestingly, dopamine seems to be an important neurotransmitter involved in learning and memory. 68 It is reported that dopamine depletion or blockade in cortex or striatum leads to impaired working memory performance. 69 So the implications of our observation should be investigated in detail to clarify the effects of aspartame in cognitive functions.
Serotonin plays a key role in many behavioral and neuropsychological processes including mood, reward, aggression, and memory. The present study found a significant decrease in serotonin in the corpus striatum at high dose of aspartame. The synthesis of serotonin occurs via hydroxylation of tryptophan to 5-hydroxytryptophan catalyzed by tryptophan hydroxylase. Subsequently 5-hydroxytryptophan is converted into serotonin. The rate of serotonin synthesis is tied to local concentrations of its amino acid precursor tryptophan. 70 So an increase in phenylalanine level competes with tryptophan for transport across the blood–brain barrier as has been observed in this study. Another mechanism by which serotonin synthesis is regulated is through the activity of tryptophan hydroxylase. But in this study, tryptophan hydroxylase activity was found to be unaltered. In this context, it is plausible that the reduction of serotonin in striatum may be caused by the decrease in brain tryptophan as a result of the competitive inhibition caused by elevated phenylalanine. The findings of our study contradict a previous study in which an acute administration of aspartame increases serotonin levels in the striatum and reduces aggressive behavior in rats. Serotonergic dysfunction has been reliably associated with aggressive behaviors in animals and humans. 71 However, it is not clear about the implications of our findings on aggression.
Conclusion
In conclusion, the results of this study demonstrate that long-term consumption of aspartame produces disturbances in ionic homeostasis, regional monoamine concentrations, and induced apoptosis in brain. However, changes were prominent at high dose, while the ADI dose elicited limited effects. The data obtained in the present study are relevant since there is increasing body of evidence related to the cognitive effects of aspartame. Further research must be performed to clarify these observations on cognitive functions and elucidation of the mechanisms.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.