
Abstract
The feasibility and relevance of screening a library of raw actinomycete extracts (ECUM library) for the identification of antituberculosis activities was assessed on 11,088 extracts using a multiple-screening approach. Each extract was first tested at two concentrations against noninfected macrophages as a control, then against Mycobacterium tuberculosis growing in broth medium as well as infecting murine macrophages. The screening results indicated a library of good quality with an apparent low proportion of cytotoxic extracts. A correlation was found between both bacterial assays, but the intracellular assay showed limitations due to low rates of cell survival. Several extracts of interest were highlighted by this multiple screening. A focus on the strain producing the two most effective revealed similarities with known producers of active molecules, suggesting the possibility of selecting relevant extracts using this strategy.
Introduction
Tuberculosis (TB) is an infectious disease caused by Mycobacterium tuberculosis (Mtb) and commonly affecting the lungs. Despite the existence of a vaccine, diagnostic tools, and an effective chemotherapy, TB still remains one of the deadliest infectious diseases in the world, with 1.5 million deaths and 9 million new infections reported in 2014. 1 Thanks to the efforts deployed by pharmaceutical companies and academics during the past 15 years, new chemical entities have started to be available for the treatment of multidrug-resistant (MDR) and extensively drug-resistant (XDR) strains—namely, bedaquiline 2 and delamanid. 3 Both drugs have received conditional regulatory approval for the treatment of MDR infections. However, the high capacity of Mtb to escape drug treatment by acquiring genomic mutations urges us to use these drugs with the utmost precaution. The recent notification of clinical isolates already displaying resistance to bedaquiline 4 or both bedaquiline and delamanid 5 is particularly concerning and prompts us to continue our search for new effective drugs. In this context, the characterization of yet unexplored sources of chemical entities is of primary interest.
Nature is a source of unique chemical structures and diversity, and has been a source of medicinal products for thousands of years. 6 Since the 1940s, actinomycetes’ secondary metabolites, as well as their associated derivatives, have played a major role in the development of anti-TB therapy. This is best exemplified by streptomycin, an extremely potent aminoglycoside targeting protein synthesis in Mtb. Streptomycin was the first clinical drug available against TB7,8 and is still widely used nowadays for the treatment of drug-resistant TB infections. Similarly, rifampicin was obtained by chemical modifications of rifamycin B, a natural product of the rifamycin family. 9 Rifampicin is one of the most important drugs of first-line therapy against TB, along with isoniazid. Its sterilizing ability makes it an essential component, as it helps to reduce the treatment duration and the relapse rate.10,11 Both streptomycin and rifampicin are part of the list of essential medicines as defined by the World Health Organization (WHO), 12 highlighting their value and importance as antibiotics. In addition, many other natural products or derivatives displaying growth inhibition against Mtb are regularly reported in the literature, 13 proving the value of natural products for the field of anti-TB drugs.
More natural products are yet to be discovered, and the entry into the genomic era has led to a new wave of interest in their use for drug discovery.
14
In South Korea, the Center for Nutraceutical and Pharmaceutical Material (CNPM, www.cnpm.re.kr) in Myongji University collected microorganisms from various geographical regions displaying unusual or extreme climate, with the aim to isolate distinct actinomycete species. Isolated microorganisms with actinomycete-like morphologies were cultivated in the rich, glucose-soybean-starch (GSS;
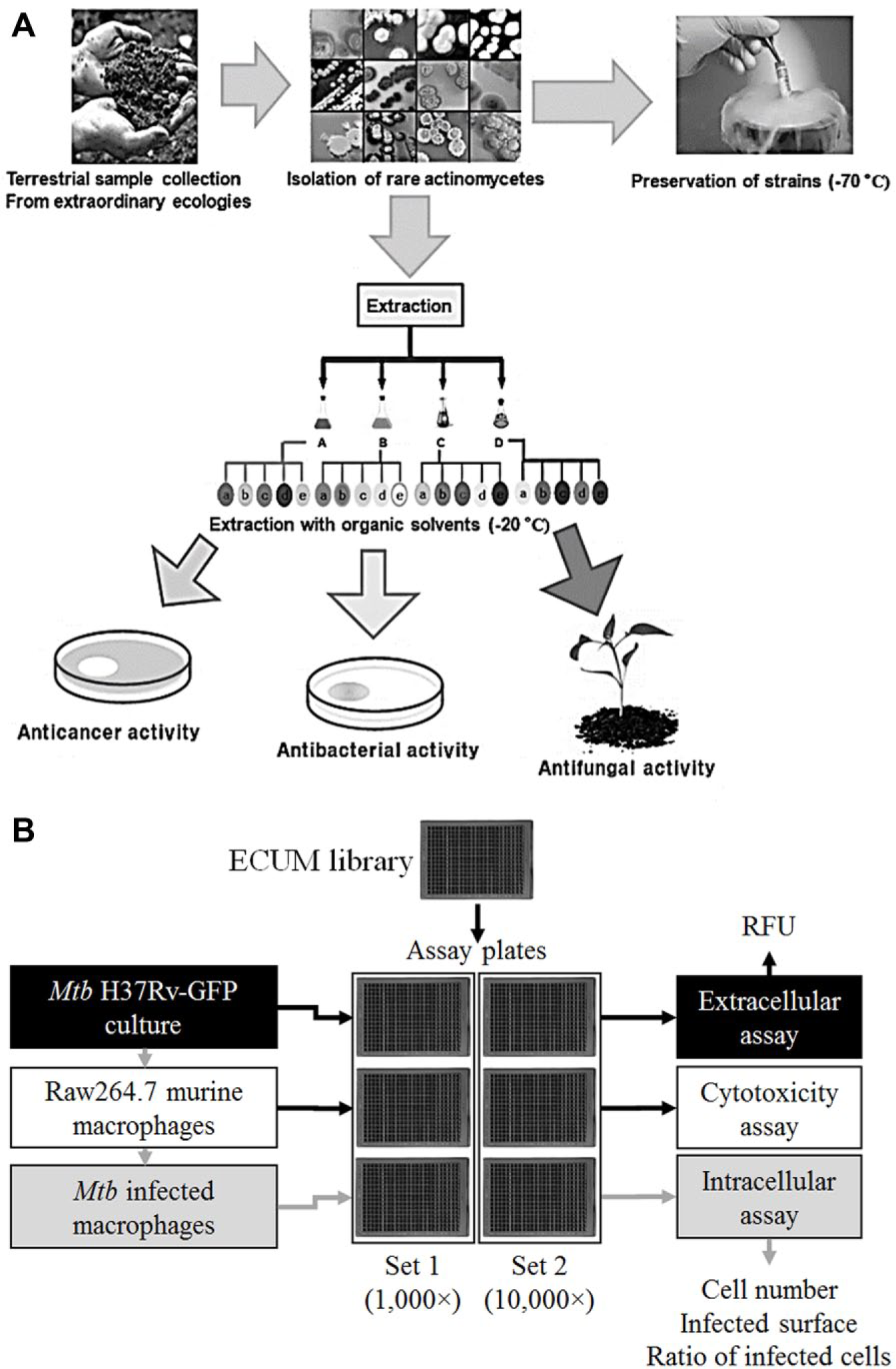
(
Materials and Methods
Reagents
DMSO, rifampicin and methanol were from Sigma-Aldrich (Yongin, South Korea). Rifampicin stock solutions were prepared in DMSO at 100 µg/mL and stored at −20 °C. Mycobacteria were cultured in 7H9 broth (Difco; Middlebrook, BD Biosciences, Seoul, South Korea) supplemented with 0.05% Tween 80 (Sigma-Aldrich), 0.2% glycerol (Invitrogen, Thermo Fisher Scientific, Seoul, South Korea), 10% OADC (Middlebrook), and 50 µg/mL hygromycin B (Gibco, Thermo Fisher Scientific, Seoul, South Korea), referred to as 7H9-Tw-Gly medium. Raw264.7 murine macrophages were cultured in RPMI 1640 medium without phenol red (Welgene, Gyeongsan, South Korea, South Korea), supplemented with 10% fetal bovine serum (FBS; Gibco), and referred to as RPMI-FBS medium. Phosphate-buffered saline (PBS), free of calcium and magnesium, was from Welgene.
Extract Preparation
After fermentation of each actinomycete strain, the supernatant and cell pellet were separated by centrifugation at 13,000 rpm for 20 min. One volume of ethyl acetate was added to one volume of supernatant and the mixture vigorously mixed for 20 min. After decantation, the ethyl acetate and water layers were separated and dried under vacuum to afford the crude ethyl acetate and water extracts, respectively. For the cell pellet, metabolites were extracted by addition of methanol (1:1, w/w) and incubation at room temperature for 12 h with mild shaking. The methanol phase was separated from the cell pellet by centrifugation at 13,000 rpm for 20 min and dried under vacuum to afford the methanol extract.
Assay Plate Preparation
Dried extracts (330 µg for water and methanol extractions; 20 µg for ethyl acetate extraction) were delivered in 384-well polypropylene plates (Nunc, Thermo Fisher Scientific, Seoul, South Korea) and resuspended in 20 µL methanol. Sealed plates were left overnight at room temperature with mild shaking to ensure complete solubilization. DMSO (80 µL) was added to generate the mother plates. Five sets of daughter plates were prepared by transferring 15 µL of the mother plate into 15 µL of DMSO. All plates were sealed and stored at −20 °C until further use. On the day of the screening, a full set of daughter plates was thawed at room temperature, and 2 µL was transferred into 18 µL of PBS to generate the intermediate plates. Assay plates were prepared by transferring 5 µL (1000×) or 0.5 µL (10,000×) of the intermediate plate into sterile 384-well plates adapted for confocal microscopy (781091; Greiner Bio-One, Han Sung Sci, Anyang, South Korea). For positive and negative controls, 0.5 µL of stock solutions (100 µg/mL rifampicin and 100% DMSO, respectively) was dispensed in the assay plates. In each plate, 16 wells were dedicated to the positive control and 16 for the negative control. Where required, 4.5 µL PBS was added prior to extract dispensing (control wells and 10,000× dilution).
Extracellular Assay
Mycobacterium tuberculosis strain H37Rv (ATCC 27294) was transformed with the Ms6-based integrative plasmid pNIP48, harboring a green fluorescent protein (GFP) gene from Aequorea victoria under the control of the pBlaF promoter from Mycobacterium fortuitum. 15 The plasmid confers resistance to hygromycin. The resulting strain, Mtb H37Rv-GFP, was propagated at 37 °C in 7H9-Tw-gly medium, washed with PBS, aliquoted, and stored at −80 °C. To obtain dispersed cultures for screening, aliquots were first thawed in 50 mL 7H9-Tw-gly medium and grown for 1 week at 37 °C, 5% CO2. Cultures were further diluted with fresh medium to an OD600 nm value of 0.1 before incubation for 1 more week at 37 °C, 5% CO2. Bacteria were harvested at early log-phase by centrifugation at 3000 × g for 10 min, washed twice with PBS, and resuspended in fresh 7H9-Tw-gly to reach an OD600 nm value of 0.01 (106 bacteria/mL). The quality of the bacterial suspension was assessed microscopically before the assay to ensure the absence of clumps. The bacterial suspension was dispensed in assay plates (45 µL/well) using an automated dispenser (Matrix WellMate; Thermo Scientific, Seoul, South Korea). Plates were incubated for 5 days at 37 °C, 5% CO2 before reading of the fluorescence (excitation, 490 nm; emission, 535 nm) using a microplate reader (Victor 3; PerkinElmer, Seoul, South Korea).
Cytotoxicity and Intracellular Assays
Mtb H37Rv-GFP strain was grown at 37 °C in 7H9-Tw-gly medium as dispersed cultures, harvested at early log-phase by centrifugation at 3000 × g for 10 min, washed twice with PBS, and resuspended in fresh RPMI-FBS to reach an OD600 nm value of 0.02 (2.106 bacteria/mL). In parallel, Raw264.7 macrophages were grown at 37 °C, 5% CO2 in RPMI-FBS medium up to 70% confluence, detached by mechanical scrapping and collected by centrifugation. The cell pellet was resuspended in fresh RPMI-FBS medium, with the cell concentration determined using a Thoma cell-counting chamber and the concentration of the cell suspension adjusted to 106 cells/mL. One volume of cell suspension was mixed with one volume of bacterial suspension to reach a multiplicity of infection (MOI) of 2, and the mixture was incubated in Erlenmeyer flasks for 2 h at 37 °C under mild shaking (100 rpm). Infected cells were pelleted, washed with RPMI-FBS, and finally resuspended in RPMI-FBS before dispensing in the assay plate (45 µL/well) using an automated dispenser (Matrix WellMate; Thermo Scientific). For the cytotoxicity assay, using noninfected macrophages, Raw264.7 cells were prepared as described above and the suspension adjusted at 5 × 105 cells/mL in RPMI-FBS prior to dispensing (45 µL/well). Plates were incubated for 5 days at 37 °C, 5% CO2. At the end of the incubation period, cell nuclei were stained using Hoechst 33342 at a final concentration of 2.5 µg/mL for 30 min at 37 °C, 5% CO2. Plates were sealed and imaged using an automated fluorescence microscope (Operetta; PerkinElmer) equipped with a 10× objective (0.4 NA) and configured for reading in the blue (excitation, 405 nm; emission, 450 nm) and the green (excitation, 488 nm; emission, 520 nm) channels. Three fields were imaged per well, and pictures were analyzed using an in-house software that counted the number of nuclei in the well using the blue channel and determining the position and area of the bacteria using the green channel. The number of cells was determined based on the nuclei population, segmented using an in-house seed growth algorithm. 16 In the case of the cytotoxicity assay, an additional plate without extract was also prepared, centrifuged, and imaged on day 0 to obtain the average number of cells present in the well after plating. Bacteria were detected using a classification method based on a support vector machine algorithm, allowing precise segmentation of the pixels positive for a GFP signal versus background signals. 17 A cell was considered infected if at least two contiguous pixels of bacteria were found within their cytoplasm. The ratio of infected cells was calculated by dividing the number of infected cells by the total number of cells. Data obtained for each field were averaged to give the value per well.
Data Analysis and Normalization
Values obtained for the positive and negative controls were used for data normalization and assay quality. Values obtained from the 66 plates of the same assay were considered to calculate
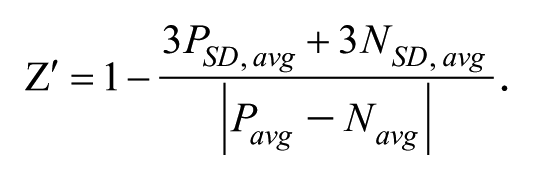
A Z′ value above 0.5 was considered representative of a very robust assay, and between 0.2 and 0.5 was considered a robust assay. For the extracellular assay, the percentage of growth inhibition (or percentage of inhibition) was calculated using the RFU value as follows:
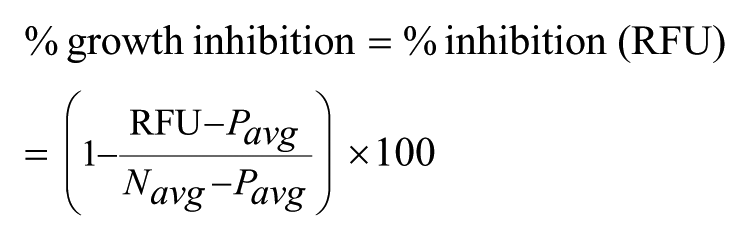
For the cytotoxicity assay, the percentage of viability was calculated as follows, using C, the number of cells in the well, and Pavg, the average number of cells found in the control wells (1% DMSO):
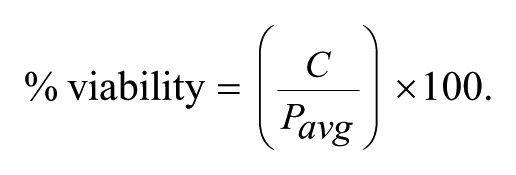
An extract was considered cytotoxic if the number of cells counted after 5 days was lower than the average number of cells counted after plating at day 0, which corresponded to 40% cell survival in our conditions. Extracts presenting 40% to 80% cell survival where considered either cytostatic or partially cytotoxic. For the infected macrophage assays, the percentage of cell survival was determined using the cell number
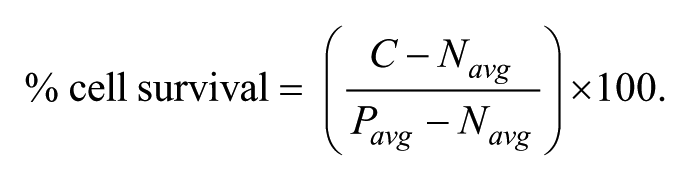
In this case, the percentage of cell survival can be a negative number, as the number of cells determined in the well in the presence of extract (C) can be lower than the number of cells found in the negative control (DMSO-treated infected cells, Navg). This is particularly true in the case of cytotoxic/cytostatic extracts. The percentage of infected surface was determined using the infected surface S (expressed in number of pixels) as follows:
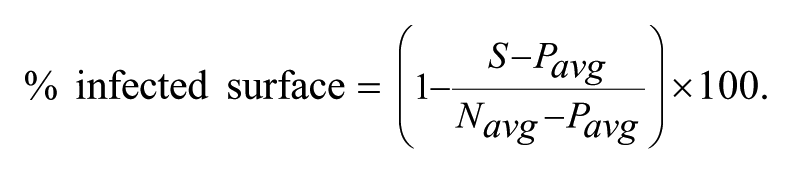
Finally, the percentage of inhibition, calculated based on the percentage of infected surface or the ratio of infected cells, was obtained using the following formulas:


Normalization, visualization, and extract selection were performed using the Spotfire software (Tibco, Palo Alto, CA). Relative frequency distributions and curve fitting were performed using the Prism software (GraphPad Software, La Jolla, CA). Statistical differences were calculated by Prism software as two-tailed p values.
DNA Purification and 16S Ribosomal RNA Sequencing
Selected microorganisms’ genomic DNA was extracted with the CoreBio genomic DNA isolation kit (CoreBio, Seoul, South Korea), following the recommended protocol. The 16S ribosomal DNA (rDNA) was amplified and sequenced using the universal primers: 27F (forward), 5′-AGAGTTTGATCATGGCTCAG-3′, and 1492R (reverse), 5′-AAGGAGGTGATCCARCCGCA-3′. The sequence was aligned with the closest Streptomyces strains in BLAST using CLUSTAL X, 19 and the phylogenic tree was constructed by the neighbor-joining method, using MEGA6.0 software (MEGA, Tempe, AZ). 20 The topology of the phylogenic tree was calculated by 1000 bootstrap replications. 21
Results
Extract Reformatting
A subset of 11,088 extracts was taken from the complete ECUM library. They originated from 1344 distinct strains, chosen according to their morphological and physiological resemblance to actinomycete strains. To increase the variety of secondary metabolites obtained from each strain, different culture media were used. Three-fourths of the selected strains (1008 strains, 75%) were grown in three different media (GSS, rich medium; BN, Bennett’s regular medium; DYC, poor medium; see
Cytotoxicity Assay
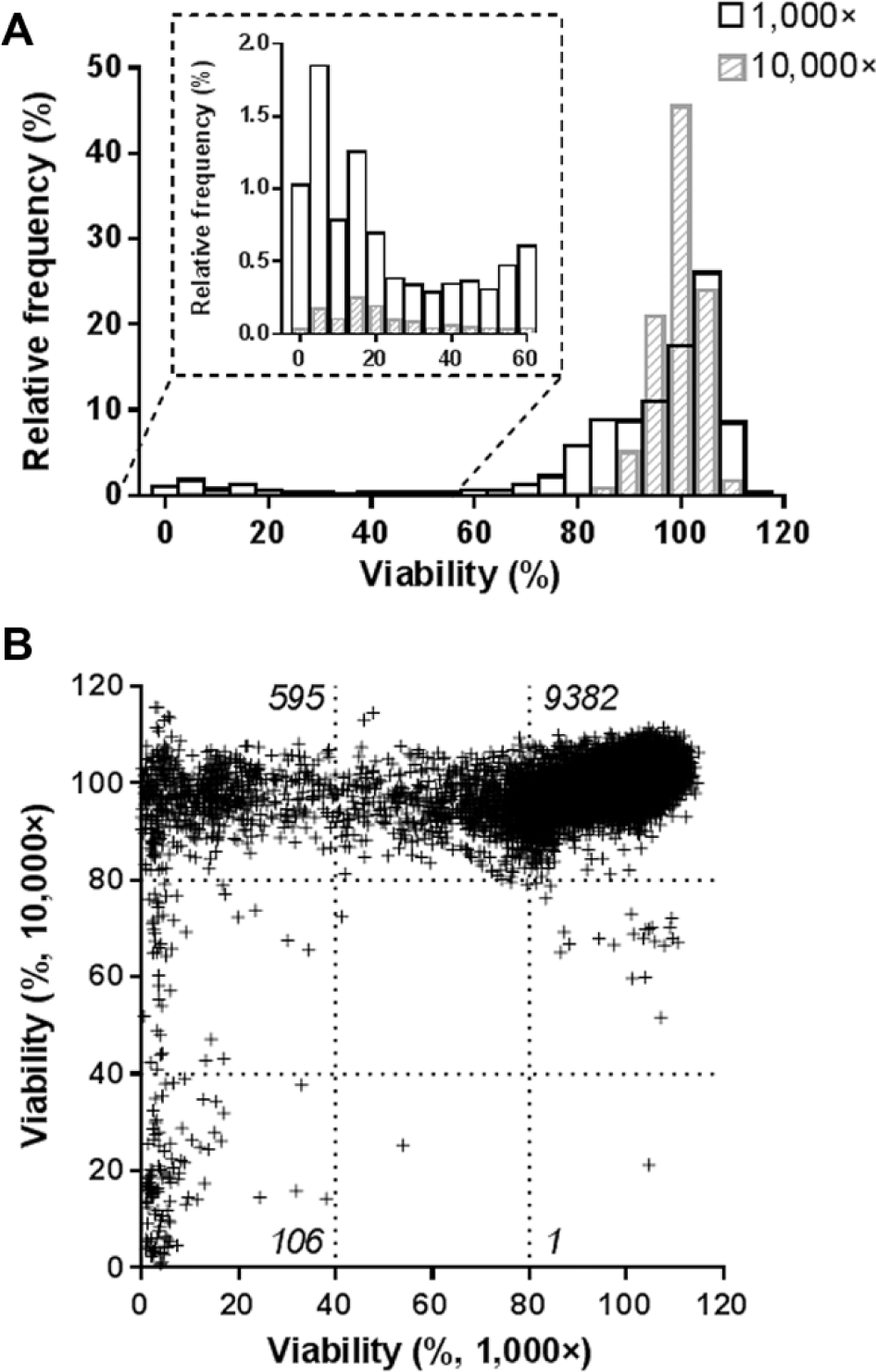
Each extract was first tested at both concentrations against noninfected macrophages to identify and quantify the extent of cytotoxic effects within this subset of the library. As shown in Figure 2A , most of the extracts were not cytotoxic or cytostatic for the cells. Indeed, at the 10,000× and 1000× dilution, 98.4% and 84.7% of the extracts displayed above 80% of viability, respectively. As expected, when the extracts were used at higher concentrations (1000×), more cytotoxicity was observed, as indicated by the increased proportion of extracts presenting less than 40% of viability ( Fig. 2A , insert). Correlating the results obtained at both concentrations indicated that most of the extracts were not cytotoxic at both dilutions ( Fig. 2B ). A small proportion was cytotoxic at both dilutions (1.0%) or only at the 1000× dilution (5.4%). The number of extracts with toxicity at only the 10,000× dilution was negligible (1 of 11,088). Counter-checking the pictures taken from the well, some extracts appeared to have a high fluorescence background, but this was without an impact on the detection of the cells due to their much higher fluorescence intensity. Altogether, these results showed that a low proportion of extracts was altering the cell viability, suggesting that this subset of the library was suitable to carry out a phenotypic assay using infected host macrophages.
(
Extracellular Assay
The ability of the extracts to prevent Mtb replication in broth medium was assessed by monitoring the fluorescence (RFU) remaining in the well after 5 days of incubation in the presence of the extracts. For each set, 1000× and 10,000×, we confirmed that the distribution frequency of RFU values followed a Gaussian distribution (R2 > 0.99), centered on 10,300 and 10,600, respectively (
Fig. 3A
). DMSO (1%) and rifampicin (1 µg/mL) were used as negative and positive controls, respectively. The average Z′ value for the whole extracellular screening was 0.72 (66 plates; min. Z′: 0.41; max. Z′: 0.85;
(
Extracts Displaying More Than 50% Growth Inhibition at Both 1000× and 10,000× Dilutions for the Extracellular Assay.
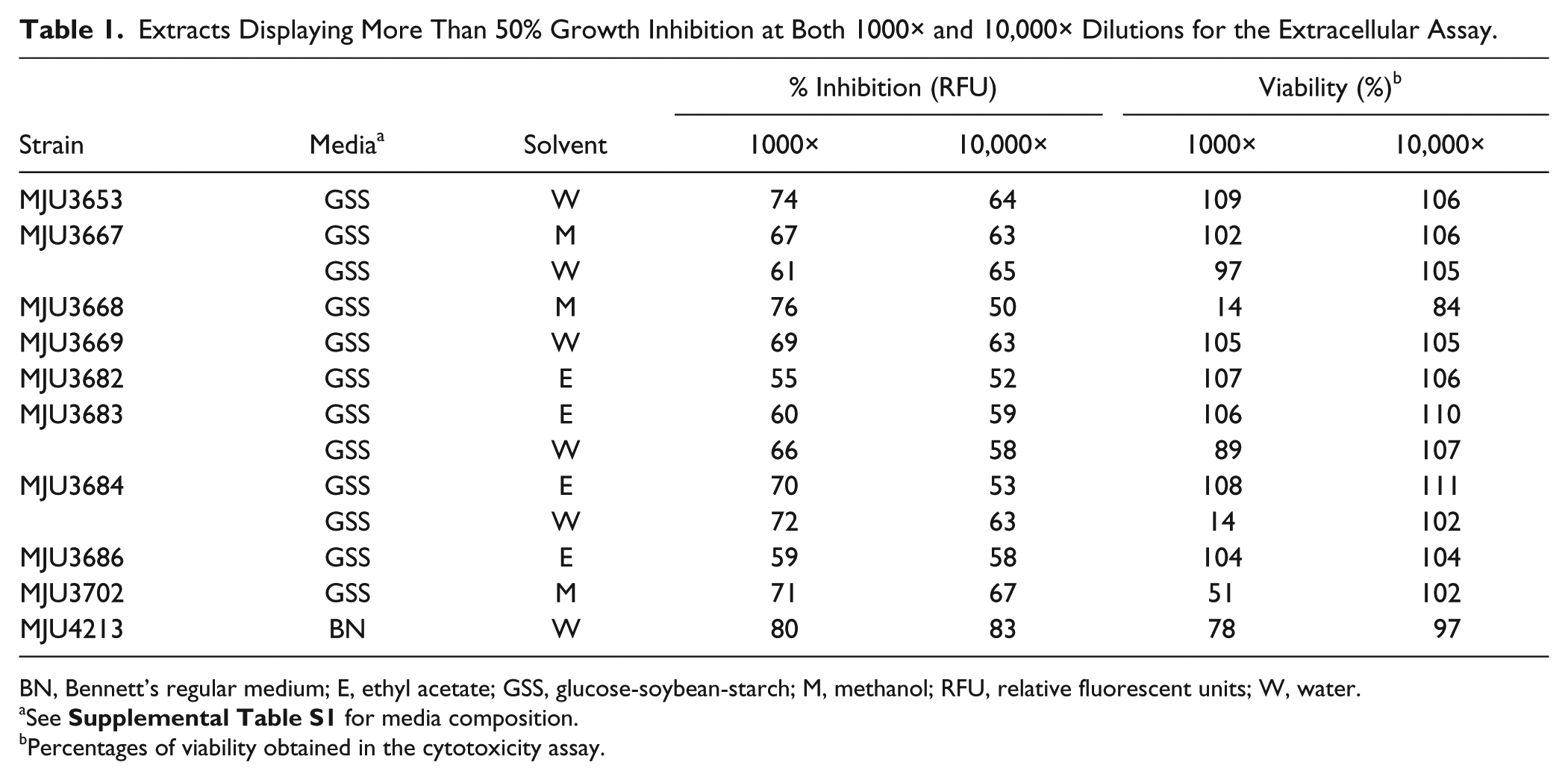
BN, Bennett’s regular medium; E, ethyl acetate; GSS, glucose-soybean-starch; M, methanol; RFU, relative fluorescent units; W, water.
See
Percentages of viability obtained in the cytotoxicity assay.
Intracellular Assay
The presence of extracts displaying an ability to inhibit bacterial replication in host macrophages was also investigated within this subset of the ECUM library. Indeed, Mtb is known to display a metabolic switch while infecting macrophages, leading to different susceptibilities toward antibiotic treatments.24,25 It was then interesting to assess, in parallel to the extracellular assay, whether some extracts would be selected as well in macrophages or be specific for one assay or the other. A robust and already validated high-content screening (HCS) approach was used (see Materials and Methods section and
Fig. 1B
).15,26 The infection ratio parameter, as it takes into account both the cellular and bacterial populations, was used to determine the assay quality. The frequency distribution for the 1000× dilution followed a normal distribution (R2 > 0.99). In the case of the 10,000× dilution, however, a high number of extracts displayed a low to moderate activity on the ratio of infected cells, and the distribution was better fitted by a sum of two Gaussians (R2 > 0.99) rather than a single Gaussian (R2 = 0.90) (
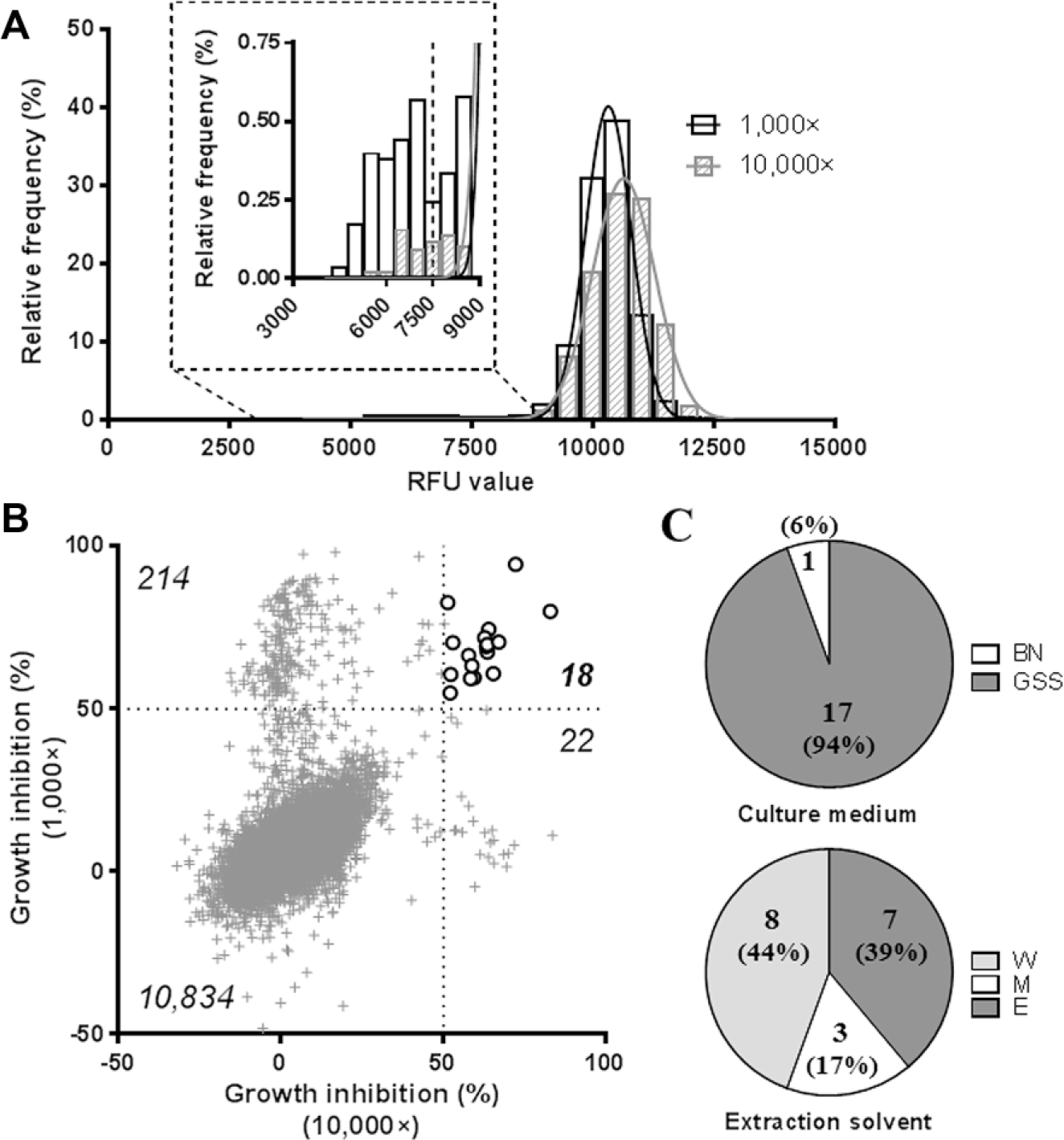
This large amount of extracts displaying a small reduction of infection ratio at 10,000× led us to investigate the cell number and the infected surface individually to better understand the fate of the cell and the bacteria. Each parameter was normalized based on the positive and negative controls (1 µg/mL rifampicin and 1% DMSO, respectively), and Gaussian distributions were confirmed, with R2 > 0.98 ( Fig. 4A , B ). The high frequency of low cell survival was striking, with more than 90% of the extracts displaying less than 10% cell survival for both dilutions ( Fig. 4A ). This result contrasted significantly with the near absence of cytotoxicity observed on noninfected macrophages ( Fig. 2A ). The low rates of cell survival observed at both dilutions in the intracellular assay may be due to cytostaticity, without visible consequences in the cytotoxicity assay but detrimental in the infected macrophage assay due to its higher stringency. Considering the low number of cells remaining in the well, we reasoned that the infection ratio would not be appropriate as a readout of the extract potency and decided to use the cell survival and the infected surface. Of importance, the infected surface, quantifying the area of bacteria in the well, was well adapted to quantify the effect of the extract on the bacterial replication. When these two parameters were plotted together, a subpopulation of extracts displaying more than 40% of cell survival and less than 50% of infected surface was revealed for each dilution ( Fig. 4C ). A total of 88 extracts (0.8%) were selectable using these criteria (50 were selectable at 1000×, 29 at 10,000×, and 9 at both dilutions). However, counter-checking the pictures taken from the corresponding wells revealed that all of these extracts were actually artifacts with high fluorescence background, either due to the extract itself (confirmed with the cytotoxicity assay) or a technical issue during the acquisition ( Fig. 4D ). This result was quite disappointing, but it is worth noting here that, from all the extracts classified as artifacts, none belonged to the list of extracts selected from the extracellular assay.
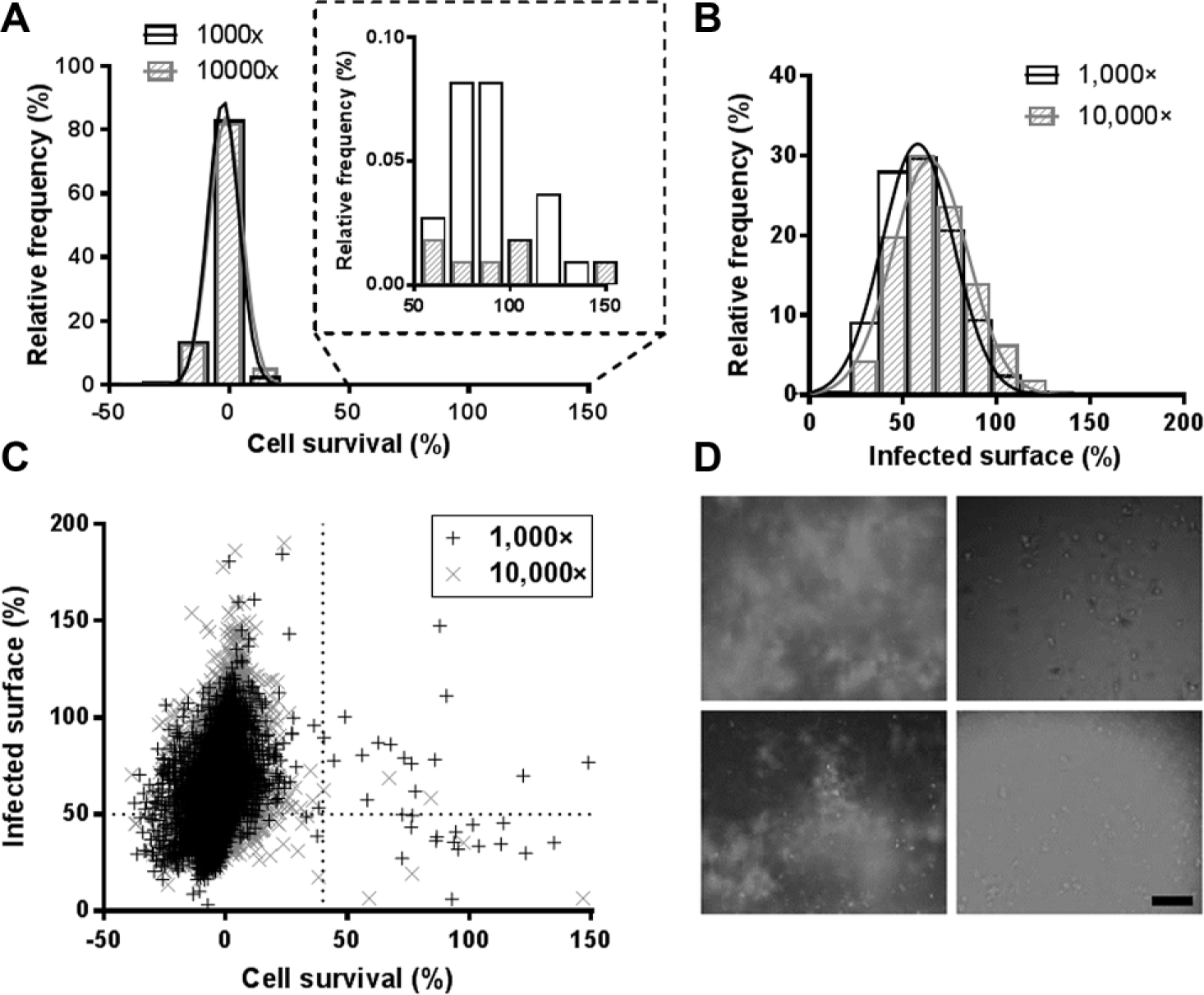
Relative frequency distributions for the intracellular screening. (
Correlation between Both Assays
Despite the lack of potent extracts obtained using these criteria from the intracellular assay, we were interested in investigating how the results were correlated between both bacterial assays. Percentages of inhibition were compared in the case of the 18 extracts selected through the extracellular assay, before removal of the cytotoxic extracts, to keep more information (
In light of these results and with the wish to extract more data from the intracellular screening, we decided to reevaluate the presence of active extracts focusing only on the infected surface parameter, without consideration for cell survival. By doing so, we drastically increased the number of selectable extracts at 50% inhibition (4718) and thus had to determine a better threshold for the selection of relevant extracts. Using the distribution frequency of the percentage of inhibition, we estimated that a threshold above 70% would allow the selection of relevant extracts with a limited rate of false positives, as this corresponds to more than 1.5 times the standard deviation of the distribution for both dilutions (
Extracts Displaying More Than 70% Inhibition of Infected Surface at Both 1000× and 10,000× Dilutions for the Intracellular Assay.
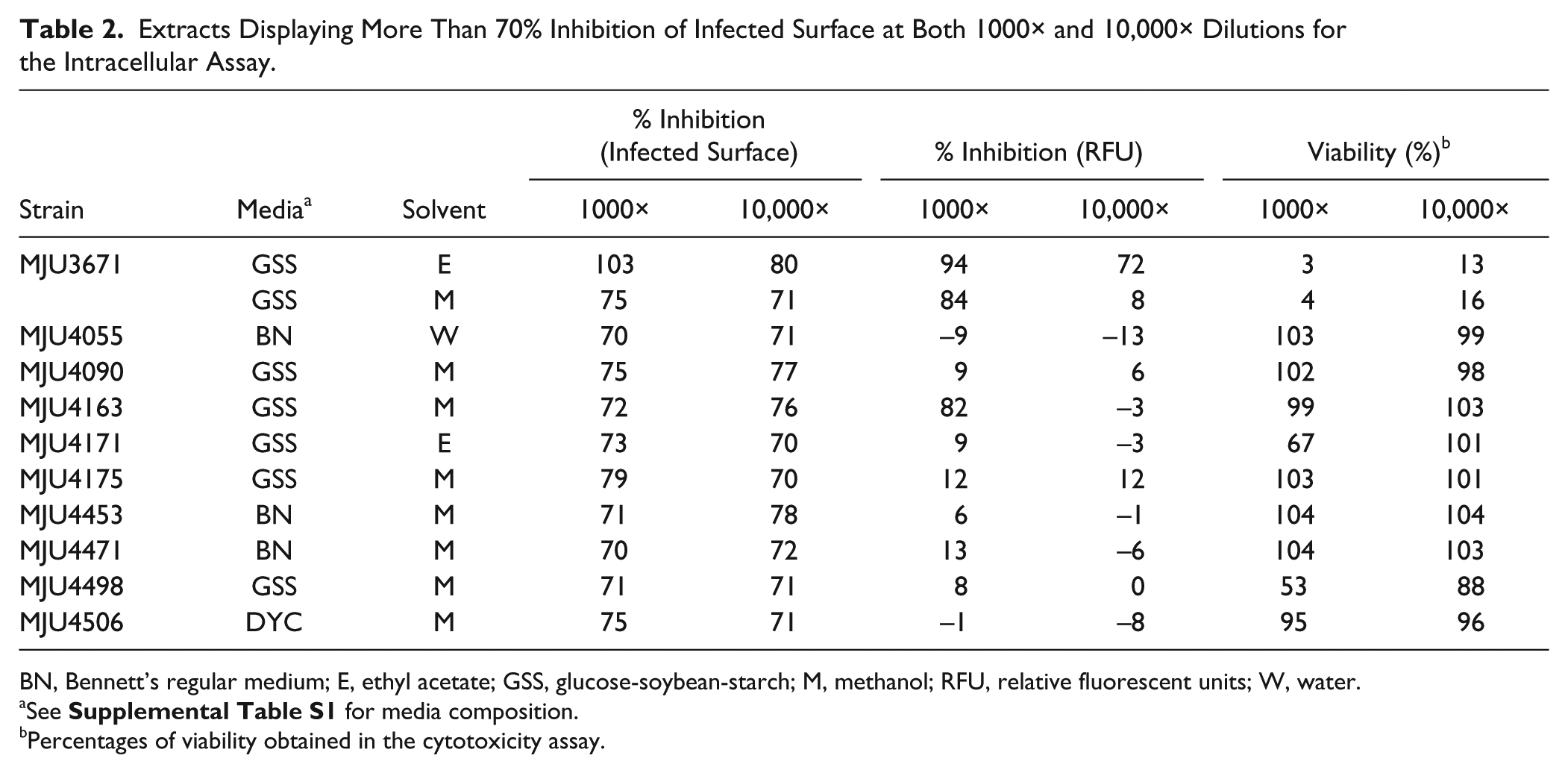
BN, Bennett’s regular medium; E, ethyl acetate; GSS, glucose-soybean-starch; M, methanol; RFU, relative fluorescent units; W, water.
See
Percentages of viability obtained in the cytotoxicity assay.
A Focus on the Strain MJU3671
This strain was isolated from a soil sample taken in Ham-Bak-Mountain, South Korea. The 16S rRNA gene of this strain was sequenced, yielding a string of 1426 bp with high similarities for the Streptomyces species. The sequence presented 99% identity with Streptomyces costaricanicus (NBRC 10077), Streptomyces murinus (NBRC 14802) and Streptomyces misionensis (NBRC 13063), and Streptomyces phaeogriseichromatogenes (NRRL 2834T), its closest neighbor within the phylogenetic tree (
Discussion
Here, we report the screening of 11,088 natural extracts obtained by solvent extraction of the mycelium and supernatant of actinomycete-like cultures. These extracts were tested as a representative subset of the ECUM library to probe the technical feasibility of a multiple screening against this library. Each extract was screened at two different dilutions, using three distinct assays: noninfected macrophages, bacteria replicating in broth medium, and bacteria replicating in macrophages. At the end of the screening campaign, a total of 24 extracts were selected ( Table 1 and Table 2 ), but only one extract was found active in both bacterial assays, while displaying a high toxicity against the noninfected macrophages. This raw ethanolic extract was obtained from a strain of the Streptomyces genus, highly similar to known antibiotic producers. The results obtained here aid us in the capacity of our multiple-screening assay to unravel anti-TB activity within complex natural extracts. Of particular interest here was the very low rates of cell survival observed in the intracellular assay, despite an apparent absence of dramatic viability loss against noninfected cells. While this needs to be investigated further, the higher stringency of this assay may be the reason for this difference. Indeed, an inhibition of the macrophage replication during the first days of incubation, through the presence of antitumor and antiproliferating agents in these extracts, may have only a little impact when using noninfected cells but could be detrimental in the case of infected macrophages, as they rapidly die and release viable bacteria in the supernatant. The use of a nonreplicating cell line, like THP-1 macrophages or non-tumor-derived primary cells, may be a way to palliate this problem and increase the chances to identify valuable fractions from the full library, with the counterparts of increasing the overall cost of the screening. Accordingly to these findings, the protocol of the intracellular assay for the screening of the entire ECUM library (~150,000 extracts) will be revised after evaluating the performance of the assay using different macrophage cells. In addition, the other strains highlighted during this prescreening will be further evaluated to determine their interest in regard to the development of drugs or the identification of pathways of interest for the fight against TB.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Institut Pasteur Korea is a member of the Institut Pasteur International Network (![]() ). This work was supported by the Korean Ministry of Science and International Planning (MSIP, project number NRF-2014K1A4A7A01074643), as well as the Cooperative Research Program for Agriculture Science and Technology Development, Rural Development Administration, Republic of Korea (project number PJ01128901) and the Gyeonggi province, Korea. VD was supported by the French Ministry of Foreign Affairs.
). This work was supported by the Korean Ministry of Science and International Planning (MSIP, project number NRF-2014K1A4A7A01074643), as well as the Cooperative Research Program for Agriculture Science and Technology Development, Rural Development Administration, Republic of Korea (project number PJ01128901) and the Gyeonggi province, Korea. VD was supported by the French Ministry of Foreign Affairs.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.