
Abstract
Embryo implantation requires a healthy embryo and a receptive uterus. In women, the uterus remains a hostile environment and must undergo functional changes to convert to a receptive state for embryo implantation. Determining uterine receptivity is vital in IVF treatment, as the timing of embryo transfer needs to be synchronized with uterine receptivity. However, to date, no reliable biochemical tests are available to determine uterine receptivity. We recently established that removal of α-dystroglycan N-terminus (α-DG-N) from the uterine surface plays an important role in the establishment of uterine receptivity. Importantly, the α-DG-N removed from the uterine tissue enters into the uterine fluid, and the levels correlate with the tissue status of receptivity. Detection of α-DG-N in uterine fluid may therefore provide a nonsurgical approach to assess uterine receptivity. In this study, we first validated three monoclonal antibodies raised against α-DG-N in our system, and then established a sandwich ELISA suitable for the detection of α-DG-N in human uterine fluid. This ELISA detected significantly higher concentrations of α-DG-N in uterine fluid of women in the receptive phase. We believe this newly established α-DG-N ELISA may provide an important tool in the development of noninvasive strategies to detect uterine receptivity in women.
Introduction
Embryo implantation requires a healthy embryo and a receptive uterus. In women, the uterus becomes receptive for embryo implantation only in the midsecretory phase of each menstrual cycle, known as the “window of implantation.” 1 Failure to establish receptivity is associated with unexplained infertility 2 and is a limiting factor in in vitro fertilization (IVF) treatment. The timing of embryo transfer needs to be synchronized with uterine receptivity in IVF; however, in current practice, no biochemical tests are available to determine uterine receptivity. A practical test is thus needed to improve IVF outcomes.
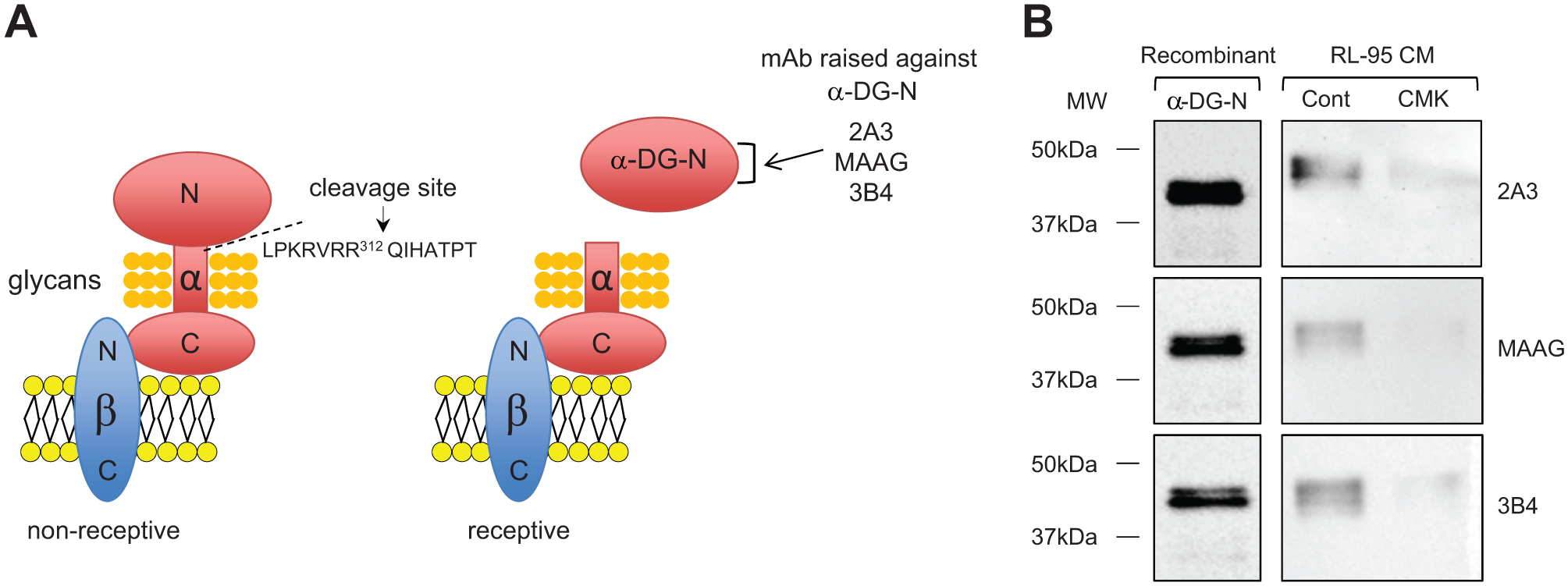
Our recent study has discovered that dystroglycan (DG), a large cell surface glycoprotein/adhesion molecule, plays an important role in the establishment of uterine receptivity and may provide a novel biomarker. 3 DG has been extensively studied in various tissues as an important adhesive molecule, as it acts as a linker between the extracellular matrix and the cytoskeleton. 4 In the human, DG is comprised of β- and α-DG subunits that are derived from a single gene product. 5 While β-DG is anchored within the plasma membrane, α-DG is noncovalently associated with the extracellular N-terminus of β-DG 5 ( Fig. 1A ). The α-DG has a well-defined dumbbell shape comprised of two large globular domains and a central mucin-like region 6 ( Fig. 1A ). The central region of α-DG (amino acids 316–485) is highly glycosylated and known to mediate cellular adhesion;7,8 however, this region is masked by its large N-terminus (α-DG-N). Proteolytic cleavage at amino acid 312 removes α-DG-N9,10 and unmasks the central glycosylated region for binding to extracellular molecules.
(
We have recently reported that α-DG-N removal plays a critical role in the establishment of uterine receptivity for embryo implantation in women. 3 The full-length α-DG on the uterine surface acts as a barrier for embryo attachment, and uterine receptivity requires proteolytic removal of α-DG-N through cleavage at RVRR312 by proprotein convertase 6 (PC6) 3 ( Fig. 1A ). Furthermore, the α-DG-N removed from the uterine tissue is detected in the uterine fluid, and the levels are significantly increased in the receptive phase. 3 We thus have proposed that α-DG-N in uterine fluid, which can be obtained noninvasively, might provide a potential biomarker for the detection of uterine tissue receptivity. 3 However, there are no high-throughput assays available for α-DG-N. In this study, we aimed to develop an enzyme-linked immunosorbent assay (ELISA) suitable for the detection of α-DG-N in the human uterine fluid.
Materials and Methods
Western Blot Analysis of Recombinant α-DG-N and Conditioned Media of RL-95 Cells
Recombinant α-DG-N (50 ng, Abnova, Taiwan) was analyzed by standard Western blot (10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis [SDS-PAGE] gels) using three monoclonal antibodies (mAbs) raised against α-DG-N: 2A3 (at 1 µg/mL, Sigma Aldrich, Castle Hill, NSW, Australia), MAAG, and 3B4 (at 1 µg/mL, Creative Diagnostic, Shirley, NY), and the signals were visualized by chemiluminescence (Thermo Scientific, Waltham, MA). Human endometrial epithelial RL-95 cells (ATCC, Manassas, VA) were cultured in Dulbecco’s Modified Eagle’s Medium—Nutrient Mixture F-12 (DMEM/F12, Life Technologies, Carlsbad, CA) supplemented with 10% (v/v) fetal calf serum (FCS; Life Technologies), and treated with or without 200 µM of PC inhibitor decanoyl-Arg-Val-Lys-Arg-chloromethylketone (to be referred to as CMK, Bachem, Torrance, CA) for 48 h. The conditioned media were collected, centrifuged at 1000 g for 5 min, and analyzed by Western blot as for the recombinant α-DG-N.
Biotinylation of α-DG-N Monoclonal Antibodies
The α-DG-N mAbs were buffer exchanged with Zeba desalting spin 7K MWCO columns (Thermo Scientific) that were washed with BupH PBS (0.1 M sodium phosphate, 0.15 M NaCl, pH 7.2, Thermo Scientific) and assessed by nanodrop (ND-1000, Biolab Group, Montrose, VIC, Australia) for concentration. Chromalink Biotin (Solulink, San Diego, CA) was dissolved in 100% dimethylformamide (DMF, Sigma Aldrich) to make a stock concentration of 10 mg/mL; it was further diluted in PBS to 5 mg/mL and used to conjugate the mAbs at a 1:30 ratio of biotin to antibody (example: 1 µg biotin to 30 µg antibody). After adding the biotin to the mAbs, the tubes were wrapped in foil to protect it from light and incubated at room temperature for 2 h on a rotating wheel. The biotin–mAb mixture was then passed through the Zeba columns by centrifugation at 1000 g for 2 min, and the conjugated mAbs were collected from the flow-through. The biotinylated antibodies were assessed for concentration by nanodrop, aliquoted, and stored at −20 °C for future use.
Establishment of an α-DG-N Sandwich ELISA
To establish an α-DG-N sandwich ELISA, we tested the three α-DG-N mAbs as either the coating or the biotinylated detection antibody (six possible combinations in total) to select the best combination. Half-area 96-well plates were coated overnight at 4 °C with 5 µg/mL of each α-DG-N mAb in 0.1 M sodium carbonate/bicarbonate buffer, pH 9.6 (Sigma Aldrich). The wells were then washed with PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 1.8 mM KH2PO4) containing 0.05% Tween 20 (PBS-T), and blocked with 1% bovine serum albumin (BSA; Bovogen, East Keilor, VIC, Australia) in PBS for 90 min at 37 °C. The wells were washed with PBS-T again and incubated with recombinant α-DG-N or analytes for 2 h at room temperature with gentle agitation. Following wash, the wells were incubated first with 5 µg/mL of biotinylated detection antibody for 1 h at room temperature, and then with polystreptavidin-HRP (Thermo Scientific) at 1:25,000 dilution for 50 min at room temperature with gentle agitation, and finally with ultra-TMB substrate (used undiluted, Thermo Scientific) for 7–10 min in the dark at room temperature without agitation. The reaction was stopped with 1 M sulfuric acid (Banksia Scientific Company, Hamilton, QLD, Australia) and the absorbance at 450 nm was measured (Envision Multilabel Reader, PerkinElmer, Waltham, MA). The best antibody combination was selected and further optimized by titration for optimal concentration of both the coating and the detection antibody and the polystreptavidin-HRP. The final α-DG-N ELISA consisted of 2A3 (0.5 µg/mL) for coating and biotinylated 3B4 (3 µg/mL) for detection. The sensitivity and intra- and interassay variabilities of this assay were determined on the standards and quality controls (QCs).
ELISA Detection of α-DG-N in Media of Ishikawa Cells Stably Expressing Different Forms of DG and in Human Uterine Lavage
Our previous studies have established human endometrial Ishikawa cells stably expressing two forms of DG: wild-type DG (WT-DG), which contains the endogenous PC6 cleavage site at RVRR312, and mutant DG (Mut-DG), in which the PC6 cleavage site was mutated to prevent the removal of α-DG-N. 3 These cells were first cultured to approximately 80% confluency in complete media containing MEM medium (Life Technologies) supplemented with 10% (v/v) FCS (Life Technologies) and 2% geneticin (Sigma Aldrich, St. Louis, MO), and then in serum-free media for 48 h. Serum-free conditioned media were collected, centrifuged at 1000 g for 5 min to remove cellular debris, and assayed for α-DG-N by the established α-DG-N ELISA.
Uterine lavages (n = 6) were obtained from women with proven fertility during the proliferative (Prolif, n = 3) and secretory (Sec, n = 3) phases of the menstrual cycle as previously described. 2 Ethical approval was obtained from appropriate institutional ethical committees, along with written informed consent from each individual. Uterine α-DG-N levels in these uterine lavages were also determined by the α-DG-N ELISA.
Statistics: Data are expressed as mean ± SEM. Statistical analysis was performed on raw data; two group comparisons used unpaired t test (PRISM version 6.00, GraphPad Software, San Diego, CA). p < 0.05 was taken as significant and p < 0.0005 was considered highly significant.
Results and Discussion
Confirmation of α-DG-N mAbs by Western Blot Analysis
We obtained three α-DG-N mAbs raised against α-DG-N ( Fig. 1A ). To confirm the specificity of these α-DG-N mAbs, Western blot analysis was performed on recombinant α-DG-N and conditioned media of human endometrial epithelial cell line RL-95 treated with or without a proprotein convertase inhibitor CMK. All three mAbs detected recombinant α-DG-N ( Fig. 1B ). All these mAbs also detected bands of expected sizes in the conditioned media of RL-95 cells, and the levels were reduced following CMK treatment ( Fig. 1B ). These data are consistent with α-DG-N removal from the plasma membrane to the media being inhibited by CMK in RL-95 cells. These data in turn confirmed the specificity of these mAbs for the detection of the cleaved α-DG-N in media.
Establishment of an α-DG-N ELISA
To develop a sandwich ELISA for the detection of α-DG-N in human uterine fluid, we tested these three α-DG-N mAbs in a total of six possible antibody combinations, with each as the catch or the detection antibody. We first tested these combinations on recombinant α-DG-N ( Fig. 2A ). All six combinations equally detected recombinant α-DG-N ( Fig. 2A ). However, this was not the case when a human uterine lavage sample was tested ( Fig. 2B ); out of the six combinations, the pair with mAb 2A3 as the coating and mAb 3B4 as the biotinylated detection antibody stood out as the best for the detection of α-DG-N in uterine fluid ( Fig. 2B ). This pair was further optimized by titration of all the components involved, and a representative standard curve for the final α-DG-N sandwich ELISA is shown in Figure 2C . To confirm the specificity of this α-DG-N ELISA, we assayed the media of RL-95 cells treated with or without CMK inhibitor as for Figure 1B . The ELISA detected four times more α-DG-N in the media of control than in CMK-treated cells ( Fig. 2D ), consistent with the Western blot results ( Fig. 1B ).
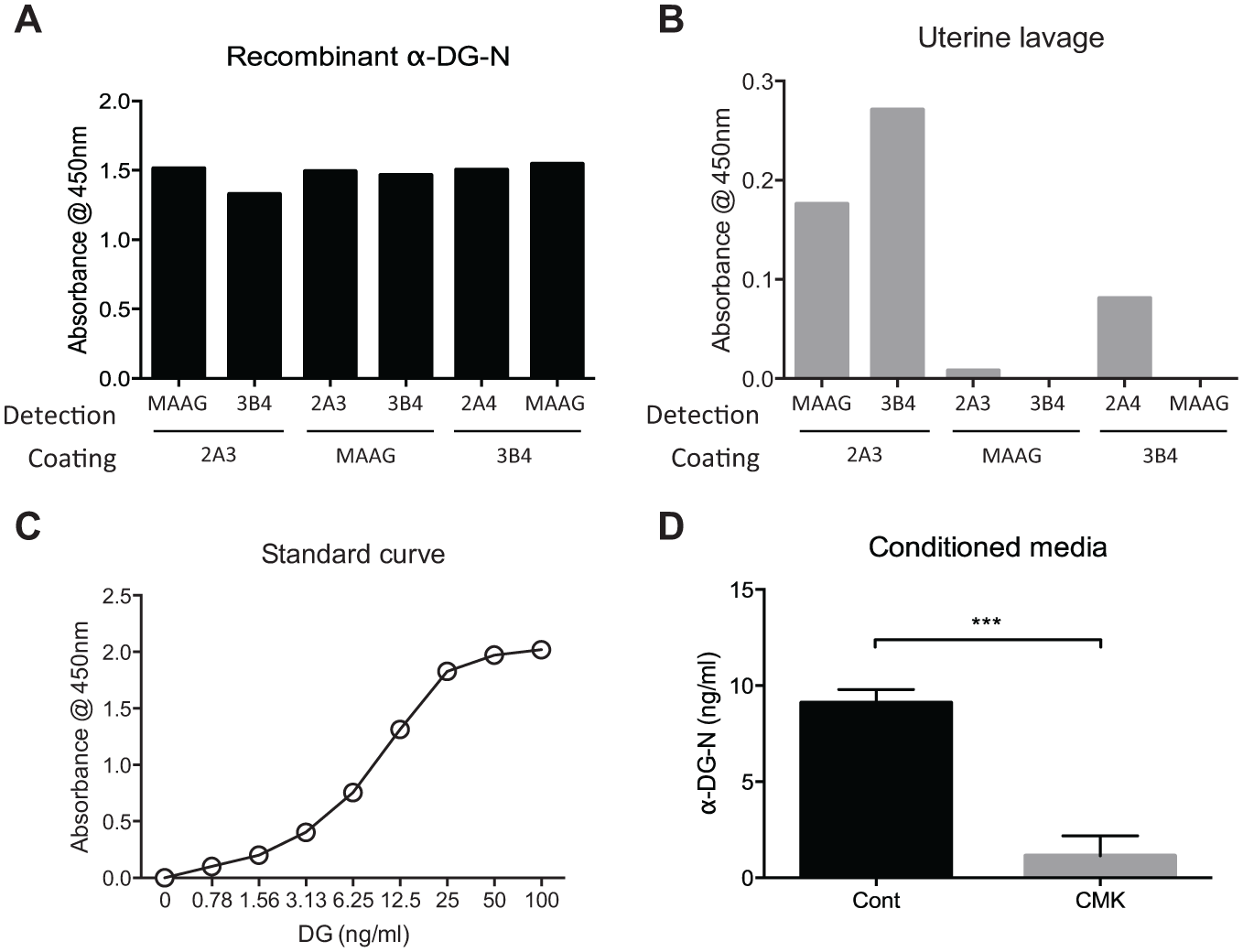
Antibody pair selection for the establishment of an α-DG-N ELISA. (
Linearity of the α-DG-N ELISA
Conditioned media from RL-95 cells transfected with DG and a uterine lavage from the secretory phase were diluted to different ranges and assayed to determine percentage recovery ( Table 1A ). The ELISA detected α-DG-N in these samples in a dilution-dependent manner, with good concentration recovery for each dilution ( Table 1A ).
Linearity Test of the α-DG-N ELISA.
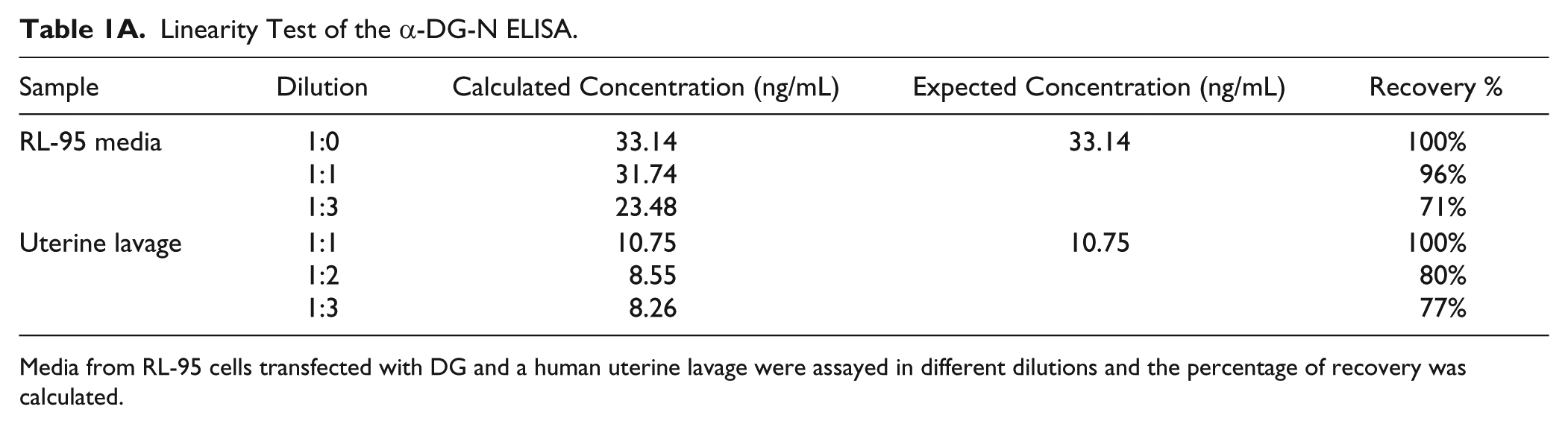
Media from RL-95 cells transfected with DG and a human uterine lavage were assayed in different dilutions and the percentage of recovery was calculated.
The Intra- and Interassay Variations of the α-DG-N ELISA
The accuracy and reproducibility of the α-DG-N ELISA were examined by determining its intra-assay (within a plate) and interassay (within different plates on different days) variations on eight replicates of standards and quality controls (conditioned media of Ishikawa cells stably expressing WT-DG) ( Table 1B ). The intra-assay variation was within 0.3%–6%, while the interassay variation was within 2%–20% ( Table 1B ).
Intra- and Interassay Variations of the α-DG-N ELISA on Standards and Quality Controls.
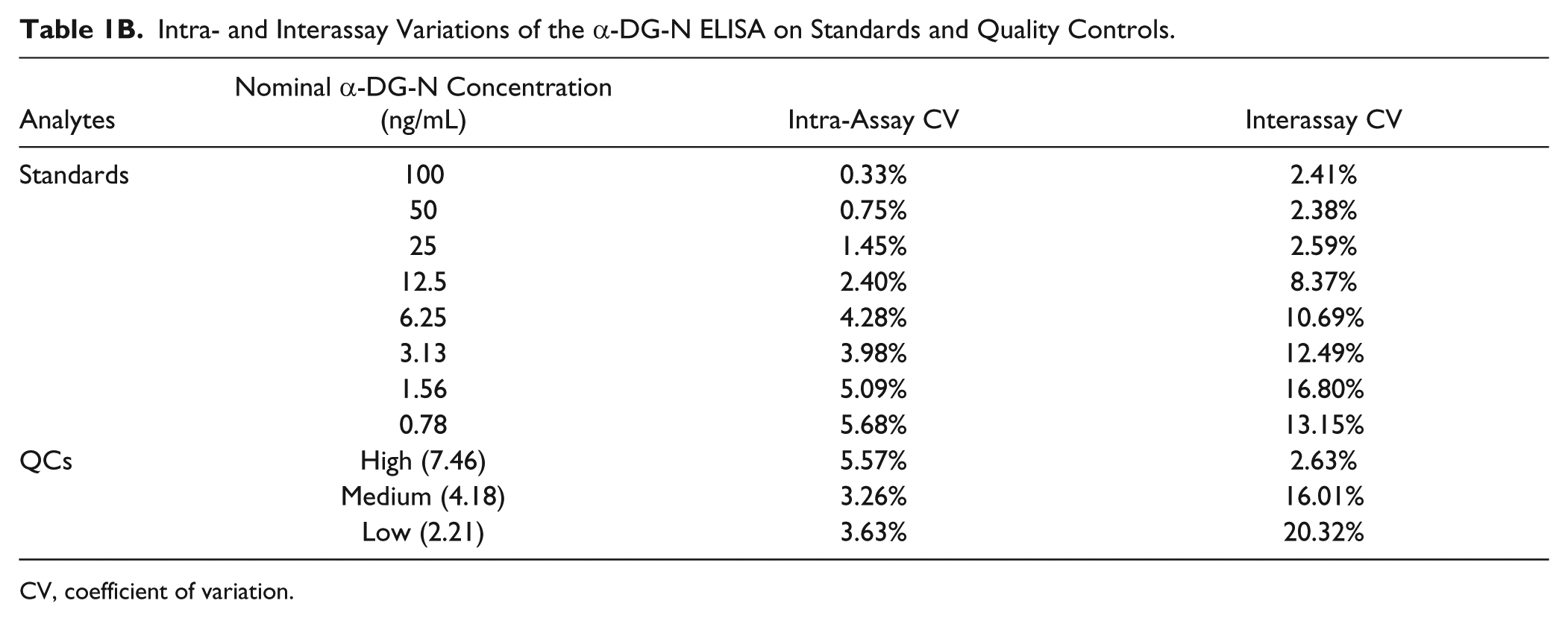
CV, coefficient of variation.
Validation of the α-DG-N ELISA on Samples Known to Contain Different Levels of Cleaved α-DG-N
The concentration of α-DG-N was determined in conditioned media of Ishikawa cells stably expressing different forms of DG: WT-DG and Mut-DG. 3 Our previous work has established that the α-DG-N levels in the media were significantly lower in cells expressing Mut-DG than WT-DG. This is because α-DG-N is cleaved off the cell membrane and released into the media in WT-DG cells, but in Mut-DG cells, α-DG-N is retained on the plasma membrane due to a mutation introduced to the RVRR312 cleavage site. 3 When media from these cells were subjected to the α-DG-N ELISA, as expected, a significant difference in α-DG-N concentration was detected between the two cell types ( Fig. 3A ).
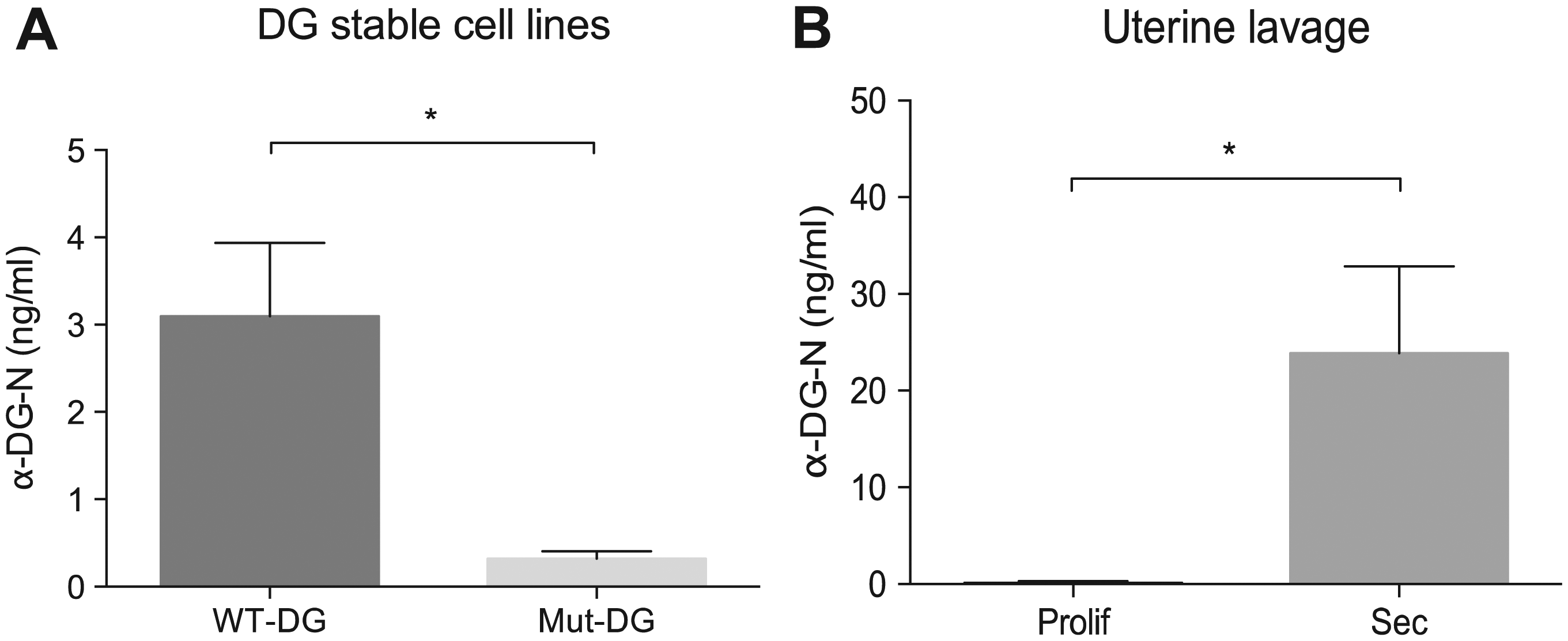
ELISA detection of α-DG-N in physiological samples. (
We next determined α-DG-N in human uterine lavage collected from women at different phases of the menstrual cycle. The levels were significantly higher in the secretory (receptive) than the proliferative (nonreceptive) phase ( Fig. 3B ). These results are again consistent with our previous findings using Western blot, 3 demonstrating the utility of this ELISA for the detection of α-DG-N in human uterine fluid.
The human uterus is a dynamic tissue that undergoes cyclic remodeling each month in preparation for embryo implantation. Defects in uterine preparation for receptivity will lead to implantation failure, which is a major concern in IVF treatment. To date, no reliable tests are available to determine uterine receptivity. Our previous study has established that uterine removal of α-DG-N, which enters into the uterine fluid, may provide a marker for receptivity. We believe this newly established α-DG-N ELISA may provide an important tool in the development of noninvasive strategies to detect uterine receptivity in women using uterine fluid instead of tissue biopsies. Future studies will need to establish whether this assay needs to be applied in a testing cycle immediately prior to the embryo transfer cycle, as previous studies indicate that disrupting the uterine fluid environment around the time of implantation may adversely affect implantation rate. 11
Footnotes
Acknowledgements
We thank Professor Lois Salamonsen and Dr. Tracey Edgell for providing the uterine lavage samples, our research nurse Judi Hocking for collecting the clinical materials, and the patients who kindly donated uterine lavage samples for this study.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Health and Medical Research Council of Australia (project grant no. 1044182 and fellowship no. 1041835 [to G.N.]), Monash IVF, and the Victorian Government Operational Infrastructure Support Program.